
Early Functional Impairment in Experimental Glaucoma Is Accompanied by Disruption of the GABAergic System and Inceptive Neuroinflammation

 and
and
Abstract
:1. Introduction
2. Results
2.1. Intraocular Injection of Ad5 Viral Vector Expressing Myocilin Y437H Results in Chronic IOP Elevation and RGC Loss
2.2. Slow Progressive RGC Degeneration after Ad5.MYOC Induced IOP Elevation
2.3. IOP Elevation Causes Rapid Retinal Expression of Inflammation Markers and Downregulation of GABAA Receptor Subunits and Glutamine Synthetase
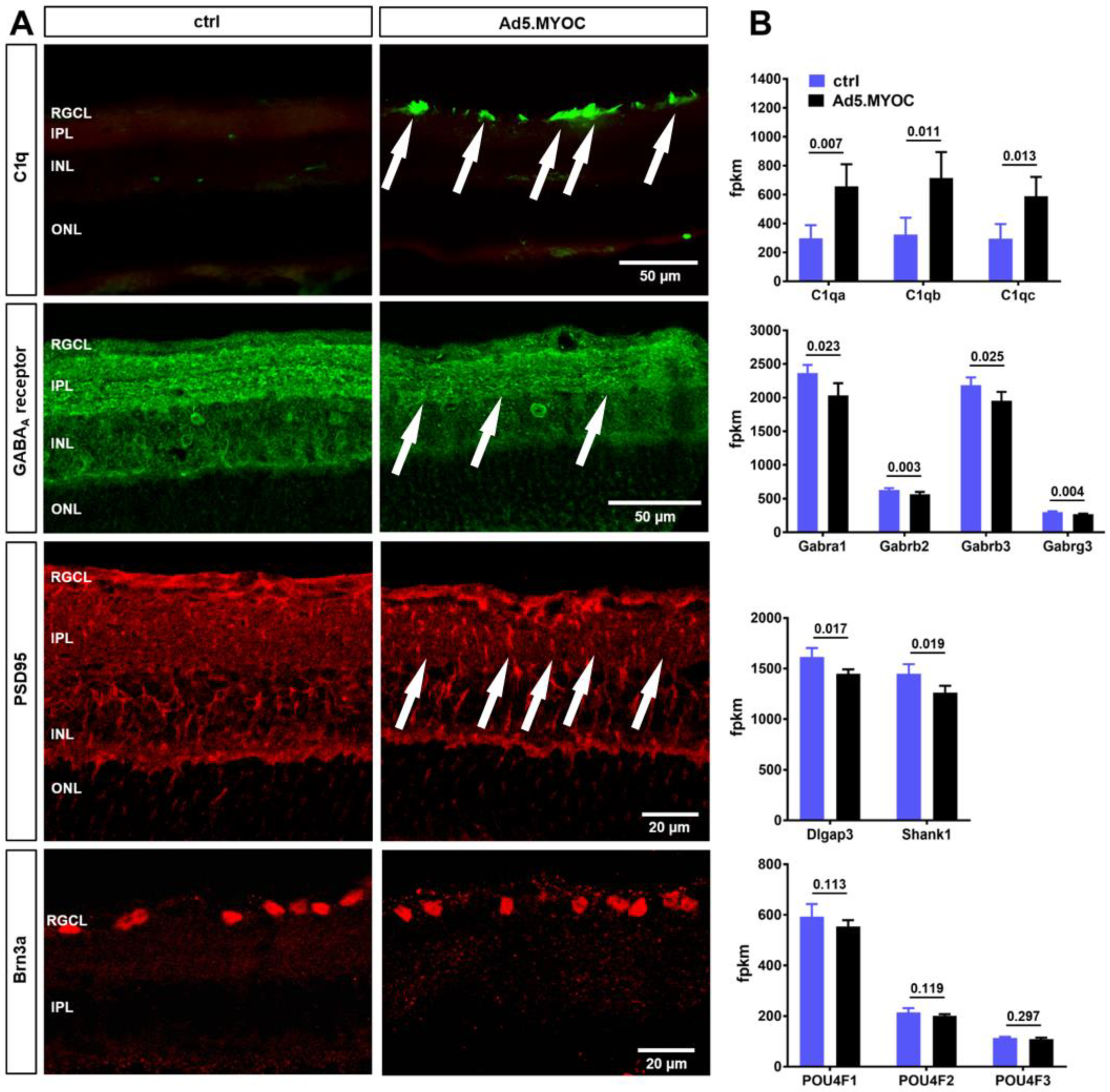
2.4. Hypertensive Eyes Display Retinal C1q Accumulation Accompanied by Reduced GABAA Receptor and PSD95 Immunolabeling
2.5. IOP Elevation Causes Decline in Pattern ERG Amplitudes
3. Discussion
3.1. Intraocular Injection of Ad5.MYOC Leads to Ocular Hypertension, Functional Decline, and Eventual RGC Loss
3.2. Functional Impairment in Hypertensive Eyes Is Accompanied by Synaptic Degeneration, Altered Glutamine Synthase, and Decreased GABAA-Receptor Expression
3.3. Inceptive Neuroinflammation Features C1q Deposition and Antigen Presentation
4. Materials and Methods
4.1. Animals
4.2. Myocilin Vector-Induced Glaucoma Model
4.3. Tonometry
4.4. Pattern ERG Recording
4.5. Analysis of RGC Survival
4.6. RNA-Sequencing
4.7. Immunohistology
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Weinreb, R.N.; Khaw, P.T. Primary open-angle glaucoma. Nat. Rev. Dis Primers 2016, 2, 16067. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.L.; Miglior, S. Risk factors for glaucoma onset and progression. Surv. Ophthalmol. 2008, 53 (Suppl. 1), S3–S10. [Google Scholar] [CrossRef]
- Leske, M.C.; Heijl, A.; Hussein, M.; Bengtsson, B.; Hyman, L.; Komaroff, E. Factors for glaucoma progression and the effect of treatment: The early manifest glaucoma trial. Arch. Ophthalmol. 2003, 121, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Leske, M.C.; Heijl, A.; Hyman, L.; Bengtsson, B.; Dong, L.; Yang, Z. Predictors of long-term progression in the early manifest glaucoma trial. Ophthalmology 2007, 114, 1965–1972. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, C.; Butt, Z. Blood flow velocity in the peripheral circulation of glaucoma patients. Ophthalmologica 1999, 213, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Flammer, J.; Orgül, S.; Costa, V.P.; Orzalesi, N.; Krieglstein, G.K.; Serra, L.M.; Renard, J.-P.; Stefánsson, E. The impact of ocular blood flow in glaucoma. Prog. Retin. Eye Res. 2002, 21, 359–393. [Google Scholar] [CrossRef]
- Inman, D.M.; Harun-Or-Rashid, M. Metabolic Vulnerability in the Neurodegenerative Disease Glaucoma. Front. Neurosci 2017, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Chrysostomou, V.; Rezania, F.; Trounce, I.; Crowston, J.G. Oxidative stress and mitochondrial dysfunction in glaucoma. Curr. Opin. Pharmacol. 2013, 13, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Harun-Or-Rashid, M.; Pappenhagen, N.; Zubricky, R.; Coughlin, L.; Jassim, A.H.; Inman, D.M. MCT2 overexpression rescues metabolic vulnerability and protects retinal ganglion cells in two models of glaucoma. Neurobiol. Dis. 2020, 141, 104944. [Google Scholar] [CrossRef]
- Soto, I.; Howell, G.R. The complex role of neuroinflammation in glaucoma. Cold Spring Harb. Perspect. Med. 2014, 4, a017269. [Google Scholar] [CrossRef]
- Bell, K.; Gramlich, O.W.; Von Thun Und Hohenstein-Blaul, N.; Beck, S.; Funke, S.; Wilding, C.; Pfeiffer, N.; Grus, F.H. Does autoimmunity play a part in the pathogenesis of glaucoma? Prog. Retin. Eye Res. 2013, 36, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Gharahkhani, P.; Jorgenson, E.; Hysi, P.; Khawaja, A.P.; Pendergrass, S.; Han, X.; Ong, J.S.; Hewitt, A.W.; Segrè, A.V.; Rouhana, J.M.; et al. Genome-wide meta-analysis identifies 127 open-angle glaucoma loci with consistent effect across ancestries. Nat. Commun. 2021, 12, 1258. [Google Scholar] [CrossRef] [PubMed]
- Fingert, J.H.; Stone, E.M.; Sheffield, V.; Alward, W.L. Myocilin glaucoma. Surv. Ophthalmol. 2002, 47, 547–561. [Google Scholar] [CrossRef]
- McDowell, C.M.; Luan, T.; Zhang, Z.; Putliwala, T.; Wordinger, R.; Millar, J.C.; John, S.W.; Pang, I.-H.; Clark, A.F. Mutant human myocilin induces strain specific differences in ocular hypertension and optic nerve damage in mice. Exp. Eye Res. 2012, 100, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Gramlich, O.W.; Godwin, C.R.; Heuss, N.D.; Gregerson, D.S.; Kuehn, M.H. T and B Lymphocyte Deficiency in Rag1−/− Mice Reduces Retinal Ganglion Cell Loss in Experimental Glaucoma. Investig. Ophthalmol. Vis. Sci. 2020, 61, 18. [Google Scholar] [CrossRef] [PubMed]
- Della Santina, L.; Inman, D.; Lupien, C.B.; Horner, P.J.; Wong, R.O.L. Differential progression of structural and functional alterations in distinct retinal ganglion cell types in a mouse model of glaucoma. J. Neurosci. 2013, 33, 17444–17457. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, J.; Seilheimer, R.L.; Tao, X.; Cowan, C.; Frankfort, B.J.; Wu, S.M. Elevated IOP alters the space-time profiles in the center and surround of both ON and OFF RGCs in mouse. Proc. Natl. Acad. Sci. USA 2017, 114, 8859–8864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puyang, Z.; Chen, H.; Liu, X. Subtype-dependent Morphological and Functional Degeneration of Retinal Ganglion Cells in Mouse Models of Experimental Glaucoma. J. Nat. Sci. 2015, 1, e103. [Google Scholar]
- Berry, R.H.; Qu, J.; John, S.W.M.; Howell, G.R.; Jakobs, T.C. Synapse Loss and Dendrite Remodeling in a Mouse Model of Glaucoma. PLoS ONE 2015, 10, e0144341. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.A.; Tribble, J.R.; Pepper, K.W.; Cross, S.D.; Morgan, B.P.; Morgan, J.E.; John, S.W.M.; Howell, G.R. Inhibition of the classical pathway of the complement cascade prevents early dendritic and synaptic degeneration in glaucoma. Mol. Neurodegener. 2016, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Zhang, R.; Zhang, S.; Wu, J.; Sun, X. Activation of 5-HT1A Receptors Promotes Retinal Ganglion Cell Function by Inhibiting the cAMP-PKA Pathway to Modulate Presynaptic GABA Release in Chronic Glaucoma. J. Neurosci. 2019, 39, 1484–1504. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zong, Y.; Zhang, R.; Zhang, X.; Zhang, S.; Wu, J.; Sun, X. Differential Modulation of GABAA and NMDA Receptors by an alpha7-nicotinic Acetylcholine Receptor Agonist in Chronic Glaucoma. Front. Mol. Neurosci. 2017, 10, 422. [Google Scholar] [CrossRef] [Green Version]
- Krizaj, D.; Ryskamp, D.A.; Tian, N.; Tezel, G.; Mitchell, C.H.; Slepak, V.Z.; Shestopalov, V.I. From mechanosensitivity to inflammatory responses: New players in the pathology of glaucoma. Curr. Eye Res. 2014, 39, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Johnson, E.C.; Cepurna, W.O.; Dyck, J.A.; Doser, T.; Morrison, J.C. Early gene expression changes in the retinal ganglion cell layer of a rat glaucoma model. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1460–1473. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.C.; Cepurna, W.O.; Tehrani, S.; Choe, T.E.; Jayaram, H.; Lozano, D.C.; Fortune, B.; Johnson, E.C. A Period of Controlled Elevation of IOP (CEI) Produces the Specific Gene Expression Responses and Focal Injury Pattern of Experimental Rat Glaucoma. Investig. Opthalmol. Vis. Sci. 2016, 57, 6700–6711. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.A.; Howell, G.R.; Barbay, J.M.; Braine, C.E.; Sousa, G.; John, S.W.M.; Morgan, J.E. Retinal ganglion cell dendritic atrophy in DBA/2J glaucoma. PLoS ONE 2013, 8, e72282. [Google Scholar] [CrossRef] [PubMed]
- Risner, M.L.; Pasini, S.; Cooper, M.; Lambert, W.; Calkins, D.J. Axogenic mechanism enhances retinal ganglion cell excitability during early progression in glaucoma. Proc. Natl. Acad. Sci. USA 2018, 115, E2393–E2402. [Google Scholar] [CrossRef] [Green Version]
- Bach, M.; Unsoeld, A.S.; Philippin, H.; Staubach, F.; Maier, P.; Walter, H.S.; Bomer, T.G.; Funk, J. Pattern ERG as an early glaucoma indicator in ocular hypertension: A long-term, prospective study. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4881–4887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilsey, L.J.; Fortune, B. Electroretinography in glaucoma diagnosis. Curr. Opin. Ophthalmol. 2016, 27, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Preiser, D.; Lagrèze, W.A.; Bach, M.; Poloschek, C.M. Photopic negative response versus pattern electroretinogram in early glaucoma. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1182–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porciatti, V.; Saleh, M.; Nagaraju, M. The pattern electroretinogram as a tool to monitor progressive retinal ganglion cell dysfunction in the DBA/2J mouse model of glaucoma. Investig. Ophthalmol. Vis. Sci. 2007, 48, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Zode, G.S.; Kuehn, M.H.; Nishimura, D.Y.; Searby, C.; Mohan, K.; Grozdanic, S.D.; Bugge, K.; Anderson, M.; Clark, A.F.; Stone, E.M.; et al. Reduction of ER stress via a chemical chaperone prevents disease phenotypes in a mouse model of primary open angle glaucoma. J. Clin. Investig. 2011, 121, 3542–3553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandalon, S.; Könnecke, B.; Levkovitch-Verbin, H.; Simons, M.; Hein, K.; Sättler, M.B.; Bähr, M.; Ofri, R. Functional and structural evaluation of lamotrigine treatment in rat models of acute and chronic ocular hypertension. Exp. Eye Res. 2013, 115, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Gallego, B.I.; Salazar, J.J.; De Hoz, R.; Rojas, B.; I Ramírez, A.; Navarro, M.Á.S.; Ortín-Martínez, A.; Soriano, F.J.V.; Avilés-Trigueros, M.; Pérez, M.P.V.; et al. IOP induces upregulation of GFAP and MHC-II and microglia reactivity in mice retina contralateral to experimental glaucoma. J. Neuroinflamm. 2012, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piri, N.; Kwong, J.M.; Caprioli, J. Crystallins in retinal ganglion cell survival and regeneration. Mol. Neurobiol. 2013, 48, 819–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kompass, K.S.; A Agapova, O.; Li, W.; Kaufman, P.L.; A Rasmussen, C.; Hernandez, M.R. Bioinformatic and statistical analysis of the optic nerve head in a primate model of ocular hypertension. BMC Neurosci. 2008, 9, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Q.J.; Cook, A.C.; Dumitrescu, A.; Kuehn, M.H. Lack of immunoglobulins does not prevent C1q binding to RGC and does not alter the progression of experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6370–6377. [Google Scholar] [CrossRef] [Green Version]
- Kuehn, M.H.; Kim, C.Y.; Ostojic, J.; Bellin, M.; Alward, W.L.; Stone, E.M.; Sakaguchi, D.; Grozdanic, S.D.; Kwon, Y. Retinal synthesis and deposition of complement components induced by ocular hypertension. Exp. Eye Res. 2006, 83, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Brown, K.M.; Stephan, D.A.; Morrison, J.C.; Johnson, E.C.; Tomarev, S.I. Microarray Analysis of Changes in mRNA Levels in the Rat Retina after Experimental Elevation of Intraocular Pressure. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1247–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tezel, G.; Yang, X.; Luo, C.; Kain, A.D.; Powell, D.W.; Kuehn, M.H.; Kaplan, H.J. Oxidative stress and the regulation of complement activation in human glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5071–5082. [Google Scholar] [CrossRef]
- Stasi, K.; Nagel, D.; Yang, X.; Wang, R.-F.; Ren, L.; Podos, S.M.; Mittag, T.; Danias, J. Complement component 1Q (C1Q) upregulation in retina of murine, primate, and human glaucomatous eyes. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1024–1029. [Google Scholar] [CrossRef] [Green Version]
- Wassle, H.; Koulen, P.; Brandstätter, J.H.; Fletcher, E.; Becker, C.-M. Glycine and GABA receptors in the mammalian retina. Vis. Res. 1998, 38, 1411–1430. [Google Scholar] [CrossRef] [Green Version]
- Euler, T.; Wassle, H. Different contributions of GABAA and GABAC receptors to rod and cone bipolar cells in a rat retinal slice preparation. J. Neurophysiol. 1998, 79, 1384–1395. [Google Scholar] [CrossRef]
- Eggers, E.D.; Lukasiewicz, P.D. Multiple pathways of inhibition shape bipolar cell responses in the retina. Vis. Neurosci. 2011, 28, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.T.; Alyahya, K.; Gionfriddo, J.R.; Dubielzig, R.R.; Madl, J.E. Loss of glutamine synthetase immunoreactivity from the retina in canine primary glaucoma. Vet. Ophthalmol. 2008, 11, 150–157. [Google Scholar] [CrossRef]
- Moreno, M.C.; Sande, P.; Marcos, H.A.; De Zavalía, N.; Sarmiento, M.I.K.; Rosenstein, R.E. Effect of glaucoma on the retinal glutamate/glutamine cycle activity. FASEB J. 2005, 19, 1161–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, M. Abnormalities in glutamate metabolism and excitotoxicity in the retinal diseases. Scientifica 2013, 2013, 528940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaked, I.; Ben-Dror, I.; Vardimon, L. Glutamine synthetase enhances the clearance of extracellular glutamate by the neural retina. J. Neurochem. 2002, 83, 574–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, T.; Harada, C.; Nakamura, K.; Quah, H.-M.A.; Okumura, A.; Namekata, K.; Saeki, T.; Aihara, M.; Yoshida, H.; Mitani, A.; et al. The potential role of glutamate transporters in the pathogenesis of normal tension glaucoma. J. Clin. Investig. 2007, 117, 1763–1770. [Google Scholar] [CrossRef] [Green Version]
- Bak, L.K.; Schousboe, A.; Waagepetersen, H.S. The glutamate/GABA-glutamine cycle: Aspects of transport, neurotransmitter homeostasis and ammonia transfer. J. Neurochem. 2006, 98, 641–653. [Google Scholar] [CrossRef]
- Barnett, N.L.; Pow, D.V.; Robinson, S.R. Inhibition of Muller cell glutamine synthetase rapidly impairs the retinal response to light. Glia 2000, 30, 64–73. [Google Scholar] [CrossRef]
- Dhingra, A.; Vardi, N. “mGlu Receptors in the Retina”—WIREs Membrane Transport and Signaling. Wiley Interdiscip Rev. Membr. Transp. Signal 2012, 1, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Cuenca, N.; Pinilla, I.; Fernández-Sánchez, L.; Salinas-Navarro, M.; Alarcón-Martínez, L.; Avilés-Trigueros, M.; De La Villa, P.; De Imperial, J.M.; Villegas-Pérez, M.P.; Vidal-Sanz, M. Changes in the inner and outer retinal layers after acute increase of the intraocular pressure in adult albino Swiss mice. Exp. Eye Res. 2010, 91, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.J.; Frankfort, B.J.; Gross, R.L.; Wu, S.M. Elevated intraocular pressure decreases response sensitivity of inner retinal neurons in experimental glaucoma mice. Proc. Natl. Acad. Sci. USA 2015, 112, 2593–2598. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Luo, X.; Liu, S.; Shen, Y.; Nawy, S.; Shen, Y. Rod bipolar cells dysfunction occurs before ganglion cells loss in excitotoxin-damaged mouse retina. Cell Death Dis. 2019, 10, 905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naisbitt, S.; Kim, E.; Tu, J.C.; Xiao, B.; Sala, C.; Valtschanoff, J.; Weinberg, R.J.; Worley, P.F.; Sheng, M. Shank, a novel family of postsynaptic density proteins that binds to the NMDA receptor/PSD-95/GKAP complex and cortactin. Neuron 1999, 23, 569–582. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, M.; Hata, Y.; Hirao, K.; Toyoda, A.; Irie, M.; Takai, Y. SAPAPs. A family of PSD-95/SAP90-associated proteins localized at postsynaptic density. J. Biol. Chem. 1997, 272, 11943–11951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowston, J.G.; Kong, Y.X.G.; Trounce, I.; Dang, T.M.; Fahy, E.T.; Bui, B.; Morrison, J.C.; Chrysostomou, V. An acute intraocular pressure challenge to assess retinal ganglion cell injury and recovery in the mouse. Exp. Eye Res. 2015, 141, 3–8. [Google Scholar] [CrossRef]
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.S.; Nouri, N.; Micheva, K.D.; Mehalow, A.K.; Huberman, A.D.; Stafford, B.; et al. The classical complement cascade mediates CNS synapse elimination. Cell 2007, 131, 1164–1178. [Google Scholar] [CrossRef] [Green Version]
- Riese, R.J.; Wolf, P.R.; Brömme, D.; Natkin, L.R.; Villadangos, J.; Ploegh, H.L.; A Chapman, H. Essential role for cathepsin S in MHC class II-associated invariant chain processing and peptide loading. Immunity 1996, 4, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.P.; Villadangos, J.; Dranoff, G.; Small, C.; Gu, L.; Haley, K.J.; Riese, R.; Ploegh, H.L.; A Chapman, H. Cathepsin S required for normal MHC class II peptide loading and germinal center development. Immunity 1999, 10, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Bailey, J.N.; Yaspan, B.L.; Pasquale, L.R.; Hauser, M.A.; Kang, J.H.; Loomis, S.J.; Brilliant, M.; Budenz, D.L.; Christen, W.G.; Fingert, J.; et al. Hypothesis-independent pathway analysis implicates GABA and acetyl-CoA metabolism in primary open-angle glaucoma and normal-pressure glaucoma. Hum. Genet. 2014, 133, 1319–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murshid, A.; Gong, J.; Calderwood, S.K. The role of heat shock proteins in antigen cross presentation. Front. Immunol. 2012, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Binder, R.J. Functions of heat shock proteins in pathways of the innate and adaptive immune system. J. Immunol. 2014, 193, 5765–5771. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, M.; Flajnik, M.F. Origin and evolution of the specialized forms of proteasomes involved in antigen presentation. Immunogenetics 2019, 71, 251–261. [Google Scholar] [CrossRef]
- Williams, J.J.; Munro, K.M.; Palmer, T.M. Role of Ubiquitylation in Controlling Suppressor of Cytokine Signalling 3 (SOCS3) Function and Expression. Cells 2014, 3, 546–562. [Google Scholar] [CrossRef] [Green Version]
- Campello, L.; Esteve-Rudd, J.; Cuenca, N.; Martín-Nieto, J. The ubiquitin-proteasome system in retinal health and disease. Mol. Neurobiol. 2013, 47, 790–810. [Google Scholar] [CrossRef] [PubMed]
- Quaranta, L.; Bruttini, C.; Micheletti, E.; Konstas, A.G.; Michelessi, M.; Oddone, F.; Katsanos, A.; Sbardella, D.; De Angelis, G.; Riva, I. Glaucoma and neuroinflammation: An overview. Surv. Ophthalmol. 2021, 66, 693–713. [Google Scholar] [CrossRef] [PubMed]
- Imani, F.; Soloski, M.J. Heat shock proteins can regulate expression of the Tla region-encoded class Ib molecule Qa-1. Proc. Natl. Acad. Sci. USA 1991, 88, 10475–10479. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.; Kalb, S.; Liang, B.; Aldrich, C.J.; Lemonnier, F.A.; Jiang, H.; Cotter, R.; Soloski, M.J. A peptide from heat shock protein 60 is the dominant peptide bound to Qa-1 in the absence of the MHC class Ia leader sequence peptide Qdm. J. Immunol. 2003, 170, 5027–5033. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Harper, M.; Kecova, H.; Aldamus, G.; Kardon, R.; Grozdanic, S.D.; Kuehn, M.H. Neuroinflammation in advanced canine glaucoma. Mol. Vis. 2010, 16, 2092–2108. [Google Scholar]
- Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Grus, F.H. Antibodies to alpha B-Crystallin, Vimentin, and Heat Shock Protein 70 in Aqueous Humor of Patients with Normal Tension Glaucoma and IgG Antibody Patterns Against Retinal Antigen in Aqueous Humor. Curr. Eye Res. 2007, 32, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Joachim, S.C.; Wax, M.B.; Seidel, P.; Pfeiffer, N.; Grus, F.H. Enhanced characterization of serum autoantibody reactivity following HSP 60 immunization in a rat model of experimental autoimmune glaucoma. Curr. Eye Res. 2010, 35, 900–908. [Google Scholar] [CrossRef]
- Wax, M.B.; Tezel, G.; Yang, J.; Peng, G.; Patil, R.V.; Agarwal, N.; Sappington, R.; Calkins, D.J. Induced autoimmunity to heat shock proteins elicits glaucomatous loss of retinal ganglion cell neurons via activated T-cell-derived fas-ligand. J. Neurosci. 2008, 28, 12085–12096. [Google Scholar] [CrossRef] [Green Version]
- Grotegut, P.; Hoerdemann, P.J.; Reinehr, S.; Gupta, N.; Dick, H.B.; Joachim, S.C. Heat Shock Protein 27 Injection Leads to Caspase Activation in the Visual Pathway and Retinal T-Cell Response. Int. J. Mol. Sci. 2021, 22, 513. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cho, K.-S.; Vu, T.H.K.; Shen, C.-H.; Kaur, M.; Chen, G.; Mathew, R.; McHam, M.L.; Fazelat, A.; Lashkari, K.; et al. Commensal microflora-induced T cell responses mediate progressive neurodegeneration in glaucoma. Nat. Commun. 2018, 9, 3209. [Google Scholar] [CrossRef]
- Tang, J.; Tang, Y.; Yi, I.; Chetn, D.F. The role of commensal microflora-induced T cell responses in glaucoma neurodegeneration. Prog. Brain Res. 2020, 256, 79–97. [Google Scholar] [PubMed]
- Kim, C.Y.; Kuehn, M.H.; Anderson, M.; Kwon, Y. Intraocular pressure measurement in mice: A comparison between Goldmann and rebound tonometry. Eye 2007, 21, 1202–1209. [Google Scholar] [CrossRef]
- Nadal-Nicolas, F.M.; Jiménez-Loópez, M.; Sobrado-Calvo, P.; Nieto-Loópez, L.; Cánovas-Martínez, I.; Salinas-Navarro, M.; Vidal-Sanz, M.; Agudo, M. Brn3a as a marker of retinal ganglion cells: Qualitative and quantitative time course studies in naive and optic nerve-injured retinas. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3860–3868. [Google Scholar] [CrossRef] [Green Version]
- Hedberg-Buenz, A.; Christopher, M.A.; Lewis, C.J.; Fernandes, K.A.; Dutca, L.M.; Wang, K.; Scheetz, T.; Abramoff, M.; Libby, R.T.; Garvin, M.K.; et al. Quantitative measurement of retinal ganglion cell populations via histology-based random forest classification. Exp. Eye Res. 2015, 146, 370–385. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, L.J.; Kuhn, M.; Stark, M.; Chaffron, S.; Creevey, C.; Muller, J.; Doerks, T.; Julien, P.; Roth, A.; Simonovic, M.; et al. STRING 8—A global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res. 2009, 37, D412–D416. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Downregulated mRNA Expression | ||||
| Gene | WT (FPKM ± SD) | Glaucoma (FPKM ± SD) | t Test | |
| GABAA receptor and GABA neurotransmitter recycling | Adcy2 | 1871 ± 81 | 1671 ± 82 | 0.0133 |
| Cacna1d | 979 ± 32 | 874 ± 57 | 0.0182 | |
| Gabbr2 | 1790 ± 53 | 1543 ± 87 | 0.0029 | |
| Gabra1 | 2367 ± 121 | 2034 ± 182 | 0.0226 | |
| Gabrb2 | 628 ± 29 | 565 ± 36 | 0.0328 | |
| Gabrb3 | 2188 ± 113 | 1954 ± 110 | 0.0249 | |
| Gabrg3 | 300 ± 11 | 268 ± 9 | 0.0037 | |
| Gad1 | 6470 ± 379 | 5441 ± 397 | 0.0095 | |
| Gad2 | 1269 ± 68 | 1113 ± 76 | 0.0226 | |
| Glul | 47,282 ± 621 | 33,233 ± 4840 | 0.0012 | |
| Gnao1 | 8319 ± 338 | 7421 ± 507 | 0.0257 | |
| Gng2 | 1226 ± 42 | 1066 ± 82 | 0.0132 | |
| Slc6a1 | 10,796 ± 661 | 9056 ± 913 | 0.0214 | |
| Slc38a1 | 5525 ± 205 | 4845 ± 405 | 0.0244 | |
| Cholinergic and glutamatergic synapses | Adcy2 | 1871 ± 81 | 1671 ± 82 | 0.0133 |
| Cacna1d | 979 ± 32 | 874 ± 57 | 0.0182 | |
| Camk4 | 566 ± 33 | 502 ± 23 | 0.0183 | |
| Chrm2 | 195 ± 3 | 172 ± 6 | 0.0004 | |
| Chrna4 | 624 ± 34 | 556 ± 34 | 0.0290 | |
| Dlgap3 | 1616 ± 87 | 1449 ± 43 | 0.0166 | |
| Glul | 47,282 ± 621 | 33,233 ± 4840 | 0.0012 | |
| Gnao1 | 8319 ± 338 | 7421 ± 507 | 0.0257 | |
| Gng2 | 1226 ± 42 | 1066 ± 82 | 0.0132 | |
| Gria2 | 1711 ± 56 | 1469 ± 70 | 0.0016 | |
| Grm2 | 339 ± 12 | 299 ± 24 | 0.0284 | |
| Grm6 | 2334 ± 163 | 2005 ± 100 | 0.0136 | |
| Kcnj2 | 217 ± 3 | 195 ± 7 | 0.0015 | |
| Prkca | 6764 ± 127 | 5909 ± 350 | 0.0037 | |
| Shank1 | 1449 ± 95 | 1263 ± 67 | 0.0186 | |
| Slc5a7 | 311 ± 17 | 268 ± 10 | 0.0054 | |
| Slc38a1 | 5525 ± 205 | 4845 ± 406 | 0.0244 | |
| Axon guidance | Dpysl2 | 1726 ± 56 | 1518 ± 76 | 0.0046 |
| Dpysl5 | 1231 ± 24 | 1094 ± 56 | 0.0042 | |
| Efnb3 | 522 ± 39 | 451 ± 32 | 0.0327 | |
| Epha8 | 1215 ± 71 | 997 ± 64 | 0.0038 | |
| L1cam | 1877 ± 119 | 1655 ± 83 | 0.0223 | |
| Plxna1 | 1114 ± 63 | 998 ± 47 | 0.0254 | |
| Rgs3 | 726 ± 38 | 646 ± 38 | 0.0251 | |
| Robo2 | 1986 ± 112 | 1742 ± 139 | 0.0334 | |
| Sema4g | 1067 ± 46 | 905 ± 80 | 0.0123 | |
| Sema6b | 791 ± 37 | 708 ± 28 | 0.0115 | |
| Slit1 | 523 ± 24 | 468 ± 18 | 0.0109 | |
| Slit2 | 2512 ± 179 | 2146 ± 138 | 0.0176 | |
| Srgap3 | 4500 ± 193 | 3953 ± 276 | 0.0176 | |
| Unc5b | 338 ± 12 | 292 ± 28 | 0.0228 | |
| Upregulated mRNA Expression | ||||
| Gene | WT (FPKM ± SD) | Glaucoma (FPKM ± SD) | t Test | |
| Antigen processing and presentation | Ctss | 561 ± 207 | 1319 ± 354 | 0.0101 |
| H2-T23 | 248 ± 48 | 452 ± 141 | 0.0336 | |
| Ifi30 | 151 ± 12 | 201 ± 29 | 0.0184 | |
| Psme2 | 500 ± 19 | 600 ± 49 | 0.0090 | |
| Tapbp | 1709 ± 136 | 2287 ± 268 | 0.0085 | |
| Complement cascade | A2m | 476 ± 157 | 1269 ± 297 | 0.0033 |
| C1qa | 297 ± 92 | 656 ± 154 | 0.0072 | |
| C1qb | 324 ± 116 | 715 ± 179 | 0.0107 | |
| C1qc | 295 ± 102 | 589 ± 133 | 0.0125 | |
| C3ar1 | 52 ± 4 | 65 ± 6 | 0.0109 | |
| C4b | 835 ± 599 | 2781 ± 1120 | 0.0221 | |
| Cfh | 309 ± 44 | 412 ± 54 | 0.0248 | |
| Cfi | 68 ± 2 | 83 ± 9 | 0.0199 | |
| Pros1 | 359 ± 41 | 513 ± 111 | 0.0405 | |
| Serping1 | 411 ± 186 | 1081 ± 489 | 0.0430 | |
| Cytokine–cytokine receptor signaling | Ccr5 | 44 ± 3 | 53 ± 4 | 0.0117 |
| Csf1r | 444 ± 112 | 865 ± 175 | 0.0066 | |
| Cx3cr1 | 199 ± 34 | 306 ± 54 | 0.0153 | |
| Cxcl10 | 41 ± 2 | 53 ± 9 | 0.0416 | |
| Dock2 | 61 ± 4 | 76 ± 8 | 0.0194 | |
| Gnb3 | 8209 ± 663 | 11,825 ± 1307 | 0.0026 | |
| Hck | 41 ± 2 | 50 ± 5 | 0.0225 | |
| Il10ra | 62 ± 3 | 75 ± 6 | 0.0066 | |
| Jak3 | 138 ± 13 | 206 ± 40 | 0.0175 | |
| Ltbr | 198 ± 18 | 252 ± 38 | 0.0430 | |
| Ncf1 | 65 ± 5 | 81 ± 9 | 0.0233 | |
| Osmr | 269 ± 55 | 507 ± 129 | 0.0146 | |
| Stat1 | 1613 ± 293 | 2730 ± 770 | 0.0351 | |
| Stat2 | 1117 ± 88 | 1343 ± 158 | 0.0467 | |
| Stat3 | 1815 ± 280 | 3209 ± 846 | 0.0204 | |
| Tnfrsf1a | 308 ± 60 | 517 ± 139 | 0.0334 | |
| Vav1 | 50 ± 3 | 61 ± 6 | 0.0173 | |
| Clusters of differentiation | Cd52 | 57 ± 5 | 74 ± 9 | 0.0177 |
| Cd53 | 63 ± 3 | 76 ± 9 | 0.0296 | |
| Cd63 | 975 ± 69 | 1351 ± 223 | 0.0183 | |
| Cd68 | 83 ± 9 | 108 ± 16 | 0.0323 | |
| Cd72 | 46 ± 4 | 55 ± 4 | 0.0171 | |
| Ribosomal subunits | Rpl10 | 2480 ± 74 | 3010 ± 250 | 0.0067 |
| Rpl13a | 7349 ± 247 | 8843 ± 789 | 0.0112 | |
| Rpl17 | 3028 ± 134 | 3756 ± 362 | 0.0092 | |
| Rpl19 | 2983 ± 133 | 3615 ± 268 | 0.0055 | |
| Rpl32 | 2190 ± 69 | 2631 ± 190 | 0.0048 | |
| Rpl41 | 2805 ± 98 | 3427 ± 337 | 0.0121 | |
| Rpl9 | 7008 ± 340 | 8670 ± 961 | 0.0172 | |
| Rplp0 | 4190 ± 138 | 5225 ± 478 | 0.0059 | |
| Rplp1 | 5660 ± 98 | 6845 ± 558 | 0.0058 | |
| Rps10 | 2794 ± 87 | 3379 ± 284 | 0.0077 | |
| Rps12 | 2473 ± 105 | 2973 ± 223 | 0.0066 | |
| Rps14 | 4689 ± 218 | 6181 ± 786 | 0.0106 | |
| Rps15 | 5240 ± 146 | 6408 ± 623 | 0.0107 | |
| Rps18 | 3420 ± 135 | 4242 ± 379 | 0.0065 | |
| Rps19 | 2340 ± 113 | 2826 ± 228 | 0.0088 | |
| Rps20 | 2210 ± 58 | 2656 ± 208 | 0.0061 | |
| Rps24 | 3900 ± 105 | 4802 ± 366 | 0.0032 | |
| Rps25 | 3129 ± 138 | 3762 ± 316 | 0.0105 | |
| Rps27a | 3820 ± 118 | 4650 ± 348 | 0.0040 | |
| Rps3 | 6347 ± 170 | 7880 ± 842 | 0.0118 | |
| Rps9 | 3916 ± 142 | 4702 ± 311 | 0.0037 | |
| Rras | 225 ± 16 | 299 ± 53 | 0.0373 | |
| Rtp4 | 63 ± 4 | 80 ± 11 | 0.0207 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gramlich, O.W.; Godwin, C.R.; Wadkins, D.; Elwood, B.W.; Kuehn, M.H. Early Functional Impairment in Experimental Glaucoma Is Accompanied by Disruption of the GABAergic System and Inceptive Neuroinflammation. Int. J. Mol. Sci. 2021, 22, 7581. https://doi.org/10.3390/ijms22147581
Gramlich OW, Godwin CR, Wadkins D, Elwood BW, Kuehn MH. Early Functional Impairment in Experimental Glaucoma Is Accompanied by Disruption of the GABAergic System and Inceptive Neuroinflammation. International Journal of Molecular Sciences. 2021; 22(14):7581. https://doi.org/10.3390/ijms22147581
Chicago/Turabian StyleGramlich, Oliver W., Cheyanne R. Godwin, David Wadkins, Benjamin W. Elwood, and Markus H. Kuehn. 2021. "Early Functional Impairment in Experimental Glaucoma Is Accompanied by Disruption of the GABAergic System and Inceptive Neuroinflammation" International Journal of Molecular Sciences 22, no. 14: 7581. https://doi.org/10.3390/ijms22147581