1. Introduction
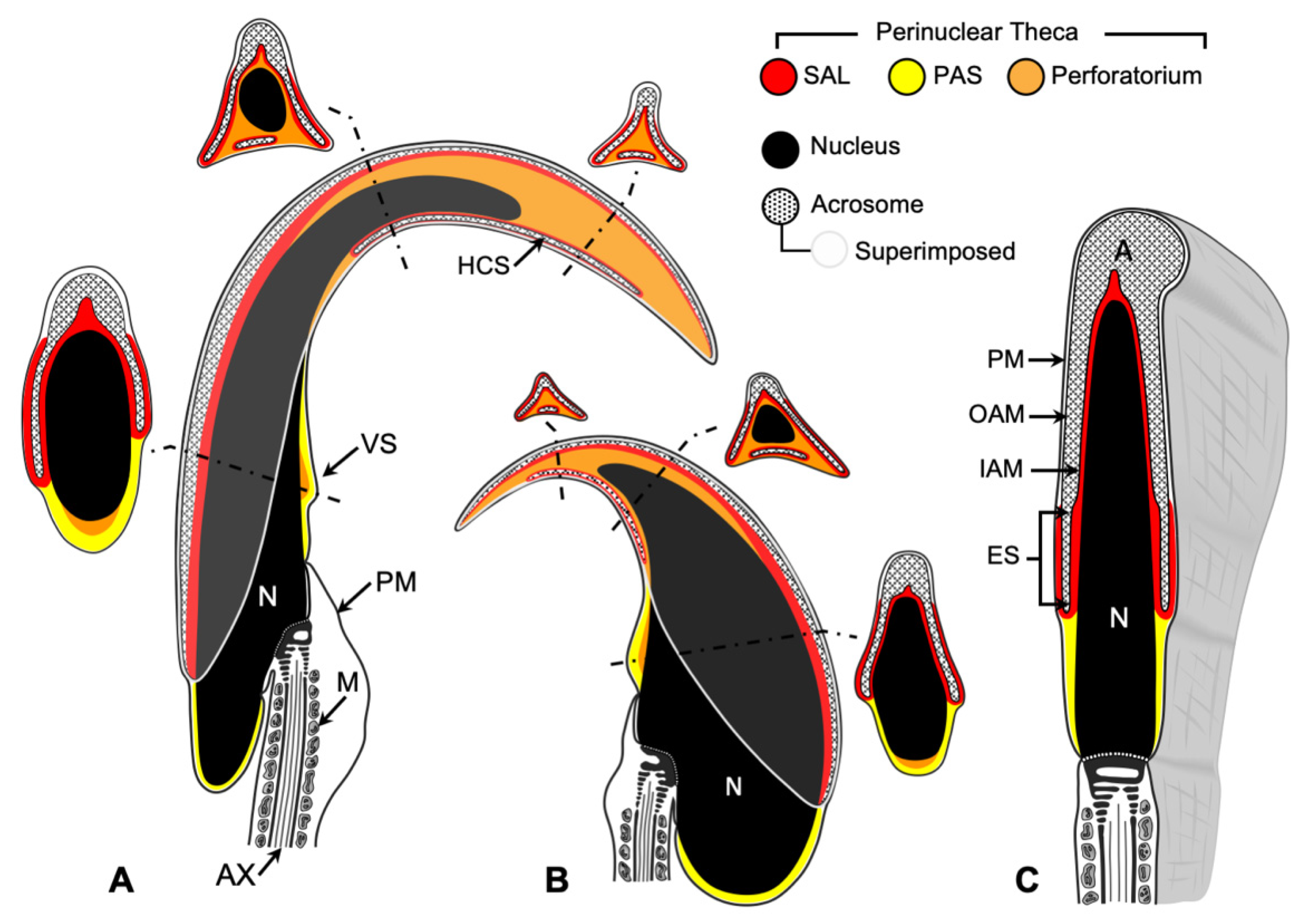
There are three main compartments within the sperm head: the nucleus, the acrosome, and the perinuclear theca (PT) (
Figure 1). The acrosome is a secretory vesicle that encapsulates the anterior region of the head and contains proteins required for oocyte penetration. Underneath the acrosome and surrounding the nucleus is the PT, a condensed cytosolic-protein-rich layer [
1]. Based on compositional and functional considerations, the PT can be divided into two compartments: the subacrosomal layer (SAL), which houses proteins involved in the assembly and attachment of the acrosome to the spermatid nucleus during spermiogenesis [
1,
2,
3,
4,
5,
6], and the post-acrosomal sheath (PAS), which houses proteins implicated in oocyte activation and zygote development during and after fertilization [
1,
7,
8,
9,
10,
11,
12,
13,
14]. In murids, the PT has an additional third compartment termed the perforatorium [
15] (
Figure 1). The perforatorium is a hook-shaped extension of the PT unique to falciform spermatozoa that encircles and extends beyond the apical nucleus. It is compositionally distinct from the SAL but has developmental, compositional, and functional similarities with the PAS [
15].
The fraction of histones retained in the mature eutherian spermatozoon nucleus is species-dependent. The amount of histones found in the spermatid nucleus to date is also highly dependent on isolation and extraction techniques; however, in contrast to the differences in histone retention, there is minimal variability in the final stages of DNA packaging and compaction in eutherian species. Research to date has established a general model accounting for changes in nucleoprotein content during spermatogenesis, which in murids entails the sequential replacement of somatic-type histones by testis-specific variants, prior to the haploid phase, followed by smaller basic proteins commensurate with the increasingly compacted spermatid nucleus. It has been established in murids that prior to and during meiosis, somatic histones H1, H2B, H2A, and H3 are largely replaced by testis-specific histones [
18]. More recently it has been shown that the gene encoding testis-specific histone variant H3t, which has a human counterpart, is essential for entry into spermatogenesis; its loss of function leads to azoospermia and infertility [
19]. A testis-specific counterpart to core histone H4 has yet to be discovered with any certainty. Regardless, the core histones that reside in the spermatid nucleus post-meiotically are replaced by transition proteins in condensing spermatid nuclei [
20,
21,
22], which are in turn replaced by protamines in late spermatids [
23,
24]. The consensus reached in most eutherian mammals is that the majority of basic proteins residing within the condensed mature sperm nucleus are protamines; however, a small amount of histones are retained in the mature sperm nucleus and have been shown to be resistant to ionic detergents used to isolate sperm nuclei [
25,
26,
27,
28] and extract perinuclear histones [
29].
We have previously shown unprecedented non-nuclear localization of core histones in the PAS of bovine spermatozoa [
29]. These histones were extractable from whole bull sperm or their isolated heads with ionic detergents or high-salt solutions; therefore, our first objective was to establish that this distinct core histone localization was not a unique feature of bovine spermatozoa, but rather a common feature of eutherian sperm. We chose the murid model to explore this possibility for several reasons. First, it has been the most commonly used model in studying the changes in nucleoprotein content during spermatogenesis. Secondly, velocity sedimentation techniques developed in murids to separate individual populations of seminiferous epithelial cells allowed us to address whether a new wave of histone synthesis occurs during spermiogenesis that provides core histones for PT assembly. Based on their localization and assembly during spermiogenesis [
30], we hypothesized that the PT histones are synthesized
de novo rather than recycled from the haploid nucleus after histone-replacement in condensing or elongating spermatids. This hypothesis is supported by the observation that hyperacetylation of histones immediately precedes and overlaps the process of nuclear histone replacement by transition proteins, and these acetylated histones are no longer observed in condensed spermatids, even after blocking deacetylase activity [
31]. Thirdly, mouse intracytoplasmic sperm injection (ICSI) is an ideal technique to assess the consequences of inhibiting or depleting PT proteins on early zygotic development [
13] because only the sperm nucleus and PT are required to achieve fertilization in the mouse via ICSI [
8]; thus, we first explored the possibility of selectively extracting histones from the PT of the murid sperm head without disrupting oocyte activation. Once we established that this was possible, we assessed the consequences of PT histone depletion on early embryonic development in ICSI-fertilized mouse oocytes.
3. Discussion
We report unequivocal evidence for the non-nuclear localization of core histones and some of their variants in mature murid spermatozoa. As was found in bull spermatozoa, these non-nuclear histones assemble as part of the postacrosomal sheath (PAS) of the perinuclear theca (PT) in murids but additionally assemble as constituents of the perforatorium (PERF). As recently investigated, the perforatorium is an apical extension of the PT, found in falciform spermatozoa of murids, which shares developmental and compositional similarities with the PAS [
15]. It should be emphasized that the histones assembled in the perinuclear theca make up the majority of the histone content in both the murid and bovine sperm heads to the extent that after high-salt extraction of the PT histones, little to no antigenic traces of histones can be found in the extracted sperm heads, even though their nuclei remain intact; therefore, investigators exploring solely nuclear histones in spermatozoa should take precaution to extract the PT histones, either with an ionic detergent or high salt buffer, from the sperm head before attempting compositional analysis and histone immunolabeling studies of the sperm nucleus. Our findings suggest that the non-nuclear histones found within the perinuclear theca are conserved constituents within eutherian sperm.
Our mass spectrometric (MS) analysis of PT-derived histones suggested that predominately core histones populate the PT of bovine and murid spermatozoa (see
Table 1). This compositional analysis was supported by comparative 2D acid–urea–Triton-SDS-PAGE coupled immunoblotting analysis of bull PT and calf thymus histones performed previously [
29]. It was almost certain from our MS analysis in both murid and bovine that PT constituents H4 and H3 are core histones that are not made up of testis-specific variants. This is not surprising as a testis-specific counterpart to core histone H4 has yet to be discovered [
40,
41] and murid testis-specific H3 (H3t), which has a human counterpart (TH3), is absent in mature mouse spermatozoa, while core H3s (H3.1–3), including somatic variant H3.3, are present [
19]. This could suggest that nucleosomal H3t, which is essential for spermatogonial differentiation and entry into meiosis [
19], is replaced by somatic H3 (H3.1-3) during the round haploid phase of spermiogenesis at the same time that H3 (H3.1-3) synthesis is directed for PT assembly for spermatid elongation. H3.3, which differs from H3.1 by 5 amino acids, can be incorporated into the nucleosome both coupled to and independent of replication and provides a mechanism for the immediate activation of gene transcription [
42,
43].
Analysis of the post-translational modifications (PTMs) on PT-derived histones from both mouse and rat spermatozoa did not show widespread acetylation, indicative of the hyperacetylated state of histones prior to nuclear expulsion, but rather higher rates of methylation (
Table 2). Additionally, as previously stated the H4K16ac modification, a critical signal and target for degradation through a polyubiquitination-independent pathway added in the nucleus during the histone-protamine transition was absent in both our mouse and rat PT-derived histone populations. Most PTMs discovered in the PT-derived histones of both mice and rats had previously been reported in spermatozoa and were present on residues implicated in transcription regulation, such as K18, K23, and K27 of histone H3 (
Table 2) [
33,
34,
35]; however, one PTM of interest, H3R26me2, found in mice has been reported to be an asymmetrical epigenetic modification between maternal and paternal genomes within the early embryo [
44] and has been speculated as a possible marker for pre-patterning in the pre-implantation embryo, as blastomeres with higher H3R26me2 levels show preferential sorting into the inner cell mass [
45]. To date, the origin of the H3R26me2 modification in the paternal genome is unknown but its presence on PT-derived histones may add further support to our hypothesis that the PT histones are not recycled from the spermatid nucleus but rather destined for post-fertilization events.
The relatively high expression of core somatic H2B mRNA found in the isolated rat round spermatids as compared to spermatocytes suggests that its translational product may be destined for PT assembly during the elongation phase of spermiogenesis. This is due to the fact that there appears to be no need for somatic H2B in the maintenance of the spermatid nucleosome, since testis-specific H2B (TH2B) is the predominant H2B-type histone of the round spermatid murid nucleosome, playing a key role in the histone-to-protamine transition of the mouse male genome [
39]; therefore, we hypothesize that the excess production of this histone is transported to and compartmentalized within the PT. This does not exclude TH2B from participating in PT assembly, since immunoblots of PT extracts of murid spermatozoa, utilizing a specific anti-TH2B antibody, as well as mass spectrometry of PT extracts, indicate that TH2B is a constituent of the PT in murids and that a portion of TH2B may even be retained by the nucleus. We are uncertain whether the PT portion of TH2B arises from
de novo synthesis in haploid spermatids or from recycled histones during the nucleosomal–protamine transition; however, there are two lines of evidence that support the former hypothesis. First, it appears that most histones removed from the nuclei of condensing spermatids are discarded and degraded [
31]. Secondly, by injecting, tracking, and comparing intra-testicular
3H-lysine, -arginine, and -thymidine over time in various isolated germ cell populations, Goldberg et al. [
46] demonstrated that histones (H1, H3, H2B, H2A, and H4) are synthesized
de novo in elongating spermatids, becoming an integral part of the spermatozoa. An inherent weakness in this investigation was that there was no accounting for a PT source of histones, as spermatid and spermatozoa “nuclei” were isolated for analysis only in the presence of non-ionic detergents.
Our MS analysis of PT extracts from bovine spermatozoa was unable to detect TH2B. The reason for this could be that as far as we are aware, testis-specific histones have not been found in bovine models. Nevertheless, it appears that somatic H2B and TH2B may be interchangeable in murids; gene ablation of
Th2b in the mouse model was shown to have no phenotypic consequences on spermatogenesis or fertilization and somatic H2B levels were significantly increased in spermatogenic cells in the absence of TH2B, suggesting compensation [
39]. In addition, there is some question as to whether TH2B should even be regarded as testis-specific, as it is also detected in metaphase II mouse oocytes, in paternal and maternal pronuclei, and in polar bodies in one-cell mouse embryos [
39]. In oocytes, the source appears to be the maternal genome, as TH2B is absent in embryos resulting from wild-type males and TH2B-less females.
In the H2A mammalian family, 10 genes encode for conventional or core histone H2A1 types, several genes encode for core histone H2A2 types, and 6 genes encode for histone variants H2AX, H2AZ, macroH2A1, macroH2A2, H2A-Bbd, and H2AV [
43,
47,
48] (UniProt Search,
https://www.uniprot.org (accessed on 7 April 2021)). Although H2A core histone types H2A1 and H2A2 appeared to prevail in the murid and bovine sperm PT, variants H2AV showed up in the PT of both families, while variant H2AX was found in murid sperm alone (
Table 1). H2AV may be essential for heterochromatin formation and repression of transcription [
48], while H2AX, which is 95% similar to conventional H2A, may facilitate the repair of chromosomal double-strand breaks when phosphorylated [
47,
49].
It is doubtful that the energy spent to synthesize and assemble the core histones as part of the PT at the end of spermiogenesis would not lead to a functional purpose. The fact that soon after sperm–oocyte fusion the PAS region of the PT begins to rapidly solubilize in the oocyte cytoplasm and release proteins essential for oocyte activation [
9] and decondensation of the sperm chromatin [
13] suggests that the histones released from the same compartment would also contribute to the fertilization process. Given their inherent ability to bind and protect DNA and regulate its activities, we hypothesized that PT-associated histones, once released in the oocyte cytoplasm, stabilize decondensing sperm chromatin during paternal pronuclear formation in the developing zygote until oocyte or zygote produced histones are able to take over. If this were true, one would expect that zygotes fertilized with PT histone-deficient sperm would experience a delay in early developmental events as the paternal pronucleus attempts to recover from the effects of chromatin instability. In agreement with this idea, developmental delay was observed in zygotes fertilized with PT-histone-depleted spermatozoa as early as the two-cell stage. This delay continued to be prominent into blastocyst stage, suggesting that destabilization of the male chromatin has a prolonged effect on the developing embryo. This interpretation warrants precaution as the possibility exists that the KCl extraction could also remove yet unknown PT-protein entities, in addition to the histones, which could contribute to the embryonic delays observed; however, our lab has not been able to identify any other PT proteins that are KCl-extractable. Furthermore, as far as we are aware, besides WPB2NL/PAWP [
7,
10] and GSTO2 [
13], which are not KCl-extractable, no other sperm PT proteins have yet been shown to be required for fertilization utilizing the ICSI process.
As the embryo transitions through the mitotic cell cycle, checkpoints ensure the completion of phase-specific tasks before proceeding to the next phase. Progression through these cell cycle checkpoints is tightly linked to feedback received from damage-seeking signaling pathways in order to protect DNA from
de novo mutations and erroneous replication [
50]. When errors are detected, the default cellular response is to delay or abort zygotic progression in order to assess the issue, and when possible to repair the damage (reviewed by [
51]). In our study, depletion of core histones from the mouse PT appeared to have triggered this damage control response, resulting in significantly reduced rates of cleavage and blastocyst formation in histone-deficient zygotes relative to ICSI and sonication controls.
Our findings align with previous reports of issues in male chromatin remodeling and early zygotic development resulting from nuclear protein deficiency. In an experiment that induced double-stranded breaks in the male genome via ionizing radiation of sperm cells, zygotes produced by in vitro fertilization experienced a delay in the first round of DNA replication [
52]. Spermatozoa from mice with a transition protein knockout [
53] and spermatozoa from chimeric mice with a protamine deficiency [
54] share common morphological characteristics, such as reduced nuclear condensation states and elevated levels of DNA strand breaks. ICSI with protamine-deficient spermatozoa resulted in normal levels of oocyte activation, but a significant failure to develop to the blastocyst stage. Inadequate amount and localization of protamine 1 was also found to be associated with defects in bull sperm chromatin structure, coinciding with reduced fertility in bulls [
55]. Taken together, these studies provide evidence of altered spermiogenesis and early zygotic development in nuclear-protein-deficient germ cells. Although the mechanism responsible for developmental delay in histone-deficient zygotes remains to be determined, we believe that the bioavailability of PT-derived histones in the ooplasm is inadequate to protect the paternal chromatin as it decondenses. As a result, the net charge of the chromatin, as well as the stoichiometric ratio of its major components, are shifted from being physiologically normal and the chromatin becomes biochemically unstable [
54]. The sperm DNA may, thus, become more susceptible to subsequent degradation in the enzymatically active maternal ooplasm.
In order to determine whether DNA instability contributes to a delay in zygotic progression in PT histone deficient zygotes, future work should consider employing TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labeling) to detect DNA fragmentation in the paternal pronuclei. By fluorescently labeling the 3′-hydroxyl termini in DNA strand breaks, this assay could be a useful tool in assessing the degree of damage sustained by the paternal chromatin following ICSI with histone-deficient spermatozoa. If paternal pronuclei were to display higher levels of TUNEL labeling than maternal pronuclei, this would directly support the hypothesis that PT-derived histones protect the paternal chromatin from damage upon entry into the ooplasm. In addition, 5-ethynyl-29-deoxyuridine (EdU) assays could be employed to determine whether DNA replication is delayed in these histone-deficient zygotes. In a study by Gawecka et al. [
56], ICSI-generated zygotes injected with spermatozoa that had chromatin fragmentation were assessed 8 h post injection using EdU assays and showed impaired DNA synthesizing activity, but were able to recover in the following hours. Confirmation of the function of PT core histones would represent an important development in our current understanding of the mechanisms at play during early embryogenesis. Given the growing issue of male infertility [
57], PT core histones could potentially become valuable biomarkers of sperm quality.
This study has confirmed the presence of histones as prominent constituents of the murid PT, expanding the list of eutherian species (i.e., bovine, porcine, and human) that possess these traditionally nuclear proteins in this non-conventional extra-nuclear sperm head compartment. The localization of primarily core histones to the PT sub-regions that are also assembled from presumed de novo histones translated towards the end of spermiogenesis suggests that the PT histones play a significant role in sexual reproduction. This proposition was supported by our ICSI trials in which mouse oocytes fertilized with PT-histone-deficient sperm frequently failed to reach specific embryonic developmental milestones within the standard accepted timeframe for the species. These results lead us to propose that PT core histones exert nuclear stability during paternal pronucleus formation. In other words, the failure of these histone-deficient zygotes to progress at the same rate as controls may be attributable to destabilization and subsequent degradation of the paternal chromatin following its incorporation into the ooplasm.
4. Materials and Methods
4.1. Animals
Protocols for animal handling and treatment procedures (protocol #1742) were reviewed and approved by the University Animal Care Committee (UACC) of Queen’s University, in accordance with the guidelines of the Canadian Council on Animal Care (CCAC). All mouse procedures were performed using CD-1 (female and male) or C57BL/6 (male) mice, aged 6–12 weeks, purchased from Charles River Laboratories (St. Constant, QC, Canada). Rat spermatozoa were collected from Sprague–Dawley rats, aged 6–12 weeks, purchased from Charles River Laboratories (St. Constant, QC, Canada) or donated by the Department of Biomedical and Molecular Sciences at Queen’s University. Bovine epididymides used for sperm collection were collected from mature bulls immediately after slaughter from local abattoirs.
4.2. Sperm Collection
Spermatozoa were obtained from the fresh cauda epididymides of sacrificed mature rats and mice. The epididymides were placed into phosphate-buffered saline (PBS) or Tris-buffered saline (TBS) solutions (pH 7.4–7.8), depending upon the subsequent methodology. Spermatozoa were collected by cutting and gently squeezing the cauda epididymides so that the spermatozoa diffused into the solution. The sperm-containing suspension was aliquoted into Eppendorf tubes and washed by centrifugation in the buffer of choice at 1000×
g for 5 min to be used for Western blotting or immediately fixed in 2% formaldehyde before being stored at 4 °C in PBS for use in indirect immunofluorescence. Sperm concentration was determined by counting cells using in a hemocytometer. Intact bovine sperm were extracted according to methods previously described by Oko and Maravei [
16] from fresh epididymides or frozen whole epididymides that were thawed.
For testicular smears, spermatogenic cells were obtained from the testes of sacrificed mature rats and mice. The testicular smears were processed for indirect immunofluorescence according to previously published protocols [
58]. Briefly, the tunica albuginea was removed from the testes and the testes were placed in 5ml 200mM PBS. The testes were then cut into smaller pieces using a razor blade and spermatids were allowed to diffuse out of the seminiferous tubules into the solution.
4.3. SDS and High-Salt Extraction of Sperm Histones
For SDS extraction, 2 × 107 murid sperm were incubated in 200 µL 2% sodium dodecyl sulfate (SDS) for 5 min at 100 °C. For high-salt extraction, 2 × 107 murid sperm cells were suspended in 200 µL 1 M potassium chloride (KCl) solution and sonicated on ice for 5 s at 0.1 A. Each solution resulting from one of the above extraction methods was promptly separated into supernatant and pellet fractions by centrifugation at 16,000× g for 4 min. Following removal of the supernatant, the pellet was washed with PBS to remove any residual solubilized proteins or salts. For storage, the proteins in the KCl supernatant were isolated by chloroform–methanol precipitation and dried. The core histone identity of extracted proteins was verified using SDS-PAGE and Western blot analyses, as described below.
4.4. SDS-PAGE and Western Blot Analysis
Immediately following extractions (see above), supernatants and pellets were incubated in reducing sample buffer (200 nM Tris pH 6.8, 4% SDS, 0.1% bromophenol blue, 5% β-mercaptoethanol, 40% glycerol) to preserve their total protein profiles. Samples were separated along a molecular weight (MW) gradient by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), as previously described by Laemmli [
59]. A BLUeye pre-stained protein ladder (GeneDirex, Taiwan) and calf thymus histones (Sigma-Aldrich, St. Louis, MO, USA, H9250) were run in adjacent lanes and served as general and histone-specific MW references, respectively. Gels were stained with Coomassie Brilliant Blue 250 (Sigma-Aldrich, St. Louis, MO, USA) or used for immunoblot analysis. For immunoblots, one sample (i.e., sperm supernatants, pellets, and whole sperm) equivalent to 5–10 million sperm was loaded per lane in each experiment and resolved on 4% stacking and 18% separating polyacrylamide. Electrophoresis-separated proteins were transferred onto methanol-activated polyvinylidene fluoride (PVDF) membranes (Millipore, Mississauga, ON, Canada) according to methods adapted from [
60]. To prevent non-specific labeling, membranes were blocked with 10% skim milk in PBS containing 0.05% Tween-20 (PBS-T). Membranes were then cut at the 25 kDa level for probing with two separate primary antibodies at 4 °C, overnight; the upper PVDF strip (>25 kDa) was incubated in anti-PAWP serum (1:1000 dilution), while the lower strip (<25 kDa) was incubated in anti-pan-histone serum (1:4000 dilution). Membranes were washed generously with PBS-T before and after incubation with an HRP-conjugated secondary antibody (diluted to 1:40,000). The resulting blots were exposed to Clarity™ Western ECL Substrate (Bio-Rad, Mississauga, ON, Canada) and developed on X-ray film (Eastman Kodak Company, Rochester, NY, USA).
4.5. Antibodies
The primary antibody used for Western blotting and immunodetection experiments was a polyclonal rabbit anti-histone made in house against purified bovine calf thymus histones (anti-pan histone antibody, also rereferred to as H5). Antibodies specific to each of the histone subtypes were subsequently made through affinity purification of the original serum using recombinant forms of each histone [
29]. An affinity purified polycolonal rabbit H3 specific antibody (Proteintech, Rosemont, IL, USA, 17168-1-AP) and a testis-specific H2B antibody (TH2B) (EMD Millipore, Burlington, MA, USA, 07-680) were also used at the manufacturer’s suggested concentrations in all applications. The anti-pan histone (H5) antibody was applied at a 1:4000 concentration for Western blot analysis and at a 1:50 concentration for immunofluorescence and immunoperoxidase histochemistry. For Western blot analysis, the secondary antibody used was goat anti-rabbit IgG-HRP (horseradish peroxidase) (VectorLab, Burlingame, CA, USA, 1:50,000), whereas for immunofluorescence, donkey anti-rabbit IgG-CFL488 (Santa Cruz, Dallas, TX, USA sc-362261, 1:200) was used. The secondary antibody used in enzymatic immunohistochemistry was a biotin-labeled anti-rabbit IgG (VectorLab, Burlingame, CA, USA 1:200). A mouse monoclonal anti-alpha tubulin antibody (Sigma-Aldrich, St. Louis, MO, USA, T6074) was used in immunofluorescence co-localization experiments at a concentration of 1:2500 and a donkey-anti mouse IgG-CFL647 (Santa Cruz, Dallas, TX, USA sc-362288 1:200) was used as the secondary antibody.
4.6. Immunofluorescence
Histones of whole or protein-extracted rat and mouse spermatozoa were analyzed by indirect immunofluorescence according to methods described by Sutovsky et al. [
61]. Spermatids or spermatozoa were mounted on poly-L-lysine-coated coverslips in KMT buffer (100 mM KCl, 2 mM MgCl
2, 10 mM Tris-HCl, 5 mM EGTA, pH 7.0). The cells were fixed with 2% formaldehyde in phosphate saline buffer (PBS) for 30 min before being permeabilized in PBS with 0.1% Triton-x-100 (PBS-Tx) for 40 min at room temperature. The cells were blocked with 5% normal goat serum (NGS) in PBS-Tx for 25 min to avoid non-specific binding, then incubated in a humidity chamber with primary antibody diluted in 1% NGS overnight at 4 °C. On the following day, the cells were washed with 1% NGS three times followed by a 40 min incubation with fluorescently tagged secondary antibodies diluted in 1% NGS at room temperature and shielded from light. The secondary antibody mixture included blue fluorescence DNA stain DAPI (4′,6-diamidino-2-phenylindole, dihydrochloride). The coverslips were then mounted onto glass slides using Vectashield mounting media (Vector Laboratories, Burlingame, CA, USA) and sealed with nail polish. Images were captured at the Queen’s University Cancer Biology Institute Imaging Centre using a Quorum Wave Effects spinning disc confocal microscope.
4.7. Immunohistochemistry
Murid testicular sections from testes that had been perfusion-fixed in Bouin’s fixative or paraformaldehyde and embedded in paraffin were deparaffinized in xylene and hydrated through a graded series of ethanol solutions. During hydration, the sections were treated to abolish endogenous peroxidase activity, to neutralize residual picric acid, and to block free aldehyde groups [
22]. Once hydrated, the sections were subjected to antigen retrieval by microwave irradiation in a 5% urea Tris-HCl, pH 9.5 solution. Immunolabeling was conducted using an avidin-biotin complex (ABC) kit (Vector Laboratories, Burlingame, CA, USA) and followed the procedure outlined by Ferrer et al. [
62].
4.8. Electron Microscopic Immunocytochemistry
Adult male Sprague–Dawley rats (350–450 g) were anesthetized and their testes were fixed by perfusion for 10 min through the abdominal aorta with 0.5% glutaraldehyde and 4% paraformaldehyde in 0.1 M phosphate buffer containing 50 mM lysine at pH 7.4. After their removal, the testes were cut into small pieces (0.5 mm3), immersed for 2 h in the above fixative at 4 °C, washed 2–3 times in PBS containing 4% sucrose (pH 7.4), and treated with PBS containing 4% sucrose and 50 mM NH4Cl for 1 h at 4 °C. The tissue pieces were washed, dehydrated in a graded series of methanol up to 90%, and embedded in LR white (Canemco, St Laurens, QC, Canada). They were then polymerized at −20 °C, cut into ultrathin sections, and mounted on Formavar coated nickel grids. During immunolabeling, grids were floated tissue-side down on drops of various solutions. Sections were blocked with 5% NGS in TBS, then incubated with affinity purified anti-histone antibodies overnight at 4 °C. They were subsequently washed in TWBS (TBS with 0.1% Tween-20, pH 8) extensively, blocked with 5% NGS for 15 min, incubated in 10 nm gold-particle-conjugated goat anti-rabbit IgG (Sigma, Mississauga, ON, Canada) for 2 h at 21 °C, washed with TWBS and deionized water, and allowed to dry. The sections were then counterstained with uranyl acetate and lead citrate, washed with deionized water, and dried. Photographs were taken using a Hitachi 7000 transmission electron microscope. The results shown are typical of three different animals and experiments.
4.9. LC-MS/MS and Data-Dependent Aquisition (DDA)
Precipitated proteins of the sperm KCl extracts (see Methods above) were resuspended in rehydration buffer (8 M Urea, 2% sodium deoxycholate, 10 mM Tris, pH 7.5). To the suspension, 10 mM DTT was added for 1h, followed by 20 mM iodoacetamide for another hour in the dark. In order to precipitate the sample, one volume of chloroform and two volumes of methanol were added to two volumes of sample and the mixture was vortexed vigorously. This sample was centrifuged (10,000× g, for 2 min), then after phase separation, the upper phase was removed. Three volumes of methanol were added to the remaining lower phase and the mixture was gently inverted twice. The solution was then centrifuged (10,000× g, 15 min) to pellet the precipitated protein. The supernatant was discarded and the pellet was allowed to air dry. Trypsin (1:50, w/w) was added to the pellet and digested overnight at 37 °C. Samples were centrifuged (17,000× g for 20 min), transferred to sample vials, and put under vacuum concentration until dry.
NanoLC-MS/MS was performed using a Dionex UltiMate 3000 nanoLC system (Dionex). Peptides from the trypsin-digested samples of the histone-enriched PT fraction were suspended in buffer A (2% ACN/0.1% TFA) and directly loaded onto a 50 cm analytical column packed with Acclaim PepMap C18 2 μm sorbent. Peptides were eluted using a 110 min gradient from 7 to 40% buffer B (95% ACN, 0.1% TFA) at 250 nL min−1 and nanoelectrosprayed into a Q-Exactive Plus (Thermo Fisher Scientific, Waltham, MA, USA). Precursor scans of intact peptides were measured in the Orbitrap by scanning from m/z 350 to 1500 (with a resolution of 70,000), then the fifteen most intense multiple charged precursors were selected for HCD fragmentation with a normalized collision energy of 32.0 and measured in the Orbitrap at a resolution of 35,000. Automatic gain control targets were 3E6 ions for Orbitrap scans and 5E5 for MS/MS scans. Dynamic exclusion was employed for 15 s. Fragmentation data were converted to peak lists using Xcalibur version 4.027.19 (Thermo Fisher Scientific) and the HCD data were processed using Proteome Discoverer 2.1 (Thermo Fisher Scientific, Waltham, MA, USA). MS spectra were then searched with Mascot V2.6 (accessed on 5 December 2019) against all mouse and rat entries in SwissProt (Release December 2019, 17,033 entries). Mass tolerances in MS and MS/MS modes were 10 ppm and 0.02 Da, respectively; trypsin was designated as the digestion enzyme and up to two missed cleavages were allowed. Carbamidomethylation of cysteine residues was designated as a fixed modification. The variable modifications included were oxidation of methionine and deamidation of asparagine or glutamine. Interrogation of the corresponding reversed database was also performed to evaluate the false discovery rate (FDR) of peptide identification using Percolator based on q-values, which were estimated from the target–decoy search approach. To filter out target peptide spectrum matches (target-PSMs) over the decoy PSMs, a FDR of <1% was set at the peptide level. An additional identification criterion sought a minimum of two uniquely matched peptides per protein.
4.10. Separation of Testicular Germ Cells from Rat Testes Using Unit Gravity Sedimentation
The enzymatic digestion of testicular cells and their sedimentation on a BSA gradient using a STAPUT apparatus were assessed using an approach adapted from previous publications on this method [
63,
64,
65].
Two Sprague–Dawley rats were asphyxiated with CO2 following the standard operating procedures of the Queen’s Animal Care Committee. Testes were retrieved, decapsulated, and separated into two 50mL tubes containing 30mL of RPMI-1640 medium (Hyclone, Logan, UT, USA, SH30011.03) with 1.5 mg/mL collagenase (Bioshop, Burlington, ON, Canada COL007.500). The tubes were incubated for 30 min at 37 °C with agitation. The separated and digested testicular tubules were collected by centrifugation at 200× g for 5 min at 4 °C, then washed once with RPMI-1640 medium. Once the supernatant was removed, the tubules were cut into pieces, mixed with 30 mL of preheated RPMI-1640 medium containing 1% BSA pH 7.4 (Fisher Bioreagent, Waltham, MA, USA, S-15898) and 1.5 mg/mL trypsin (Gibco, Waltham, MA, USA, (2.5%) #12090), and incubated for 20 min at 37 °C with agitation. Then, 10% FBS (ThermoFisher Scientific, Waltham, MA, USA) was added to the above mixture, supplemented by 15 uL/tube (150U) of DnaseI (Bioshop, Burlington, ON, Canada, DRB001.10) to prevent high viscosity due to genomic DNA. Tubular cells were released by performing gentle pulsing movements with a transfer pipette for 3–5 min. Subsequently, the cells were filtered through a 100 μm cell strainer.
Using a STAPUT sedimentation apparatus set up in a refrigerated room with minimal vibration, the testicular cell suspension was loaded on a 2–4% BSA gradient at pH 7.4. The flow rate was adjusted to form the gradient in 70 min. The cells were then placed on the gradient and allowed to flow through for 90 min. The first five 10 mL fractions that were collected were made up of unwanted debris. After this initial collection, the circuit was closed to allow for another 90 min of cell separation. Fractions of 10 mL were then collected at a rate of 10 mL/30 min and their purity levels for tetraploid cells and round spermatids were assessed and pooled accordingly. For purity analysis, cells were fixed in Carnoy’s fixative(methanol/acetic acid 3:1) and stained with DAPI or periodic acid–Schiff stain. The cellular perimeter and cellular surface of the tetraploid cells and haploid cells were assessed using phase microscopy. Cells in each pool were counted using a hemocytometer and aliquots of 1 × 106 cells/tube were compiled for RNA analysis.
4.11. RNA Isolation and Quantitative PCR (RT-qPCR) Analysis of Somatic H2B mRNA
The RNA was extracted using a combination of TRIZOL (Invitrogen, Waltham, MA, USA, 15596-018) and the Tissue Total RNA Mini Kit (GeneAid, Taiwan, RT050). Once the TRIZOL extraction was completed, following the manufacturer’s recommendations, one volume of 70% ethanol was added to the aqueous phase and loaded on an RB column. Subsequent steps were performed per the manufacturer’s recommendation. Triplicates of RNA extractions were processed for each group, since there were no reliable reporter genes that could have been used to measure the relative expression of H2B in a population of tetraploid cells compared to round spermatids.
RNA samples were digested with DNAseI, then rat testicular and somatic H2B mRNA samples were reversed-transcribed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA, #436884) according to the manufacturer’s instruction. The RT primer used was 5′-GCTGGTGTACTTGGTG-3′, which is specific to both somatic and testicular H2B. Proper controls were prepared to ensure that each sample was genomic-DNA-free. Somatic H2B qPCR primers were designed from Rattus H2B (NCBI; NM_022647.2), making sure that they were not going to amplify the testis-specific variant H2B (GenBank: M18046.1). The forward primer was 5′-ATGCCTGAGCCTGCGAAGT-3′, while the reverse primer was 5′-CATGGCCTTGGAAGAGATGC-3′.
The relative mRNA expression of rat somatic H2B was measured using Roche LightCycler® 480 System II (Roche Scientific, Laval, QC, Canada). The cycling parameters consisted of an initial denaturation step at 95 °C for 10 min, then another 20 s in the same conditions, followed by touchdown annealing temperatures from 70 to 67 °C (decreasing step size of 0.5 °C per cycle) for 15 sec each and 72 °C for 15 s to ensure precise hybridization. The total number of cycles was 43. Finally, melt curve analyses were performed to ensure the quality and specificity of qPCR products.
4.12. Gamete Isolation for Intracytoplasmic Sperm Injection
Oocyte collection: Mature CD-1 female mice aged 8–12 weeks were superovulated with intraperitoneal injections of 10 IU pregnant mare serum gonadotropin (PMSG; Sigma-Aldrich, St. Louis, MO, USA) and 10 IU human chorionic gonadotropin (hCG; Sigma-Aldrich, St. Louis, MO, USA), administered 48 h apart. Mice were euthanized at 12–14 h post-hCG injection by isoflurane inhalation and cervical dislocation and oviducts were removed and placed in Advanced KSOM (Sigma-Aldrich, St. Louis, MO, USA, MR-101-D). Cumulus-oocyte complexes were released and transferred to Advanced KSOM droplets containing 0.1% hyaluronidase (Sigma-Aldrich, St. Louis, MO, USA, H3757) for cumulus cell removal. Cumulus-free oocytes were washed and incubated in Advanced KSOM with mineral oil (Sigma-Aldrich, St. Louis, MO, USA, M8410) at 37 °C and 5% CO2 until use.
Sperm collection: Study animals were euthanized by isoflurane inhalation and cervical dislocation. Mouse cauda epididymides were isolated from adult males, pierced with a needle, and gently squeezed to allow the sperm to diffuse into a droplet of human tubal fluid (HTF) (Millipore, Burlington, MA, USA MR-070-D) insulated by mineral oil (ICSI CTRL) or into PBS for subsequent treatment. For high-salt extraction, sperm was spun down and resuspended in 200 µL 1 M potassium chloride (KCl) solution and sonicated on ice for 5 s at 0.1 A (histone-depleted, HD), whereas the sonicated control (SON CTRL) was sonicated on ice for 5 s at 0.1 A in PBS. The sperm were subsequently incubated at 37 °C and 5% CO2 until required, no more than three hours post-collection. In each ICSI replicate, spermatozoa from one male were used for all treatments to control for variations between animals.
4.13. Intracytoplasmic Sperm Injection (ICSI)
ICSI was carried out with CD-1 mice according to the methods described by Yanagimachi [
66] across 3–4 experimental trials. The procedure was performed using a Nikon Ti-S inverted microscope (Nikon Canada Inc., Mississauga, ON, Canada) fitted with Narishige micromanipulators (Narishige International US Inc., Amityville, NY, USA) and a Piezo PMM150HJ/FU drill (Prime Tech Ltd., Ibaraki, Japan). Briefly, a single spermatozoon was drawn up into the injection pipette from a sperm suspension containing 6–10% polyvinylpyrrolidone (PVP; Sigma-Aldrich, St. Louis, MO, USA, PVP360). Motile sperm were immobilized by applying several piezo pulses to the midpiece, separating heads and tails. Injections were performed in Advanced KSOM droplets; the oocyte was held in place by the holding pipette and piezo pulses were applied to “drill” through the zona pellucida with the injection pipette. Once the zona was punctured, the sperm head was expelled into the ooplasm near the opposite side of the oocyte and the pipette was gently withdrawn. Sperm-injected oocytes were left to rest at RT for 10 min before being transferred to new Advanced KSOM droplets under mineral oil at 37 °C, 5% CO
2. Injected embryos were left to incubate for 6 h, 22–24 h, 4 d, or 5 d, depending on experimental group. Oocytes were analyzed by differential interference contrast microscopy on a Nikon Ti-S inverted microscope (Nikon Canada Inc., Mississauga, ON, Canada) and photographed using a QImaging QICAM Fast 1394 digital camera.
4.14. Statistics
Statistical significance was determined based on two-tailed T-tests performed between ICSI CTRL, SON CTRL, and histone-depleted (HD) experimental groups. The standard error of the mean was calculated to reflect variation within each group. Here, p values < 0.05 were considered significant.


 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}