
LncRNA MORT (ZNF667-AS1) in Cancer—Is There a Possible Role in Gynecological Malignancies?

 ,
,  , , , , ,
, , , , ,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. LncRNA MORT (ZNF667-AS1): From Discovery to Function
3. The Functional Role of LncRNA MORT (ZNF667-AS1) in Human Diseases
4. Dysfunction of LncRNA MORT (ZNF667-AS1) in Gynecological Cancers
4.1. Ovarian Cancer
4.2. Cervical Cancer
4.3. Endometrial Cancer
5. Epigenetic Silencing of LncRNA MORT (ZNF667-AS1) in Cancer
6. Epigenetic and Gene Therapies for Gynecological Cancer Treatment
7. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AML | acute myeloid leukemia |
| CC | cervical cancer |
| CRC | colorectal carcinoma |
| EC | endometrial cancer |
| ESCC | esophageal squamous cell carcinoma |
| HCC | hepatocellular carcinoma |
| HPV | human papillomavirus |
| IRLs | immune-related lncRNAs |
| LncRNA | long non-coding RNA |
| LSCC | laryngeal squamous cell carcinoma |
| LUAD | lung adenocarcinoma |
| miRNA/miR | micro-RNA |
| MORT | mortal obligate RNA transcript |
| ncRNA | non-coding RNA |
| NPC | nasopharyngeal carcinoma |
| OC | ovarian cancer |
| OSCC | oral squamous cell carcinoma |
| PAP | Papanicolaou |
| RGCs | rare gynecologic cancers |
| UCS | uterine carcinosarcoma |
| UM | uveal melanoma |
| ZNF667-AS1 | ZNF667 antisense RNA 1 (head to head) |
References
- Srivastava, S.K.; Ahmad, A.; Zubair, H.; Miree, O.; Singh, S.; Rocconi, R.P.; Scalici, J.; Singh, A.P. MicroRNAs in gynecological cancers: Small molecules with big implications. Cancer Lett. 2017, 407, 123–138. [Google Scholar] [CrossRef]
- Liu, D.T. EMMPRIN in gynecologic cancers: Pathologic and therapeutic aspects. Tumour Biol. 2015, 36, 4883–4888. [Google Scholar] [CrossRef]
- Maheshwari, A.; Kumar, N.; Mahantshetty, U. Gynecological cancers: A summary of published Indian data. South. Asian J. Cancer 2016, 5, 112–120. [Google Scholar] [CrossRef]
- Mandilaras, V.; Karakasis, K.; Clarke, B.; Amit Oza, A.; Lheureux, S. Rare tumors in gynaecological cancers and the lack of therapeutic options and clinical trials. Expert Opin. Orphan Drugs 2017, 5, 71–83. [Google Scholar] [CrossRef]
- Ray-Coquard, I.; Trama, A.; Seckl, M.J.; Fotopoulou, C.; Pautier, P.; Pignata, S.; Kristensen, G.; Mangili, G.; Falconer, H.; Massuger, L.; et al. Rare ovarian tumours: Epidemiology, treatment challenges in and outside a network setting. Eur. J. Surg. Oncol. 2017, 45, 67–74. [Google Scholar] [CrossRef]
- Di Fiore, R.; Suleiman, S.; Ellul, B.; O’Toole, S.A.; Savona-Ventura, C.; Felix, A.; Napolioni, V.; Conlon, N.T.; Kahramanoglu, I.; Azzopardi, M.J.; et al. GYNOCARE Update: Modern Strategies to Improve Diagnosis and Treatment of Rare Gynecologic Tumors-Current Challenges and Future Directions. Cancers 2021, 13, 493. [Google Scholar] [CrossRef]
- Cancer Research UK. Available online: https://www.cancerresearchuk.org (accessed on 20 February 2021).
- Rare Cancers Europe. Available online: https://www.rarecancerseurope.org (accessed on 17 February 2021).
- Andersen, R.E.; Lim, D.A. Forging our understanding of lncRNAs in the brain. Cell Tissue Res. 2017, 371, 55–71. [Google Scholar] [CrossRef]
- Isin, M.; Dalay, N. LncRNAs and neoplasia. Clin. Chim. Acta 2015, 444, 280–288. [Google Scholar] [CrossRef]
- Yin, Y.; Yangyang, Z.; Lu, J.; Song, G.; Zhu, Y.; Li, Z.; Zhaohui, L.; Shen, B.; Huang, X.; Zhu, H.; et al. Opposing Roles for the lncRNA Haunt and Its Genomic Locus in Regulating HOXA Gene Activation during Embryonic Stem Cell Differentiation. Cell Stem Cell 2015, 16, 504–516. [Google Scholar] [CrossRef] [Green Version]
- Blokhin, I.; Khorkova, O.; Hsiao, J.; Wahlestedt, C. Developments in lncRNA drug discovery: Where are we heading? Expert Opin. Drug Discov. 2018, 13, 837–849. [Google Scholar] [CrossRef]
- Tornesello, M.L.; Faraonio, R.; Buonaguro, L.; Annunziata, C.; Starita, N.; Cerasuolo, A.; Pezzuto, F.; Tornesello, A.L.; Buonaguro, F.M. The Role of microRNAs, Long Non-coding RNAs, and Circular RNAs in Cervical Cancer. Front. Oncol. 2020, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Zhang, J.; Wei, M.; Liu, B.; Dong, K. Emerging role of long noncoding RNA-encoded micropeptides in cancer. Cancer Cell. Int. 2020, 20, 506. [Google Scholar] [CrossRef]
- Patty, B.J.; Hainer, S.J. Non-Coding RNAs and Nucleosome Remodeling Complexes: An Intricate Regulatory Relationship. Biology 2020, 9, 213. [Google Scholar] [CrossRef]
- Yao, R.W.; Wang, Y.; Chen, L.L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Zhu, Y.; Luo, M.; Brooks, M.; Clouthier, S.G.; Wicha, M.S. Biological and clinical significance of cancer stem cell plasticity. Clin. Transl. Med. 2014, 3, 32. [Google Scholar] [CrossRef] [Green Version]
- Ørom, U.A.; Shiekhattar, R. Long noncoding RNAs usher in a new era in the biology of enhancers. Cell 2013, 154, 1190–1193. [Google Scholar] [CrossRef] [Green Version]
- Wahlestedt, C. Targeting long non-coding RNA to therapeutically upregulate gene expression. Nat. Rev. Drug Discov. 2013, 12, 433–446. [Google Scholar] [CrossRef]
- Vrba, L.; Garbe, J.C.; Stampfer, M.R.; Futscher, B.W. A lincRNA connected to cell mortality and epigenetically-silenced in most common human cancers. Epigenetics 2015, 10, 1074–1083. [Google Scholar] [CrossRef] [Green Version]
- Li, J.W.; Kuang, Y.; Chen, L.; Wang, J.F. LncRNA ZNF667-AS1 inhibits inflammatory response and promotes recovery of spinal cord injury via suppressing JAK-STAT pathway. Eur. Rev. Med. Pharm. Sci. 2018, 22, 7614–7620. [Google Scholar]
- Peffers, M.J.; Fang, Y.; Cheung, K.; Wei, T.K.; Clegg, P.D.; Birch, H.L. Transcriptome analysis of ageing in uninjured human Achilles tendon. Arthritis Res. 2015, 17, 33. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, Y.; Jiang, H.; Jiang, N.; Gao, J. Integrative analysis of the contribution of mRNAs and long non-coding RNAs to the pathogenesis of asthma. Mol. Med. Rep. 2019, 20, 2617–2624. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zhu, Y.; Yao, S. LncRNAMORT is upregulated in myocardial infarction and promotes the apoptosis of cardiomyocyte by downregulating miR-93. BMC Cardiovasc. Disord. 2020, 20, 247. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, Y.; Zheng, P.; Cai, C.; Jiang, Y.; Zhang, H.; Li, Z.; Cai, Q. Long non-coding RNAs mortal obligate RNA transcript regulates the proliferation of human periodontal ligament stem cells and affects the recurrence of periodontitis. Arch. Oral. Biol. 2019, 105, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Cui, W.; Zhao, L.; Chi, W.; Cao, H.; Wang, B. Aberrant methylation and downregulation of ZNF667-AS1 and ZNF667 promote the malignant progression of laryngeal squamous cell carcinoma. J. Biomed. Sci. 2019, 26, 13. [Google Scholar] [CrossRef]
- Jin, Z.; Jiang, S.; Jian, S.; Shang, Z. Long noncoding RNA MORT overexpression inhibits cancer cell proliferation in oral squamous cell carcinoma by downregulating ROCK1. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Huang, Y.; Shi, D.; Nie, C.; Luo, Y.; Guo, L.; Zou, Y.; Xie, C. LncRNA ZNF667-AS1 Promotes ABLIM1 Expression by Adsorbing microRNA-1290 to Suppress Nasopharyngeal Carcinoma Cell Progression. Onco Targets Ther. 2020, 13, 4397–4409. [Google Scholar] [CrossRef]
- Dong, Z.; Li, S.; Wu, X.; Niu, Y.; Liang, X.; Yang, L.; Guo, Y.; Shen, S.; Liang, J.; Guo, W. Aberrant hypermethylation-mediated downregulation of antisense lncRNA ZNF667-AS1 and its sense gene ZNF667 correlate with progression and prognosis of esophageal squamous cell carcinoma. Cell Death Dis. 2019, 10, 930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Cai, M.Y.; Rong, H.; Ma, L.R.; Xu, Y.L. ZNF667-AS1, a positively regulating MEGF10, inhibits the progression of uveal melanoma by modulating cellular aggressiveness. J. Biochem. Mol. Toxicol. 2021, e22732. [Google Scholar]
- Huang, Y.; Zhuang, Q.; Zhuang, W. Mortal Obligate RNA Transcript Inhibits Cancer Cell Invasion and Migration in Lung Adenocarcinoma by Downregulating miRNA-223. Cancer Biother. Radiopharm. 2020, 35, 345–350. [Google Scholar] [CrossRef]
- Zhuang, L.; Ding, W.; Ding, W.; Zhang, Q.; Xu, X.; Xi, D. lncRNA ZNF667-AS1 (NR_036521.1) inhibits the progression of colorectal cancer via regulating ANK2/JAK2 expression. J. Cell. Physiol. 2021, 236, 2178–2193. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wu, L.; Zong, Z.; Ma, N.; Li, Y.; Jiang, Z.; Wang, Q.; Chen, S. Long non-coding RNA mortal obligate RNA transcript inhibits the migration and invasion of colon cancer cells by inactivating transforming growth factor β1. Oncol. Lett. 2020, 19, 1131–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sun, Q.; Ji, L.; Wang, G.; Niu, X.; Sun, S. lncRNA MORT Regulates Bladder Cancer Behaviors by Downregulating MicroRNA-146a-5p. Nephron 2020, 144, 351–357. [Google Scholar] [CrossRef]
- Lu, X.; Ge, G.; Ji, F.; Wang, J. LncRNA MORT Overexpression Inhibits Cancer Cell Migration and Invasion in Hepatocellular Carcinoma by Downregulating NOTCH1. Cancer Biother. Radiopharm. 2021. [Google Scholar] [CrossRef]
- Wang, N.; Feng, Y.; Xie, J.; Han, H.; Dong, Q.; Wang, W. Long Non-Coding RNA ZNF667-AS1 Knockdown Curbs Liver Metastasis in Acute Myeloid Leukemia by Regulating the microRNA-206/AKAP13 Axis. Cancer Manag. Res. 2020, 12, 13285–13300. [Google Scholar] [CrossRef]
- Yuan, Q.; Gao, C.; Lai, X.D.; Chen, L.Y.; Lai, T.B. Analysis of Long Noncoding RNA ZNF667-AS1 as a Potential Biomarker for Diagnosis and Prognosis of Glioma Patients. Dis. Markers 2020, 2020, 8895968. [Google Scholar] [CrossRef]
- Hosseini, E.S.; Meryet-Figuiere, M.; Sabzalipoor, H.; Kashani, H.H.; Nikzad, H.; Asemi, Z. Dysregulated expression of long noncoding RNAs in gynecologic cancers. Mol. Cancer 2017, 16, 107. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wu, W.; Cao, X.; Zhao, X.; Li, W.; Deng, C.; Huang, Z. lncRNA mortal obligate RNA transcript was downregulated in ovarian carcinoma and inhibits cancer cell proliferation by downregulating miRNA-21. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef]
- Li, Y.J.; Yang, Z.; Wang, Y.Y.; Wang, Y. Long noncoding RNA ZNF667-AS1 reduces tumor invasion and metastasis in cervical cancer by counteracting microRNA-93-3p-dependent PEG3 downregulation. Mol. Oncol. 2019, 13, 2375–2392. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.P.; Li, R.H.; Han, D.M.; Zhang, X.Q.; Nian, G.X.; Wu, M.X.; Feng, Y.; Zhang, L.; Sun, Z.G. Independent prognostic Factor of low-expressed LncRNA ZNF667-AS1 for cervical cancer and inhibitory function on the proliferation of cervical cancer. Eur. Rev. Med. Pharm. Sci. 2017, 21, 5353–5360. [Google Scholar]
- Zheng, J.; Cao, B.; Zhang, X.; Niu, Z.; Tong, J. Immune-Related Four-lncRNA Signature for Patients with Cervical Cancer. Biomed. Res. Int. 2020, 2020, 3641231. [Google Scholar] [CrossRef]
- Vrba, L.; Futscher, B.W. Epigenetic silencing of lncRNA MORT in 16 TCGA cancer types. F1000Research 2018, 7, 211. [Google Scholar] [CrossRef] [Green Version]
- Coburn, S.B.; Bray, F.; Sherman, M.E.; Trabert, B. International patterns and trends in ovarian cancer incidence, overall and by histologic subtype. Int. J. Cancer 2017, 140, 2451–2460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoneda, A.; Lendorf, M.E.; Couchman, J.R.; Multhaupt, H.A. Breast and ovarian cancers: A survey and possible roles for the cell surface heparan sulfate proteoglycans. J. Histochem. Cytochem. 2012, 60, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, I.J.; Menon, U. Progress and challenges in screening for early detection of ovarian cancer. Mol. Cell Proteom. 2004, 3, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Badgwell, D.; Bast, J.R.C. Early detection of ovarian cancer. Dis. Markers 2007, 23, 397–410. [Google Scholar] [CrossRef] [Green Version]
- Lowe, K.A.; Chia, V.M.; Taylor, A.; O’Malley, C.; Kelsh, M.; Mohamed, M.; Mowat, F.S.; Goff, B. An international assessment of ovarian cancer incidence and mortality. Gynecol. Oncol. 2013, 130, 107–114. [Google Scholar] [CrossRef]
- Hunn, J.; Rodriguez, G.C. Ovarian cancer: Etiology, risk factors, and epidemiology. Clin. Obs. Gynecol. 2012, 55, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Trabert, B.; DeSantis, C.E.; Miller, K.D.; Samimi, G.; Runowicz, C.D.; Gaudet, M.M.; Jemal, A.; Siegel, R.L. Ovarian cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Mota, A.; S Oltra, S.; Moreno-Bueno, G. Insight updating of the molecular hallmarks in ovarian carcinoma. Eur. J. Cancer Suppl. 2020, 15, 16–26. [Google Scholar] [CrossRef]
- Vaughan, S.; Coward, J.I.; Bast, R.C., Jr.; Berchuck, A.; Berek, J.S.; Brenton, J.D.; Coukos, G.; Crum, C.C.; Drapkin, R.; Etemadmoghadam, D.; et al. Rethinking ovarian cancer: Recommendations for improving outcomes. Nat. Rev. Cancer 2011, 11, 719–725. [Google Scholar] [CrossRef] [Green Version]
- Bowtell, D.D.; Böhm, S.; Ahmed, A.A.; Aspuria, P.J.; Bast, R.C., Jr.; Beral, V.; Berek, J.S.; Birrer, M.J.; Blagden, S.; Bookman, M.A.; et al. Rethinking ovarian cancer II: Reducing mortality from high-grade serous ovarian cancer. Nat. Rev. Cancer 2015, 15, 668–679. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salamini-Montemurri, M.; Lamas-Maceiras, M.; Barreiro-Alonso, A.; Vizoso-Vázquez, Á.; Rodríguez-Belmonte, E.; Quindós-Varela, M.; Cerdán, M.E. The Challenges and Opportunities of LncRNAs in Ovarian Cancer Research and Clinical Use. Cancers 2020, 12, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hull, R.; Mbele, M.; Makhafola, T.; Hicks, C.; Wang, S.M.; Reis, R.M.; Mehrotra, R.; Mkhize-Kwitshana, Z.; Kibiki, G.; Bates, D.O.; et al. Cervical cancer in low and middle-income countries. Oncol. Lett. 2020, 20, 2058–2074. [Google Scholar] [CrossRef]
- Frazer, I.H. Prevention of cervical cancer through papillomavirus vaccination. Nat. Rev. Immunol. 2004, 4, 46–55. [Google Scholar] [CrossRef]
- Kokka, F.; Bryant, A.; Brockbank, E.; Jeyarajah, A. Surgical treatment of stage IA2 cervical cancer. Cochrane Database Syst. Rev. 2014, 2014, CD010870. [Google Scholar] [CrossRef] [PubMed]
- Peirson, L.; Fitzpatrick-Lewis, D.; Ciliska, D.; Warren, R. Screening for cervical cancer: A systematic review and meta-analysis. Syst. Rev. 2013, 2, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Jia, J.; Wang, X.; Liu, Y.; Wang, C.; Fan, R. Long non-coding RNA HOTAIR promotes cervical cancer progression through regulating BCL2 via targeting miR-143–3p. Cancer Biol. Ther. 2018, 19, 391–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Park, S.E.; Lee, C.; Lee, S.Y.; Jo, J.H.; Kim, J.M.; Oh, Y.K. Alterations in promoter usage and expression levels of insulin-like growth factor-II and H19 genes in cervical carcinoma exhibiting biallelic expression of IGF-II. Biochim. Biophys. Acta 2002, 1586, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Liu, L.; Li, G.; Cai, M.; Tan, C.; Han, X.; Han, L. LncRNA GAS5 confers the radio sensitivity of cervical cancer cells via regulating miR-106b/IER3 axis. Int. J. Biol. Macromol. 2019, 126, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Spizzo, R.; Atlasi, Y.; Nicoloso, M.; Shimizu, M.; Redis, R.S.; Nishida, N.; Gafà, R.; Song, J.; Guo, Z.; et al. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013, 23, 1446–1461. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Sun, G.; Zhang, H.; Tian, J.; Li, Y. Long non-coding RNA ANRIL indicates a poor prognosis of cervical cancer and promotes carcinogenesis via PI3K/Akt pathways. Biomed. Pharm. 2017, 85, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Wang, H.L.; Yang, J. Low expression of long non-coding RNA LET inhibits carcinogenesis of cervical cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 806–811. [Google Scholar] [PubMed]
- Yang, M.; Zhai, X.; Xia, B.; Wang, Y.; Lou, G. Long noncoding RNA CCHE1 promotes cervical cancer cell proliferation via upregulating PCNA. Tumour Biol. 2015, 36, 7615–7722. [Google Scholar] [CrossRef] [PubMed]
- Di Cristofano, A.; Ellenson, L.H. Endometrial carcinoma. Annu. Rev. Pathol. 2007, 2, 57–85. [Google Scholar] [CrossRef]
- WHO Classification of Tumors Editoral Board. Female Genital Tumors. Lyon (France) IARC 2020 (WHO Classification of Tumors Series, 5th ed, vol.4). Available online: https://publications.iarc.fr/592 (accessed on 20 December 2020).
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [Green Version]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Pichler, M.; Calin, G.A. MicroRNAs in cancer: From developmental genes in worms to their clinical application in patients. Br. J. Cancer 2015, 113, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Saso, S.; Chatterjee, J.; Georgiou, E.; Ditri, A.M.; Smith, J.R.; Ghaem-Maghami, S. Endometrial cancer. BMJ 2011, 343, d3954. [Google Scholar] [CrossRef] [Green Version]
- Jordan, V.C. The role of tamoxifen in the treatment and prevention of breast cancer. Curr. Probl. Cancer 1992, 16, 129–176. [Google Scholar]
- Murali, R.; Soslow, R.A.; Weigelt, B. Classification of endometrial carcinoma: More than two types. Lancet Oncol. 2014, 15, e268–e278. [Google Scholar] [CrossRef]
- Morice, P.; Leary, A.; Creutzberg, C.; Abu-Rustum, N.; Darai, E. Endometrial cancer. Lancet 2016, 387, 1094–1108. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Y.; Trovik, J.; Sun, K.; Zhou, L.; Jiang, P.; Lau, T.S.; Hoivik, E.A.; Salvesen, H.B.; Sun, H.; et al. A novel wnt regulatory axis in endometrioid endometrial cancer. Cancer Res. 2014, 74, 5103–5117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazal, S.; McKinnon, B.; Zhou, J.; Mueller, M.; Men, Y.; Yang, L.; Mueller, M.; Flannery, C.; Huang, Y.; Taylor, H.S. H19 lncRNA alters stromal cell growth via IGF signaling in the endometrium of women with endometriosis. EMBO Mol. Med. 2015, 7, 996–1003. [Google Scholar] [CrossRef]
- Liu, H.; Wan, J.; Chu, J. Long non-coding RNAs and endometrial cancer. Biomed. Pharm. 2019, 119, 109396. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xing, X.; Li, D.; Zhang, B.; Mutch, D.G.; Hagemann, I.S.; Wang, T. Whole-Genome DNA Methylation Profiling Identifies Epigenetic Signatures of Uterine Carcinosarcoma. Neoplasia 2017, 19, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Cherniack, A.D.; Shen, H.; Walter, V.; Stewart, C.; Murray, B.A.; Bowlby, R.; Hu, X.; Ling, S.; Soslow, R.A.; Broaddus, R.R.; et al. Integrated Molecular Characterization of Uterine Carcinosarcoma. Cancer Cell 2017, 31, 411–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantrell, L.A.; Blank, S.V.; Duska, L.R. Uterine carcinosarcoma: A review of the literature. Gynecol. Oncol. 2015, 137, 581–588. [Google Scholar] [CrossRef]
- Miranda Furtado, C.L.; Dos Santos Luciano, M.C.; Silva Santos, R.D.; Furtado, G.P.; Moraes, M.O.; Pessoa, C. Epidrugs: Targeting epigenetic marks in cancer treatment. Epigenetics 2019, 14, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Han, P.J. Targeting epigenetics for cancer therapy. Arch. Pharm. Res. 2019, 42, 159–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, R.L.; Licht, J.D. Targeting epigenetics in cancer. Annu. Rev. Pharm. Toxicol. 2018, 58, 187–207. [Google Scholar] [CrossRef]
- Nowacka-Zawisza, M.; Wiśnik, E. DNA methylation and histone modifications as epigenetic regulation in prostate cancer (Review). Oncol. Rep. 2017, 38, 2587–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Paredes, M.; Esteller, M. Cancer epigenetics reaches mainstream oncology. Nat. Med. 2011, 17, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Salarinia, R.; Sahebkar, A.; Peyvandi, M.; Mirzaei, H.R.; Jaafari, M.R.; Riahi, M.M.; Ebrahimnejad, H.; Nahand, J.S.; Hadjati, J.; Asrami, M.O.; et al. Epi-drugs and Epi-miRs: Moving beyond current cancer therapies. Curr. Cancer Drug Targets 2016, 16, 773–788. [Google Scholar] [CrossRef]
- Jones, P.A.; Issa, J.P.J.; Baylin, S. Targeting the cancer epigenome for therapy. Nat. Rev. Genet. 2016, 17, 630–641. [Google Scholar] [CrossRef]
- Kazanets, A.; Shorstova, T.; Hilmi, K.; Marques, M.; Witcher, M. Epigenetic silencing of tumor suppressor genes: Paradigms, puzzles, and potential. Biochim. Biophys. Acta 2016, 1865, 275–288. [Google Scholar]
- Vrba, L.; Futscher, B.W. Epigenetic Silencing of MORT Is an Early Event in Cancer and Is Associated with Luminal, Receptor Positive Breast Tumor Subtypes. J. Breast Cancer 2017, 20, 198–202. [Google Scholar] [CrossRef]
- Peng, S.; Yin, X.; Zhang, Y.; Mi, W.; Li, T.; Yu, Y.; Jiang, J.; Liu, Q.; Fu, Y. Competing endogenous RNA network analysis reveals potential long non-coding RNAs as predictive biomarkers of gastric cancer. Oncol. Lett. 2020, 19, 2185–2196. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, B.; Zhang, M.; Guo, W.; Wu, Z.; Wang, Y.; Jia, L.; Li, S.; Cancer Genome Atlas Research Network. lncRNA Epigenetic Landscape Analysis Identifies EPIC1 as an Oncogenic lncRNA that Interacts with MYC and Promotes Cell-Cycle Progression in Cancer. Cancer Cell 2018, 33, 706–720.e9. [Google Scholar] [CrossRef] [Green Version]
- Belete, T.M. The Current Status of Gene Therapy for the Treatment of Cancer. Biologics 2021, 15, 67–77. [Google Scholar]
- Li, T.; Kang, G.; Wang, T.; Huang, H. Tumor angiogenesis and anti-angiogenic gene therapy for cancer. Oncol. Lett. 2018, 16, 687–702. [Google Scholar] [CrossRef] [Green Version]
- Ramarao-Milne, P.; Kondrashova, O.; Barry, S.; Hooper, J.D.; Lee, J.S.; Waddell, N. Histone Modifying Enzymes in Gynaecological Cancers. Cancers 2021, 13, 816. [Google Scholar] [CrossRef]
- Montalvo-Casimiro, M.; González-Barrios, R.; Meraz-Rodriguez, M.A.; Juárez-González, V.T.; Arriaga-Canon, C.; Herrera, L.A. Epidrug Repurposing: Discovering New Faces of Old Acquaintances in Cancer Therapy. Front. Oncol. 2020, 10, 605386. [Google Scholar] [CrossRef]
- Yamada, Y.; Yamada, Y. The causal relationship between epigenetic abnormality and cancer development: In vivo reprogramming and its future application. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2018, 94, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Sogabe, Y.; Seno, H.; Yamamoto, T.; Yamada, Y. Unveiling epigenetic regulation in cancer, aging, and rejuvenation with in vivo reprogramming technology. Cancer Sci. 2018, 109, 2641–2650. [Google Scholar] [CrossRef]
- Bates, S.E. Epigenetic Therapies for Cancer. N. Engl. J. Med. 2020, 383, 650–663. [Google Scholar] [CrossRef]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef]
- Mund, C.; Lyko, F. Epigenetic cancer therapy: Proof of concept and remaining challenges. BioEssays 2010, 32, 949–957. [Google Scholar] [CrossRef]
- Amato, R.J. Inhibition of DNA methylation by antisense oligonucleotide MG98 as cancer therapy. Clin. Genitourin Cancer 2007, 5, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Finsterer, J.; Zarrouk Mahjoub, S. Mitochondrial toxicity of antiepileptic drugs and their tolerability in mitochondrial disorders. Expert Opin. Drug Metab. Toxicol. 2012, 8, 71–79. [Google Scholar] [CrossRef]
- Chen, H.; Dzitoyeva, S.; Manev, H. Effect of valproic acid on mitochondrial epigenetics. Eur. J. Pharm. 2012, 690, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Chan, Y.T.; Tan, H.Y.; Li, S.; Wang, N.; Feng, Y. Epigenetic regulation in human cancer: The potential role of epi-drug in cancer therapy. Mol. Cancer 2020, 19, 79. [Google Scholar] [CrossRef]
- Gould, D.J.; Favorov, P. Vectors for the treatment of autoimmune disease. Gene Ther. 2003, 10, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, L.; Zhang, C. Long noncoding RNAs and tumorigenesis: Genetic associations, molecular mechanisms, and therapeutic strategies. Tumour Biol. 2016, 37, 163–175. [Google Scholar] [CrossRef]
- Áyen, Á.; Jiménez Martínez, Y.; Boulaiz, H. Targeted Gene Delivery Therapies for Cervical Cancer. Cancers 2020, 12, 1301. [Google Scholar] [CrossRef]
- Mizrahi, A.; Czerniak, A.; Levy, T.; Amiur, S.; Gallula, J.; Matouk, I.; Abu-lail, R.; Sorin, V.; Birman, T.; de Groot, N.; et al. Development of targeted therapy for ovarian cancer mediated by a plasmid expressing diphtheria toxin under the control of H19 regulatory sequences. J. Transl. Med. 2009, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Ayesh, S.; Matouk, I.; Schneider, T.; Ohana, P.; Laster, M.; Al-Sharef, W.; De-Groot, N.; Hochberg, A. Possible physiological role of H19 RNA. Mol. Carcinog. 2002, 35, 63–74. [Google Scholar] [CrossRef]
- Matouk, I.J.; DeGroot, N.; Mezan, S.; Ayesh, S.; Abu-lail, R.; Hochberg, A.; Galun, E. The H19 non-coding RNA is essential for human tumor growth. PLoS ONE 2007, 2, e845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, R.; Akhade, V.S.; Pal, D.; Rao, S.M. Long noncoding RNAs in development and cancer: Potential biomarkers and therapeutic targets. Mol. Cell. Ther. 2015, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.M.; Moura, S.R.; Teixeira, J.H.; Barbosa, M.A.; Santos, S.G.; Almeida, M.I. Long noncoding RNAs: A missing link in osteoporosis. Bone Res. 2019, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, Y.; Huarte, M. Long non-coding RNAs: Challenges for diagnosis and therapies. Nucleic Acid Ther. 2013, 23, 15–20. [Google Scholar] [CrossRef] [PubMed]



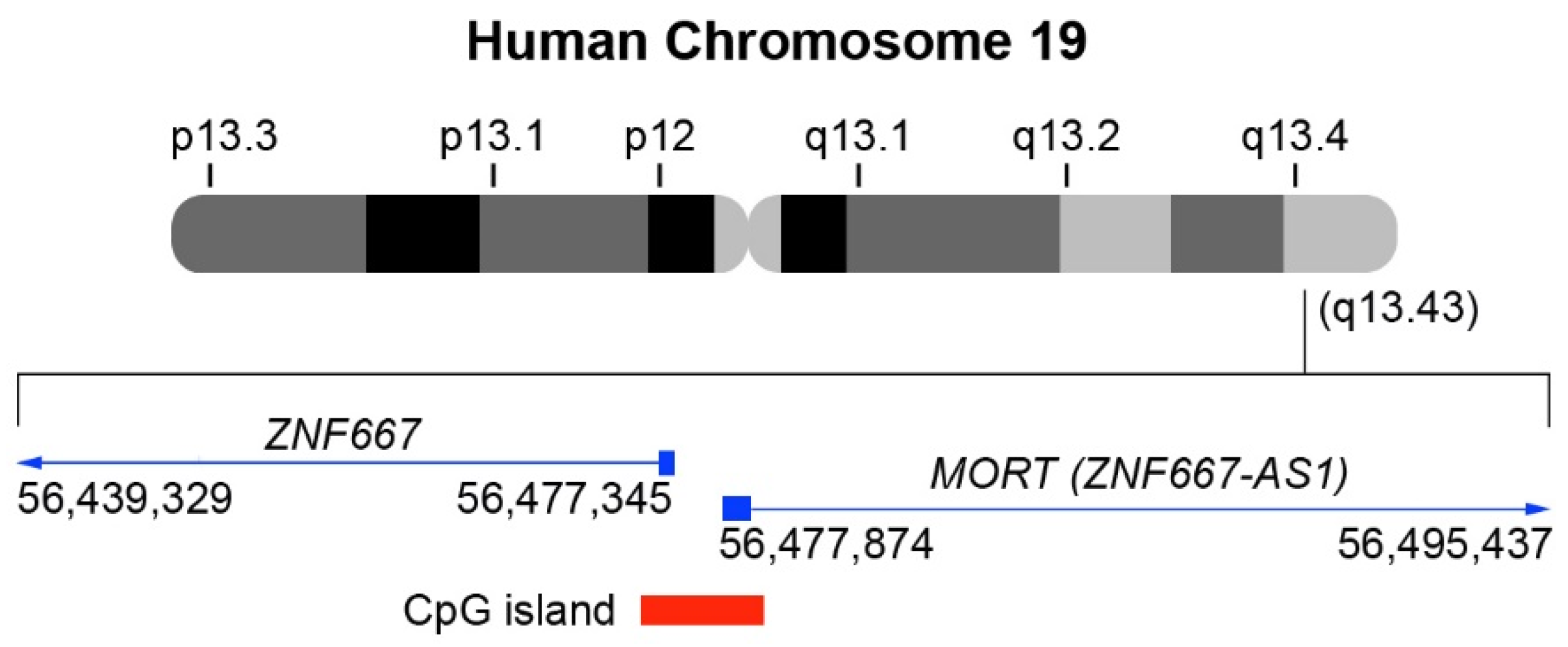{kind=link}
{kind=link}
| Cancer Type | Role | Mechanism | Effect | Reference |
|---|---|---|---|---|
| LSCC | Tumor suppressor | Upregulates ZNF667 expression | Reduces proliferation, migration, and invasion | [26] |
| OSCC | Tumor suppressor | Downregulates ROCK1 expression | Reduces proliferation | [27] |
| NPC | Tumor suppressor | Sponge for miR-1290 | Reduces proliferation both in vitro and in vivo | [28] |
| ESCC | Tumor suppressor | Upregulates ZNF667 expression | Reduces viability, migration, and invasion | [29] |
| UM | Tumor suppressor | Upregulates MEGF10 expression | Inhibits cell proliferation as well as induces apoptosis and cell cycle arrest | [30] |
| LUAD | Tumor suppressor | Downregulates miR-223 | Inhibits cancer cell invasion and migration | [31] |
| CRC | Tumor suppressor | Regulates ANK2/JAK2 expression; Regulates TGF-β1 expression | Inhibits proliferation, invasion and migration; Reduces migration and invasion | [32,33] |
| Bladder | Tumor suppressor | Downregulates miR-146a-5p | Inhibits proliferation, invasion and migration | [34] |
| HCC | Tumor suppressor | Downregulates NOTCH1 expression | Inhibits the migration and invasion | [35] |
| AML | Oncogene | Sponge for miR-206 | Enhances cell proliferation, invasion, tumorigenesis, and metastasis | [36] |
| Glioma | Oncogene | Unknown | Enhances cell proliferation | [37] |
| Cancer Type | Up-/Downregulated or Silenced | Related mRNA/miRNA | Effects | Sources | Clinico- Pathological and Prognostic Significance | Ref. |
|---|---|---|---|---|---|---|
| Ovarian cancer | Down | miR-21 | MORT (ZNF667-AS1) inhibits cell proliferation by miR-21 inhibition | 72 tumor tissues and adjacent healthy tissues; UWB1.289 and UWB1.289+BRCA cells | MORT (ZNF667-AS1) expression is affected by tumor size, but not by tumor metastasis | [39] |
| Cervical cancer | Down | miR-93-3p, PEG3 | MORT (ZNF667-AS1) suppresses proliferation and metastasis via the modulation of miR-93-3p-dependent PEG3 | GEO datasets; 64 cancer tissues and adjacent normal tissues; HeLa and C-33A cells | Unknown | [40] |
| Down | Unknown | MORT (ZNF667-AS1) inhibits cell proliferation | GEO and TCGA datasets; 60 cancer tissues and 30 normal tissues; HeLa and SiHa cells, and HcerEpic (used as control cell line) | Low levels of MORT (ZNF667-AS1) are correlated with decreased overall survival as well as increased tumor size | [41] | |
| Down | Unknown | Unknown | GSE6791 and TCGA datasets | MORT (ZNF667-AS1) could stratify CC patients into the low- and high-risk groups | [42] | |
| Endometrial cancer (EC) | Silenced by aberrant DNA methylation | Unknown | Unknown | TCGA datasets | Unknown | [20] |
| Uterine carcinosarcoma (UCS) | Silenced by aberrant DNA methylation | Unknown | Unknown | TCGA datasets | Unknown | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Fiore, R.; Suleiman, S.; Drago-Ferrante, R.; Felix, A.; O’Toole, S.A.; O’Leary, J.J.; Ward, M.P.; Beirne, J.; Yordanov, A.; Vasileva-Slaveva, M.; et al. LncRNA MORT (ZNF667-AS1) in Cancer—Is There a Possible Role in Gynecological Malignancies? Int. J. Mol. Sci. 2021, 22, 7829. https://doi.org/10.3390/ijms22157829
Di Fiore R, Suleiman S, Drago-Ferrante R, Felix A, O’Toole SA, O’Leary JJ, Ward MP, Beirne J, Yordanov A, Vasileva-Slaveva M, et al. LncRNA MORT (ZNF667-AS1) in Cancer—Is There a Possible Role in Gynecological Malignancies? International Journal of Molecular Sciences. 2021; 22(15):7829. https://doi.org/10.3390/ijms22157829
Chicago/Turabian StyleDi Fiore, Riccardo, Sherif Suleiman, Rosa Drago-Ferrante, Ana Felix, Sharon A. O’Toole, John J. O’Leary, Mark P. Ward, James Beirne, Angel Yordanov, Mariela Vasileva-Slaveva, and et al. 2021. "LncRNA MORT (ZNF667-AS1) in Cancer—Is There a Possible Role in Gynecological Malignancies?" International Journal of Molecular Sciences 22, no. 15: 7829. https://doi.org/10.3390/ijms22157829