1. Introduction
Light is a key environmental factor affecting the physiology and evolution of most organisms. An extra-cellular light signal is transformed into an intra-cellular response by a photoreceptor built of the protein fragment and a photopigment. Photoreceptor proteins bind riboflavins, which are chromophores of plants. The major flavin—riboflavin [
1]—is synthesized by plants, fungi, and various bacteria, and is a flavin precursor (FMN—flavin mononucleotide, FAD—flavin adenine dinucleotide, and RF—riboflavin). Flavins are important compounds that are present in many biological systems. They take part in cell energetic processes, where they play the role of electron and proton transporters (as coenzymes), as well as chromophores in photoreceptors sensitive to blue light [
2,
3].
Several groups of flavin-related photoreceptors have been discovered [
2,
3,
4], such as the following: cryptochromes, with FAD and a pterin as chromophores, are responsible for the entrainment of circadian rhythms generated by light in various organisms, from bacteria and plants to humans, and they are also involved in the regulation of plant growth (e.g., inhibition of stem elongation); phototropins with two FMN-binding light-oxygen-voltage (LOV) domains, are responsible for phototropism, chloroplast relocation, and stomatal opening; Zeitlupe (ZTL), Flavin-binding Kelch F-box1 (FKF1), and LOV Kelch Protein 2 (LKP2) proteins, are a family of photoreceptors involved in modulating the circadian clock and regulating flowering; and BLUF photoreceptors with a FAD-binding domain are present in photosynthetic bacteria, cyanobacteria, and Euglena, and control photosynthesis gene expression.
In recent years, studies regarding the mechanisms of photocycles, in which molecules of FMN or FAD are transformed under light, have developed [
3,
5,
6,
7,
8,
9,
10]. However, the mechanism of signal transduction in photoreceptor phenomena, in which flavins take part, is not yet completely known.
Investigations conducted in the last decade have demonstrated that flavin-based- photoreceptors act as dimers or higher aggregates in living organisms [
11,
12,
13,
14]. Using spectroscopic NMR techniques and analytical ultracentrifugation, Dorn et al. [
15] demonstrated that flavin stabilizes LOV domain conformation and that dimerization is due to the intermolecular LOV–LOV interactions. Muralidhara and Wittung-Stafshede [
16] have demonstrated the existence of FMN dimers, which form a stacking structure in flavodoxin from
Desulfovibrio desulfuricans expressed in
Escherichia coli, in which FMN is a co-factor. They [
17] also reported the formation of such FMN dimers when FMN was binding to apoflavodoxin. Six riboflavin dimers were also detected in dodecin (a flavoprotein from archaeal
Halobacterium salinarium) via crystallographic analyses [
18]. Staudt et al. [
19] also demonstrated stacked dimers of riboflavin in archaeal dodecin. The distance between the two isoalloxazine moieties in these dimers was R = 3.5 Å; hence, it was the same as in the in vitro studies (R = 3.5 ± 0.3 Å for FMN in solution [
20], and R = 3.2 ± 0.3 Å in rigid PVA) [
21].
Therefore, it seems essential to identify the spectroscopic properties and structures of dimers, as well as their involvement in the process of nonradiative excitation energy transfer. Our previous investigations [
22,
23] have shown that FMN dimers (D) constitute perfect traps of the excitation energy in water solutions while, in rigid systems, FMN dimers emit fluorescence [
7,
21,
24]. In systems where dimers emit fluorescence, one observes not only multistep energy migration between monomers (M) and forward energy transfer (FET) from M to D, but also reverse nonradiative excitation energy transfer (RET) from FMN dimers to monomers, as well as energy migration between dimers [
25,
26]. Qualitative and quantitative investigations of nonradiative energy transfer (NET) for FMN in a glycerol-water solution [
22,
27] and rigid (PVA) solutions [
25,
26] have been performed using the Bojarski theory [
28,
29] of multistep excitation forward and reverse energy transport (hopping model). Importantly, it has been shown that FMN molecules do not create higher order associates, only dimers [
20,
30,
31,
32]. This makes it possible to investigate the energy transfer processes between monomers and dimers, even in systems of high FMN concentration, leading to unambiguous results.
The FMN dimer structure [
21,
32] was determined based on spectroscopic studies and the Kasha model [
33]. The FMN monomer units in a “stacked” dimer are oriented face to face, and the angle between transition moments
and
(corresponding to S
1→S
0 transition) in monomer units in dimer is α = 71 ± 4° [
21,
26]. Such a structure, according to the Kasha theory [
33], enforces the existence of two exciton absorption levels, H and J, of the excited state of the dimer. According to Kasha’s exciton theory [
33], the splitting of energetic state of the monomer into two E
H and E
J energetic states in the dimer appears because of a dipole–dipole interaction between monomer units. If the transition moments
and
of S
1→ S
0 in two monomers in the dimer form an angle α ≠ 0° and α ≠ 180°, the transition to both H and J exciton states is allowed, and two bands, H and J, appear in the dimer absorption spectrum. The FMN dimer absorption spectrum in PVA was resolved [
21,
26] into the component H and J bands [
33].
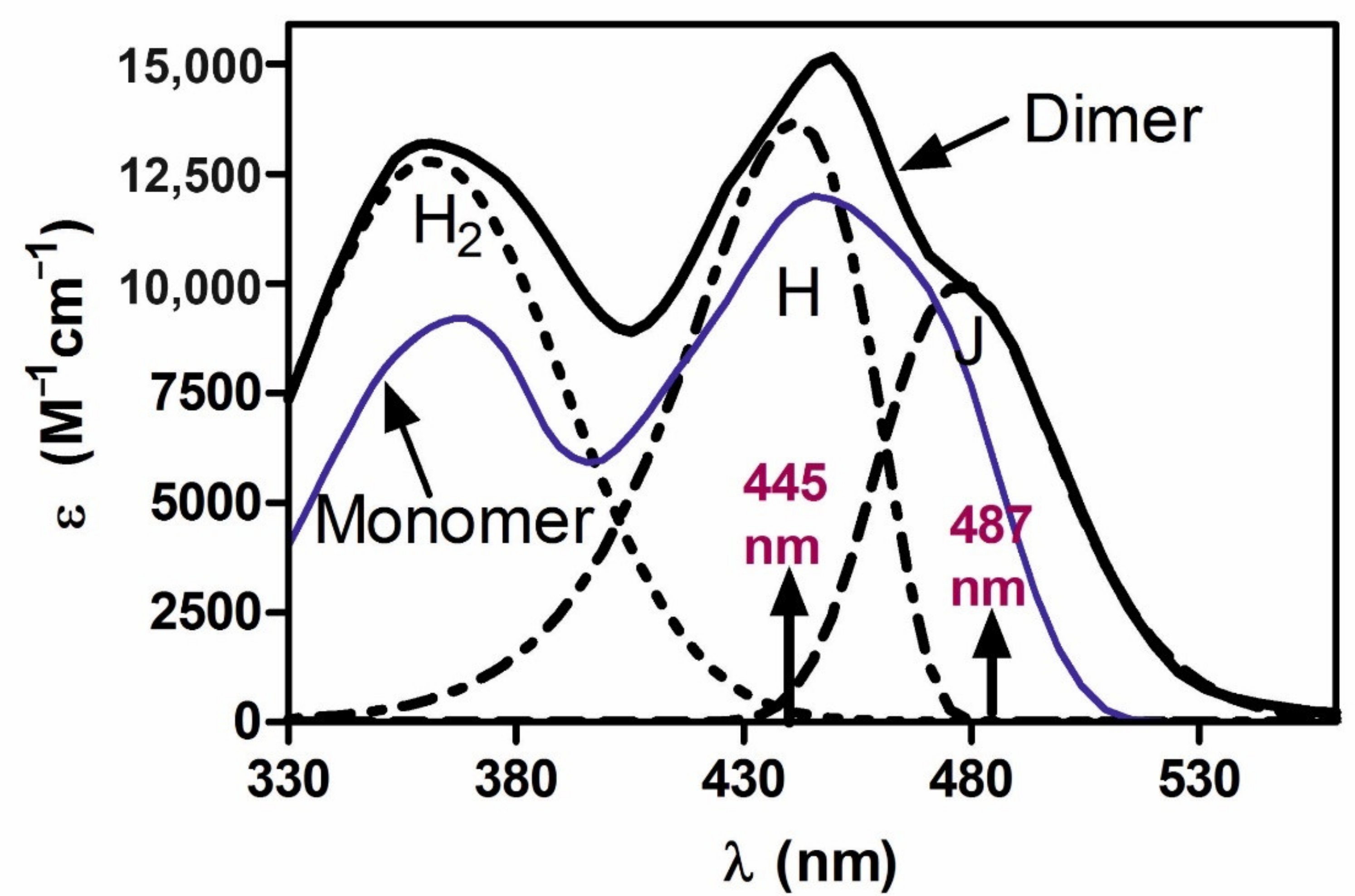
Figure 1 shows H and J components of the dimer absorption spectrum of FMN in PVA films, according to Grajek et al. [
21] and Grajek [
26]. A question then arises as to whether the two energy levels of the H and J dimer states would not affect the course of photoreception, given that two quants transfer the dimer to the reactive excited state.
The existence of two dimer states ought to be reflected in fluorescence spectra. The shape of a fluorescence spectrum and its overlapping with an absorption spectrum will significantly affect the probability of a nonradiative energy transfer.
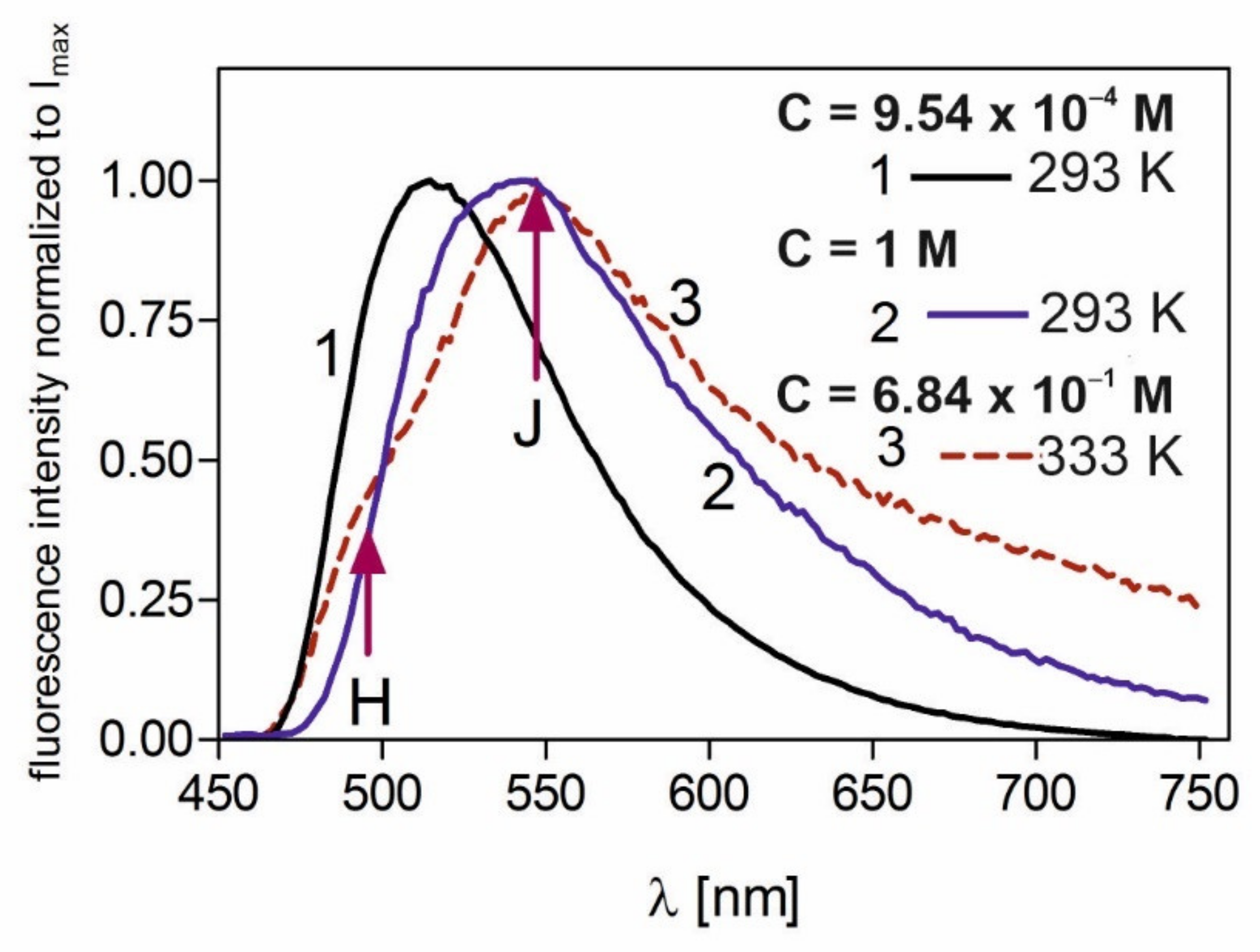
Figure 2 shows the fluorescence spectra of FMN in PVA for low concentration—C = 9.54 × 10
−4 M (curve 1)—and extremely high concentrations: C = 1 M at T = 293 K (curve 2), and C = 6.84 × 10
−1 M at T = 333 K (curve 3).
For an extremely high concentrations of FMN, at λ
exc = 445 nm, (curves 2 and 3,
Figure 2), the maximum of fluorescence band shifts to 544 ± 2 nm [
21,
26,
35]. It was proven [
21,
34] that this is due to the fluorescent dimers of FMN, the amount of which is high in this concentration (reaching 80%). The steady-state fluorescence spectra for highly concentrated samples are wide, and fluorescence from dimer H and J levels is not indistinguishable (see curve 2,
Figure 2), except for one case in which fluorescence from the H dimer level is revealed at a high temperature—curve 3
Figure 2. This study employed a time-resolved technique to reveal and confirm the fluorescence derived from two dimer states, from which energy transfer of quants of different energies is possible. The study’s other aims included investigating the effect of energy transfer between monomers and dimers (when dimers have own fluorescence) on the fluorescence decays of FMN in PVA, examining fluorescence decays coming from both H and J dimer states, and establishing their fluorescence lifetimes using time-resolved fluorescence techniques. Thus far, for any dye-forming dimers, it was not possible to observe dimer fluorescence alone; this was due to the fact that these compounds formed higher aggregates with increases in concentration. As molecules of FMN do not form higher aggregates but only dimers [
30,
32,
34], we examined samples of extremely high concentrations. In this way, it was possible to investigate the fluorescence decays coming from the H and J dimer states of FMN.
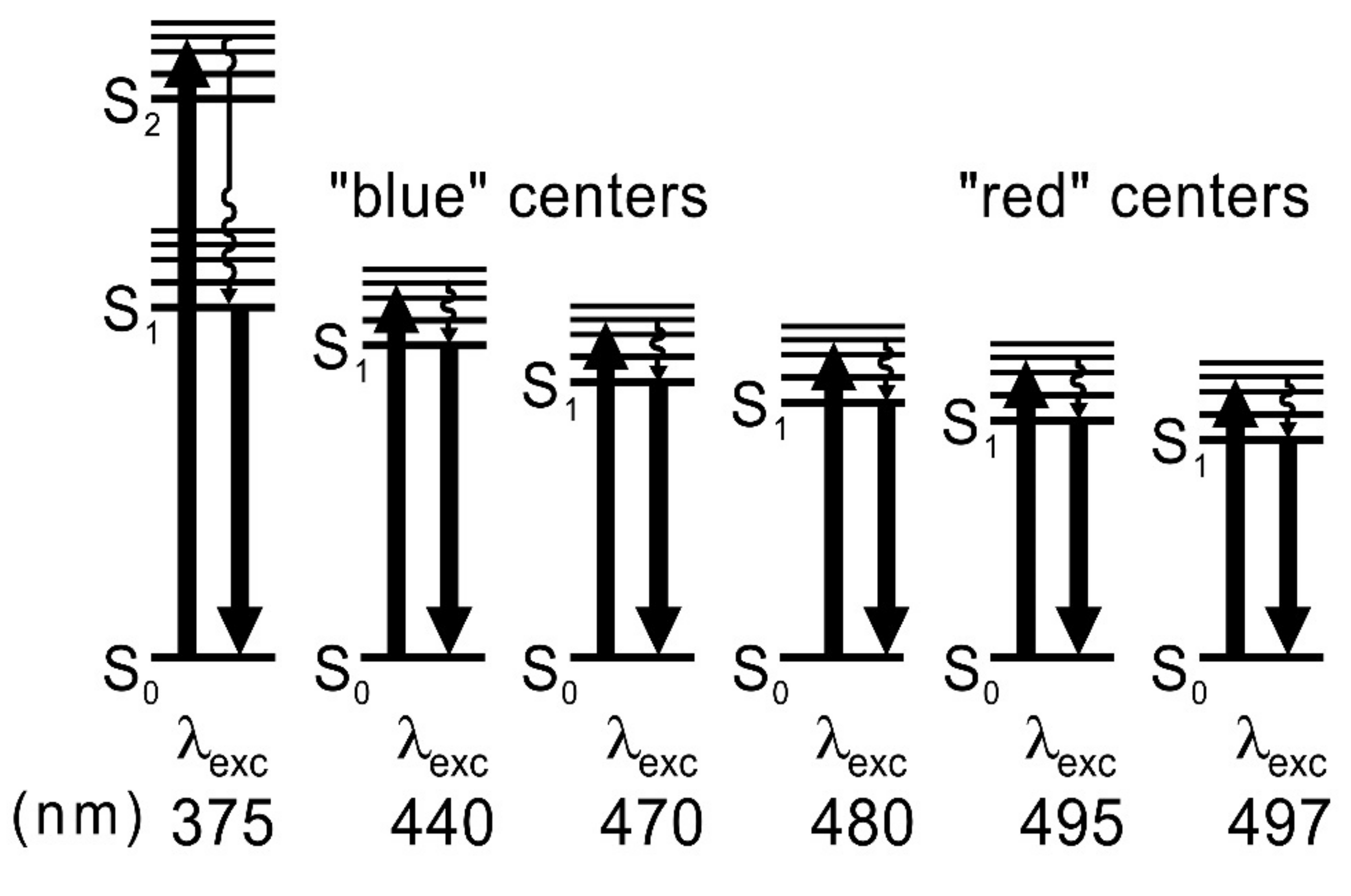
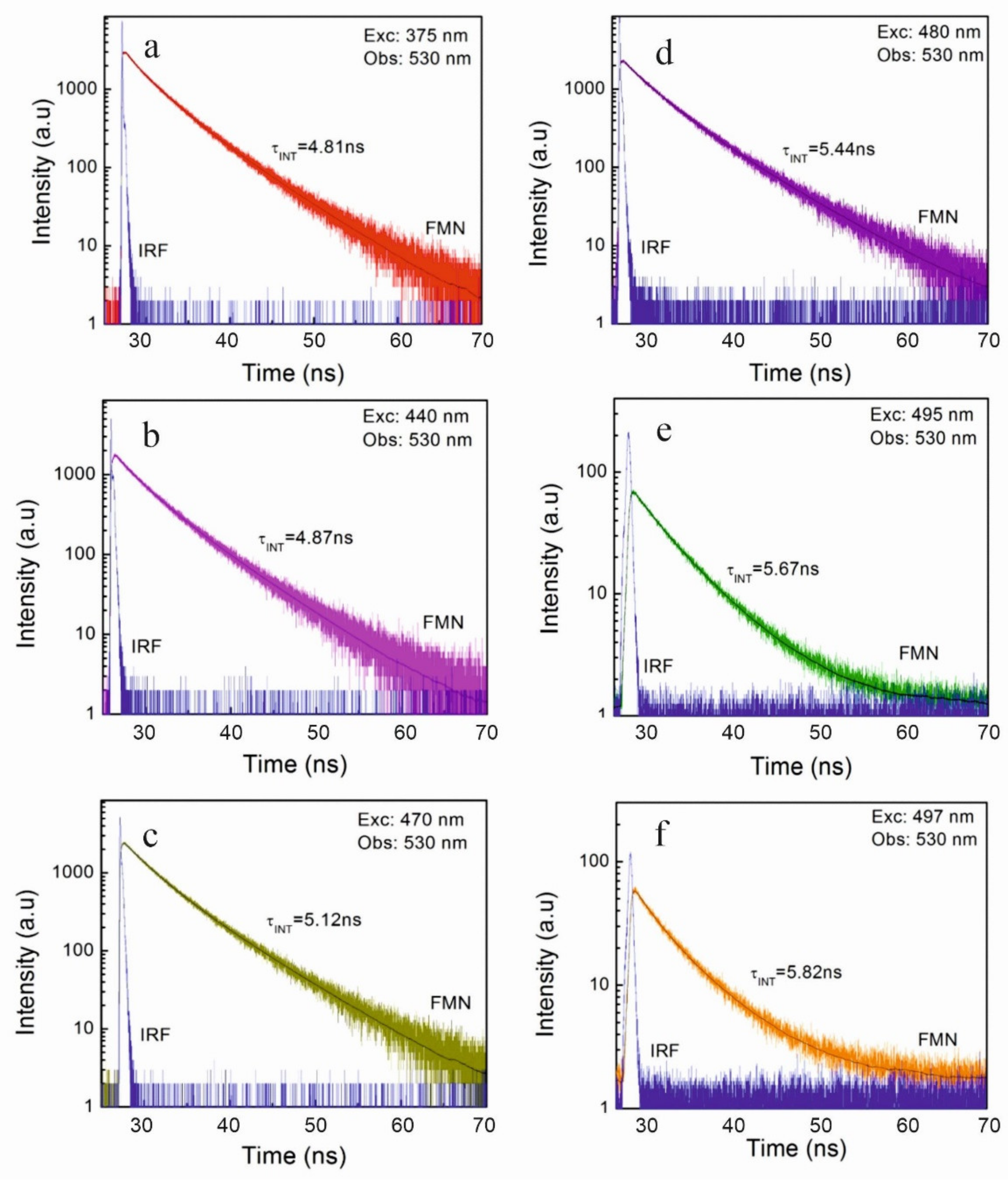
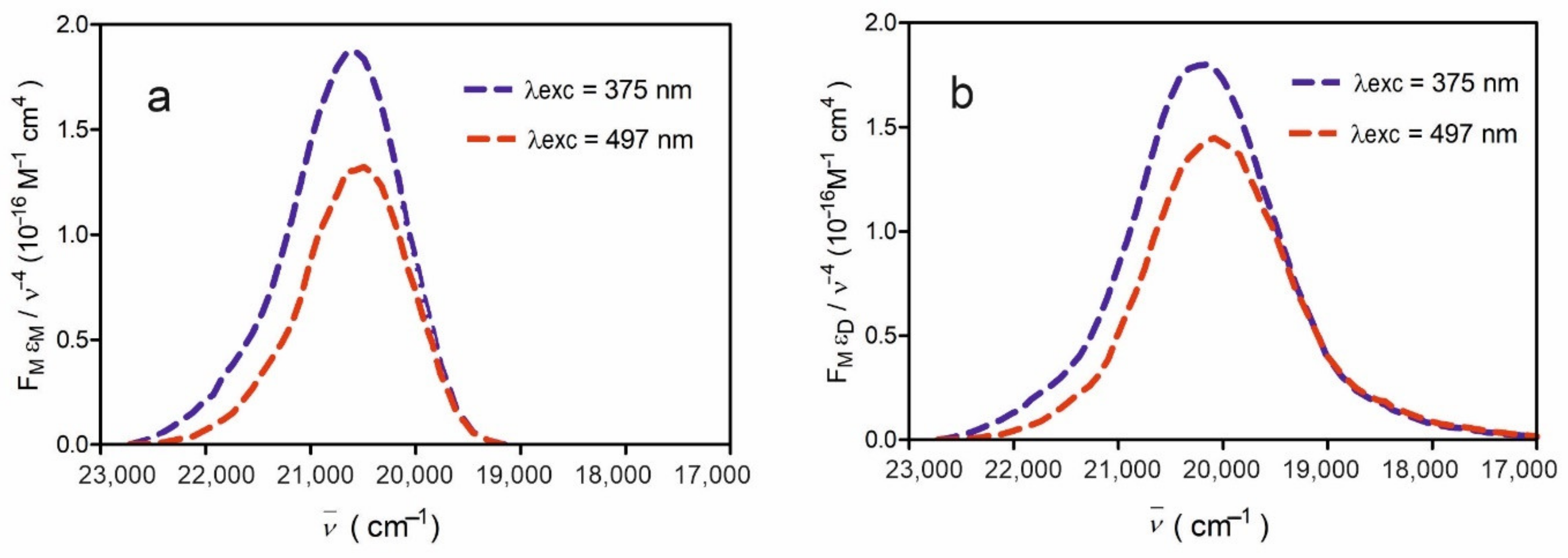
An interesting phenomenon has been observed at low dye concentrations in a rigid environment upon excitation by different wavelengths. It has been shown [
21,
26,
35] that, at low FMN concentrations in a rigid PVA, “blue” and “red” fluorescence centers appear, and the fluorescence maximum shifts from 512 nm to 526 nm with the change in the excitation wavelength from λ
exc = 445 nm to 490 nm; this is because of the phenomenon of inhomogeneous orientational broadening of dye energy levels (IOBEL) [
36,
37]. IOBEL results from the existence of a certain distribution of the 0–0 transition energies caused by different interaction energies between the fluorescent molecule and its environment—the polymer matrix. The “blue” and “red” fluorescent centers for FMN in PVA were, up until now, investigated using the steady-state fluorescence technique [
21,
26,
35]. In this work, we have undertaken broader research into these centers using a time-resolved technique, and we have investigated their impact on energy transfer between flavins.
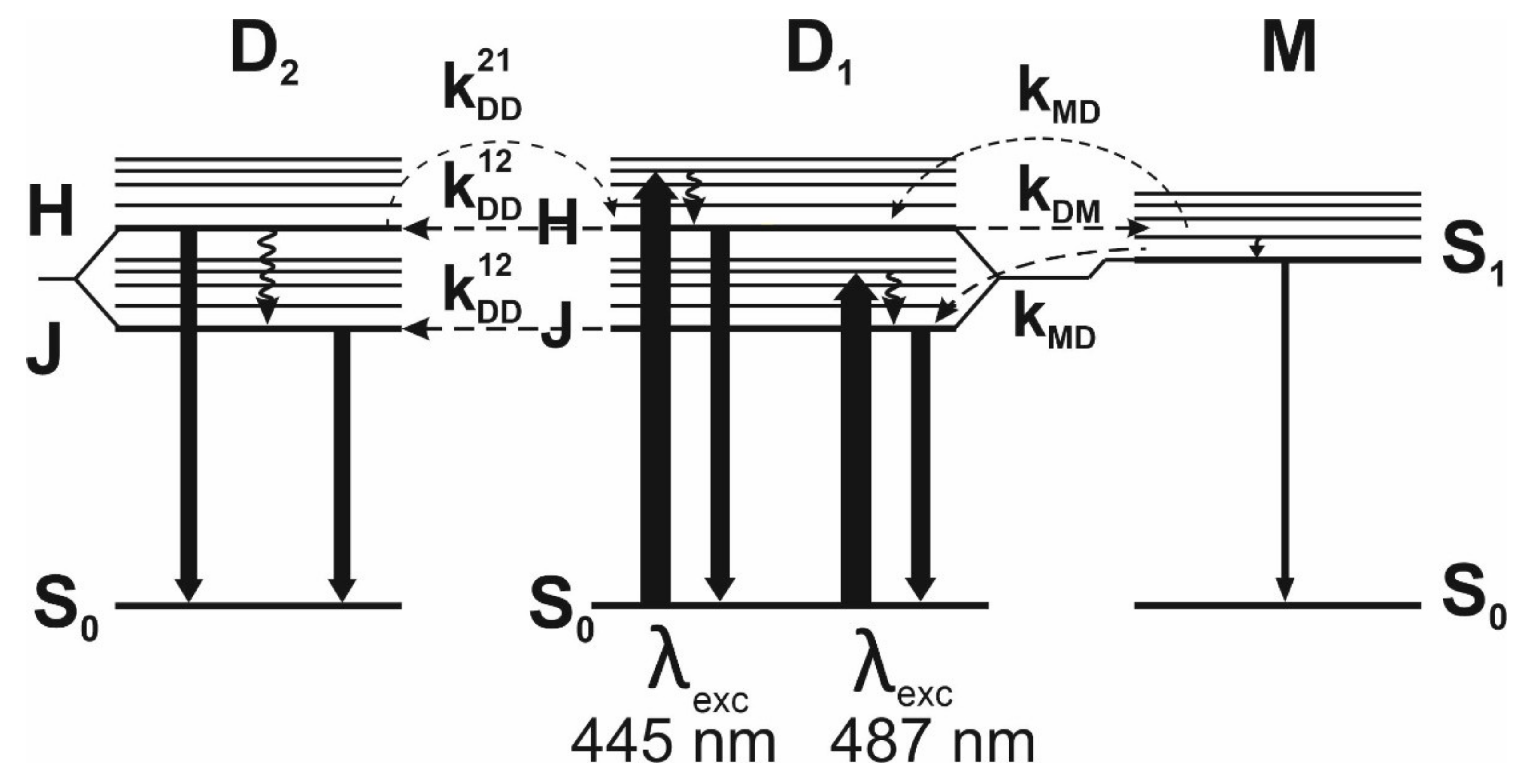
While studying photoreception cycles with FMN or FAD transformations, especially including flavins incorporated into the protein pocket of an RoLOV1 photoreceptor, some authors [
5] have obtained shifted fluorescence spectra at various excitation waves. However, they have taken no account of the shifts stemming from the IOBEL effect (with orientational interaction of flavin in a rigid protein pocket). Goett-Zink et al. [
38] have demonstrated that the intracellular environment affects LOV photoreceptor mechanisms in general. Moreover, in-cell fluorescence spectroscopy has found that the intracellular environment slows down recovery of the light-induced flavin adduct.
Our study into the effect of interactions among flavins with a stiffened environment on their fluorescence may contribute to innovative interpretations of phenomena occurring in biological systems examined by spectroscopic methods. It is therefore important to perform in vitro studies into the impact of the stiffened environment on flavin spectroscopy, particularly flavin fluorescence. The stiffened environment can be a (simple) model of molecules entrapped in rigid protein structures in plant organisms. It is possible that the stiffening of flavin, e.g., in a protein pocket, leads to the emergence of various FMN fluorescence centers and enables the observation of fluorescence coming from these centers. Their appearance will entail changes in the energy of the light quantum transfer and affect the yield of energy transfer in photoreception phenomena because different energy quanta could then be transferred, depending on flavin positioning, against the protein matrix. This may alter the interpretation of many phenomena occurring in biological systems that have been examined by spectroscopic methods. For instance, it seems interesting which centers will facilitate a more effective energy transfer for FMN. Hence, we have undertaken a study in the model system in which FMN is embedded in rigid PVA in order to determine the magnitude of changes induced in the values of FMN fluorescence energy quanta by the IOBEL phenomenon, as well as to establish the greatest likelihood of energy transfer.
The next aim of the study was to investigate the fluorescence decays coming from individual fluorescence centers, from “blue” to “red”, and examine the influence of the phenomenon of inhomogeneous orientational broadening of energy levels on the lifetimes of fluorescence from lower and lower FMN centers. Such fluorescent centers may appear in biological systems (in photoreceptors), where FMN molecules are built in protein pockets, as well as in other compounds such as chlorophylls.
3. Materials and Methods
Flavin mononucleotide FMN (riboflavin-5′-monophosphate, sodium salt (C17H20N4NaO9P·2H2O) of analytical purity was purchased from Fluka Chemie AG, Buchs, Switzerland. Polyvinyl alcohol of spectroscopic grade was obtained from Loba-Chemie, Vien-Fishamend, Austria. Homogeneous 10% aqueous solution of PVA at a temperature of about 350 K was prepared. FMN was added to this solution and carefully dissolved. The FMN concentrations in PVA films varied from C = 9.5 × 10−5 M to C = 6.8 × 10−1 M. Samples of final concentration C < 10−2 M were prepared by careful distribution of small amounts of solution on horizontally placed polished glass plates, followed by water evaporation. The samples of concentrations C > 10−2 M were obtained by dip coating method and left for slow water evaporation. This method allows one to obtain the samples of such a thickness that the optical density is below 0.1 and the inner filter effects are insignificant.
The fluorescence intensity decays of FMN-doped PVA films were studied by the means of the time-correlated single photon counting (TCSPC) technique. Two separate experimental setups were used. The first one, with an FWHM of IRF of about 35 ps, was custom-built by Lorenc et al. [
52]. The second, with FWHM of IRF of about 70 ps, was the FluoTime 200 (PicoQuant, GmbH, Berlin, Germany) time-resolved spectrofluorometer. This instrument contains a multichannel plate detector (Hamamatsu, Japan). The fluorescence intensity decays were measured in magic angle conditions and data were analyzed with FluoFit version 4.5.3 software (PicoQuant GmbH, Berlin, Germany). For each experimental condition (concentration, temperature, excitation wavelength, and emission wavelength) three decays were collected by TCSPC technique. Additionally, for each decay, the fits were performed for at least five different sets of starting parameters to make sure that the global minimum was found by the fitting procedure (Levenberg–Marquardt algorithm). The fitting software, apart from the best parameters of the fit, provided the standard deviation of the fitted parameters. The decays were accumulated to 10,000 counts in a maximum in 1024 channels. For such an accumulation, the typical ratio of the standard deviation to its parameter is equal to 2%, while this ratio increases to 4% for smaller accumulations such as 1000 at its maximum. The quality of the fit was judged by the value of a χ
2 test and the distribution of weighed residuals. The full width at half maximum (FWHM) of IRF was 35 ps or 70 ps, which was much lower than the typical time constants obtained in the paper. Laser pulse duration was about 1.5 ps, and it was stable during the experiment. The theoretically calculated range of χ
2 values in which the fit can be assumed to be correct is from 0.80 to 1.20 (
p = 0.05) [
53].
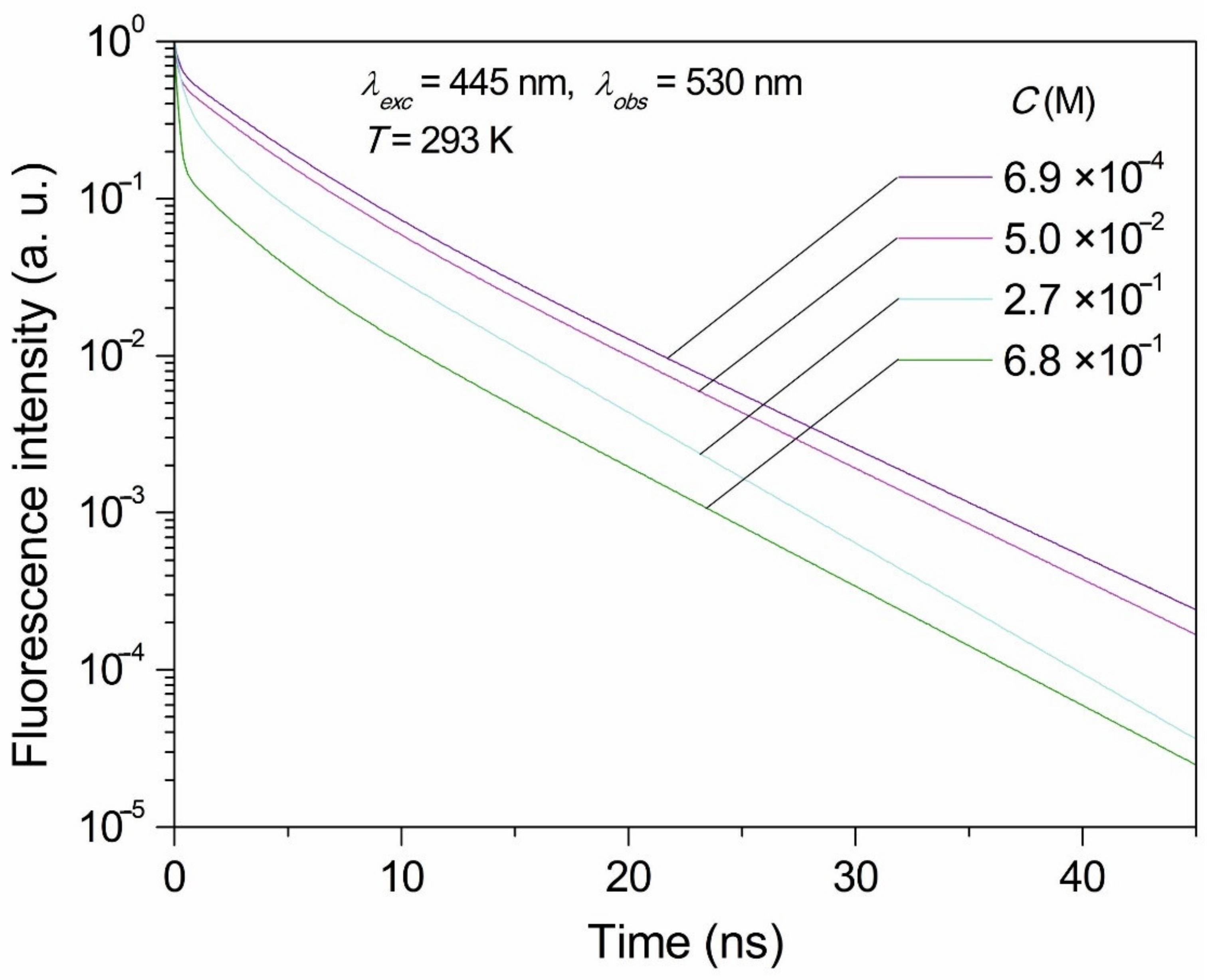
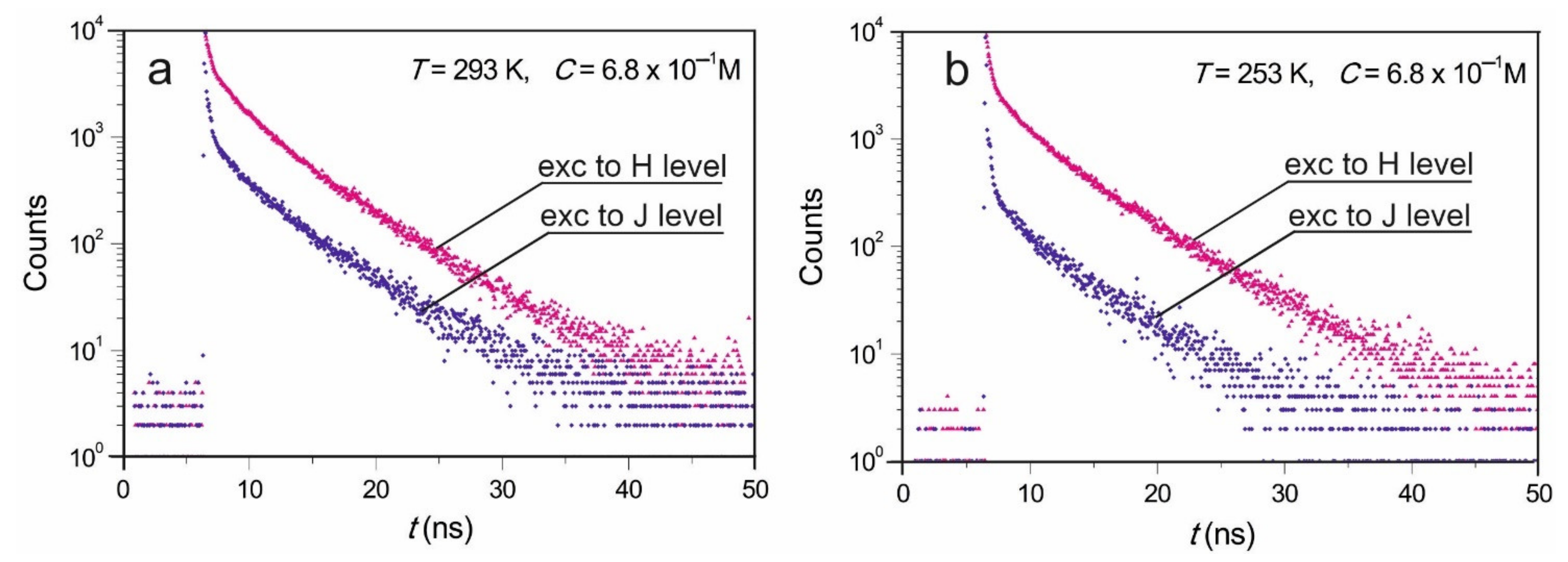
Excitations at 375 nm, 440 nm, 445 nm, 470 nm, 480 nm, 487 nm, 495 nm, and 497 nm were applied, and emissions at the 530 nm and the 600 nm were observed. The different wavelength of excited light was applied to observe the IOBEL effect. Concentration dependence of FMN fluorescence decays was performed at excitations at 445 nm and observations were performed at 530 nm. With both setups, consistent results were obtained. Emissions were observed at room temperature, and some results were collected at 253 K using Oxford N2 cryostat.
The analysis of fluorescence intensity decays was carried out using Fluofit software (Picoquant), and was also based on home-built software [
53]. A multiexponential fluorescence decay model was employed and standard procedures for judging the quality of the fit were applied [
53].
Concentrations of monomers,
, and dimers,
, for samples of given concentration
were calculated from the formulas:
K is the dimerization constant, which was calculated on the base on (of) the modified Förster–Levshin method [
54,
55]. This method was used to calculate pure dimer
and monomer
spectra, as well as the value of K = 11.6 M
−1 for FMN [
21].
The contributions of dimers
to the total absorption of the sample have been obtained from the following formula:
where
and
denote the extinction coefficients of monomers and dimers at a given excitation wavelength
.
The Förster radii for energy transfer were calculated from the well-known formula [
56].
where
n denotes the refractive index of the medium,
N—the Avogadro’s number,
η0M—the absolute fluorescence quantum yield of the donor (monomer) in the absence of any energy transfer processes,
—the averaged orientation factor dependent on the mutual molecular alignment, and
IMX—the overlap integral of the donor fluorescence spectrum
(monomer) with the acceptor absorption spectrum of the monomer
or dimer
:
where the donor emission spectral distribution is normalized:
.
Fluorescence decays have been analyzed using a multiexponential approximation
where
.
Based on this manner of evaluated values of
and
the average lifetime of fluorescence
has been calculated using the formulae [
57]:
The degree of nonexponentiality of decay can be characterized by parameter
, which is defined as [
58]:
It is clear from Equation (10) that describes the difference between the considered decay and the pure exponential decay of the same area under the decay curve. The normalization factor, , ensures that all possible values of are limited to the interval {0, 1}. If the fluorescence decay is exponential, then its degree of nonexponentiality is equal to zero.
4. Conclusions
The examination of fluorescence decays clearly shows that NET processes and dimer formation with increasing concentration of FMN greatly shortens the fluorescence decay time of FMN samples in concentrations ranging from 6.9 × 10−4 M to 6.8 × 10−1 M.
Due to the fact that we managed to achieve a high concentration of FMN samples at which the concentration of dimers reached 80%, and the molecules of FMN did not form higher associates but only dimers, it was possible to examine the fluorescence decays from H and J dimer levels. The mean fluorescence times from the dimer levels were calculated.
FMN in biological systems form dimers [
17,
18,
19,
59]. Knowledge of fluorescence originating from both exciton levels of dimers may open the way for further studies of dimers’ spectroscopy in vivo, as well as energy transfer between FMN dimers in more complex systems.
Fluorescence decays studies in rigid systems have clearly shown that the IOBEL phenomenon cannot be omitted in the interpretation of fluorescence spectra and time-resolved experimental results, because IOBEL causes a deviation in the decay curve from one-exponential character to a multi-exponential character; moreover, the mean values of lifetimes depend on excitation wavelengths. The IOBEL phenomenon may also have an effect on the transfer of excitation energy primarily absorbed by plants, where photoreceptors are rigidified in biological structures.
The environment may have a significant impact on the operation of LOV flavin photoreceptors. In this work, we managed to show the influence of the stiffened environment (e.g., protein) and the mutual orientation of FMN molecules with respect to the polymer matrix on fluorescence characteristics. Depending on the positioning of FMN molecules, different local interactions of molecules with the environment (the so-called IOBEL centers) exist, which affects the results of spectroscopic research. Considering the IOBEL phenomenon in study design can provide more complete information in the research of photoreceptors and signal transduction in photoreceptor processes. Many other biological compounds found in cell membranes, such as proteins, phytosterols, chlorophylls, ion channels, etc., are stiffened, and the IOBEL phenomenon can also cause changes in their spectroscopic properties. Therefore, it is important to take this phenomenon into account. It should be noted that spectroscopic methods are commonly used to analyze biological compounds.


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}