
Retinal Pigment Epithelium Expressed Toll-like Receptors and Their Potential Role in Age-Related Macular Degeneration



Abstract
:1. Introduction
2. Results
2.1. Toll-Like Receptors in AMD
2.1.1. TLR Polymorphisms and Patient Studies
2.1.2. Pre-Clinical Data
2.2. Toll-Like Receptors in the RPE
2.2.1. TLR Expression in the RPE
2.2.2. TLR Activation in the RPE
2.2.3. TLR-Activated RPE and Microglia/Monocytes
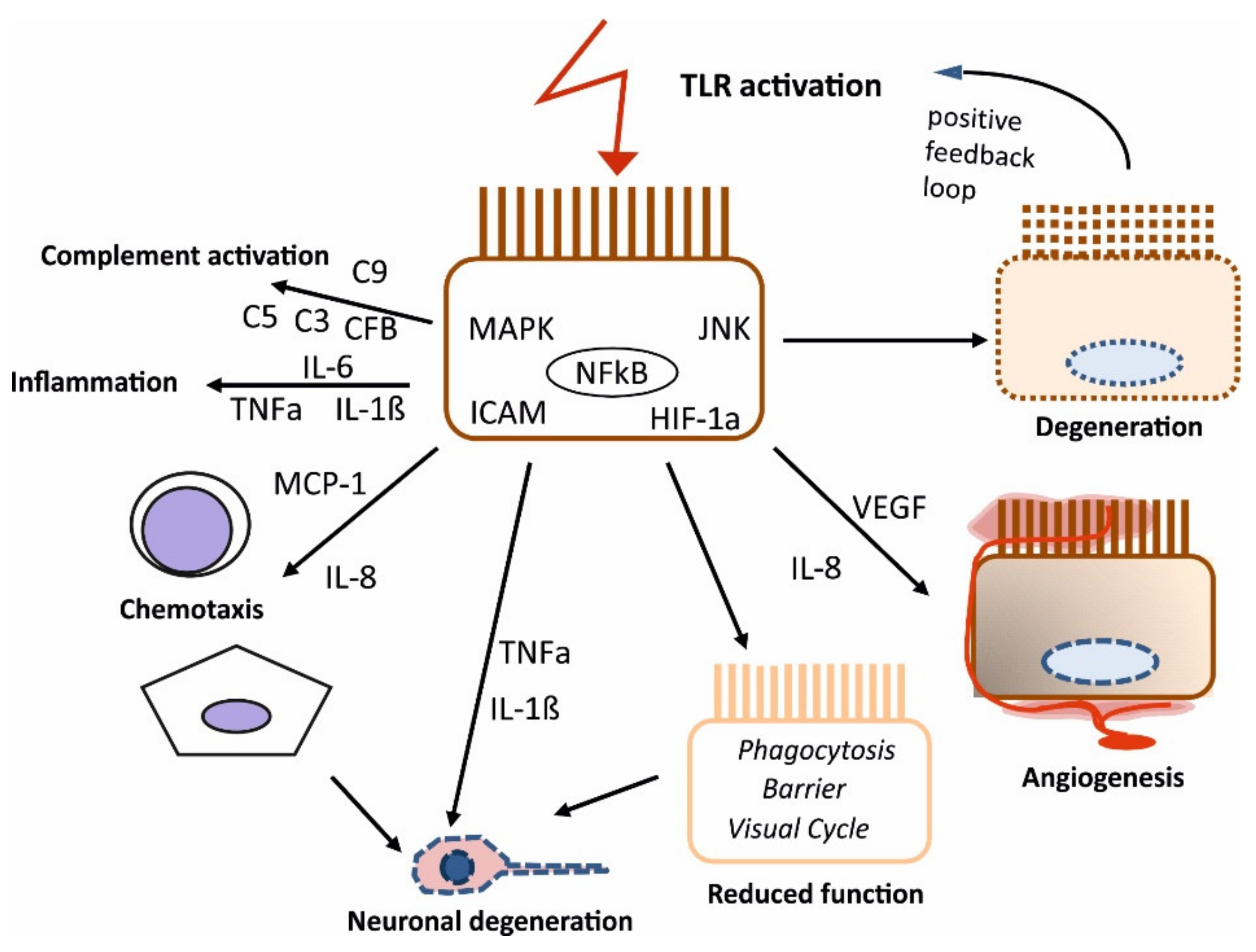
2.3. Potential Role of TLR Activation of the RPE in the Development of AMD
3. Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bourne, R.R.A.; Flaxman, S.R.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; Leasher, J.; Limburg, H.; et al. Magnitude, temporal trends, and projections of the global prevalence of blindness and distance and near vision impairment: A systematic review and meta-analysis. Lancet. Glob. Health 2017, 5, e888–e897. [Google Scholar] [CrossRef] [Green Version]
- Hageman, G.S.; Luthert, P.J.; Chong, N.V.; Johnson, L.V.; Anderson, D.H.; Mullins, R.F. An integrated hypothesis that considers drusen as biomarkers of immune-mediated processes at the RPE-Bruch’s membrane interface in aging and age-related macular degeneration. Prog. Retin. Eye Res. 2001, 20, 705–732. [Google Scholar] [CrossRef]
- Miller, J.W. Beyond VEGF-The Weisenfeld Lecture. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6911–6918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.Y.; Creuzot-Garcher, C.; Darmon, J.; Desmettre, T.; Korobelnik, J.F.; Levrat, F.; Quentel, G.; Paliès, S.; Sanchez, A.; de Gendre, A.S.; et al. Types of choroidal neovascularisation in newly diagnosed exudative age-related macular degeneration. Br. J. Ophthalmol. 2007, 91, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Erfurth, U.; Chong, V.; Loewenstein, A.; Larsen, M.; Souied, E.; Schlingemann, R.; Eldem, B.; Mones, J.; Richard, G.; Bandello, F. Guidelines for the management of neovascular age-related macular degeneration by the European Society of Retina Specialists (EURETINA). Br. J. Ophthalmol. 2014, 98, 1144–1167. [Google Scholar] [CrossRef] [PubMed]
- Versteeg-Tijmes, N.T.; de Jong, P.T.; Bos, P.J.; Bakker, D.; Greve, E.L.; Oosting, J.; Nagelkerke, N.J.; vd Horst-Breetveld, H.M. Argon laser treatment of pigment epithelial detachments and of subretinal neovascular membranes in Junius-Kuhnt’s senile disciform macular degeneration. A prospective, randomized study. Graefe’s Arch. Clin. Exp. Ophthalmol. 1982, 218, 271–274. [Google Scholar] [CrossRef]
- Curcio, C.A.; Johnson, M.; Rudolf, M.; Huang, J.D. The oil spill in ageing Bruch membrane. Br. J. Ophthalmol. 2011, 95, 1638–1645. [Google Scholar] [CrossRef]
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular endothelial growth factor a in intraocular vascular disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef]
- Shaw, P.X.; Stiles, T.; Douglas, C.; Ho, D.; Fan, W.; Du, H.; Xiao, X. Oxidative stress, innate immunity, and age-related macular degeneration. AIMS Mol. Sci. 2016, 3, 196–221. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and its role in age-related macular degeneration. Cell. Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Chen, M.; Forrester, J.V. Para-inflammation in the aging retina. Prog. Retin. Eye Res. 2009, 28, 348–368. [Google Scholar] [CrossRef]
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klettner, A.; Kauppinen, A.; Blasiak, J.; Roider, J.; Salminen, A.; Kaarniranta, K. Cellular and molecular mechanisms of age-related macular degeneration: From impaired autophagy to neovascularization. Int. J. Biochem. Cell Biol. 2013, 45, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.Z. Age-related macular degeneration (AMD): Pathogenesis and therapy. Pharmacol. Rep. 2006, 58, 353–363. [Google Scholar]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef] [Green Version]
- Klettner, A. Oxidative stress induced cellular signaling in RPE cells. Front. Biosci. 2012, 4, 392–411. [Google Scholar] [CrossRef]
- Klettner, A. The Retinal Pigment Epithelium in the Immune Regulation of the Retina. In Recent Advances in Eye Research; Hogarth, A., Ed.; Nova Science Publishers: New York, NY, USA, 2015; pp. 31–52. [Google Scholar]
- Detrick, B.; Hooks, J.J. Immune regulation in the retina. Immunol. Res. 2010, 47, 153–161. [Google Scholar] [CrossRef]
- Jørgensen, A.; Wiencke, A.K.; La Cour, M.; Kaestel, C.G.; Madsen, H.O.; Hamann, S.; Lui, G.M.; Scherfig, E.; Prause, J.U.; Svejgaard, A.; et al. Human retinal pigment epithelial cell-Induced apoptosis in activated T cells. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1590–1599. [Google Scholar]
- Sugita, S.; Usui, Y.; Horie, S.; Futagami, Y.; Aburatani, H.; Okazaki, T.; Honjo, T.; Takeuchi, M.; Mochizuki, M. T-cell suppression by programmed cell death 1 ligand 1 on retinal pigment epithelium during inflammatory conditions. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2862–2870. [Google Scholar] [CrossRef]
- Gregerson, D.S.; Heuss, N.D.; Lew, K.L.; McPherson, S.W.; Ferrington, D.A. Interaction of retinal pigmented epithelial cells and CD4 T cells leads to T-cell anergy. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4654–4663. [Google Scholar] [CrossRef]
- Hooks, J.J.; Nagineni, C.N.; Hooper, L.C.; Hayashi, K.; Detrick, B. IFN-Beta provides immuno-Protection in the retina by inhibiting ICAM-1 and CXCL9 in retinal pigment epithelial cells. J. Immunol. 2008, 180, 3789–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamiri, P.; Masli, S.; Kitaichi, N.; Taylor, A.W.; Streilein, J.W. Thrombospondin plays a vital role in the immune privilege of the eye. Investig. Ophthalmol. Vis. Sci. 2005, 46, 908–919. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, N.; Chen, L.; Tokura, T.; Ushio, H.; Iwatsu, M.; Murakami, A. Distinct functions between toll-like receptors 3 and 9 in retinal pigment epithelial cells. Ophthalmic Res. 2007, 39, 155–163. [Google Scholar] [CrossRef]
- Bernard, J.J.; Cowing-Zitron, C.; Nakatsuji, T.; Muehleisen, B.; Muto, J.; Borkowski, A.W.; Martinez, L.; Greidinger, E.L.; Yu, B.D.; Gallo, R.L. Ultraviolet radiation damages self noncoding RNA and is detected by TLR3. Nat. Med. 2012, 18, 1286–1290. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.C.; Young, D.W.; Golenbock, D.T.; Christ, W.J.; Gusovsky, F. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem. 1999, 274, 10689–10692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Schwandner, R.; Dziarski, R.; Wesche, H.; Rothe, M.; Kirschning, C.J. Peptidoglycan- and lipoteichoic acid-induced cell activation is mediated by toll-like receptor 2. J. Biol. Chem. 1999, 274, 17406–17409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Micera, A.; Stampachiacchiere, B.; Aronni, S.; dos Santos, M.S.; Lambiase, A. Toll-like receptors and the eye. Curr. Opin. Allergy Clin. Immunol. 2005, 5, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.A.; Wei, R.; Branch, M.J.; Sidney, L.E.; Amoaku, W.M. Expression of Toll-like receptors in human retinal and choroidal vascular endothelial cells. Exp. Eye Res. 2015, 138, 114–123. [Google Scholar] [CrossRef]
- Redfern, R.L.; McDermott, A.M. Toll-like receptors in ocular surface disease. Exp. Eye Res. 2010, 90, 679–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Yu, F.S. Toll-like receptors and corneal innate immunity. Curr. Mol. Med. 2006, 6, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, N.; Fukuda, K.; Fujitsu, Y.; Lu, Y.; Chikamoto, N.; Nishida, T. Lipopolysaccharide-induced expression of intercellular adhesion molecule-1 and chemokines in cultured human corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2005, 46, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.C.; Heinzel, F.P.; Diaconu, E.; Sun, Y.; Hise, A.G.; Golenbock, D.; Lass, J.H.; Pearlman, E. Activation of toll-like receptor (TLR)2, TLR4, and TLR9 in the mammalian cornea induces MyD88-dependent corneal inflammation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Bonini, S.; Micera, A.; Iovieno, A.; Lambiase, A.; Bonini, S. Expression of Toll-like receptors in healthy and allergic conjunctiva. Ophthalmology 2005, 112, 1528, discussion 1548–1529. [Google Scholar] [CrossRef]
- Talreja, D.; Singh, P.K.; Kumar, A. In Vivo Role of TLR2 and MyD88 Signaling in Eliciting Innate Immune Responses in Staphylococcal Endophthalmitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1719–1732. [Google Scholar] [CrossRef] [Green Version]
- Novosad, B.D.; Astley, R.A.; Callegan, M.C. Role of Toll-like receptor (TLR) 2 in experimental Bacillus cereus endophthalmitis. PLoS ONE 2011, 6, e28619. [Google Scholar] [CrossRef] [Green Version]
- Mulfaul, K.; Rhatigan, M.; Doyle, S. Toll-Like Receptors and Age-Related Macular Degeneration. Adv. Exp. Med. Biol. 2018, 1074, 19–28. [Google Scholar] [CrossRef]
- Güven, M.; Batar, B.; Mutlu, T.; Bostancı, M.; Mete, M.; Aras, C.; Ünal, M. Toll-Like Receptors 2 and 4 Polymorphisms in Age-Related Macular Degeneration. Curr. Eye Res. 2016, 41, 856–861. [Google Scholar] [CrossRef]
- Yang, Z.; Stratton, C.; Francis, P.J.; Kleinman, M.E.; Tan, P.L.; Gibbs, D.; Tong, Z.; Chen, H.; Constantine, R.; Yang, X.; et al. Toll-like receptor 3 and geographic atrophy in age-related macular degeneration. N. Engl. J. Med. 2008, 359, 1456–1463. [Google Scholar] [CrossRef] [PubMed]
- Allikmets, R.; Bergen, A.A.; Dean, M.; Guymer, R.H.; Hageman, G.S.; Klaver, C.C.; Stefansson, K.; Weber, B.H. Geographic atrophy in age-Related macular degeneration and TLR3. N. Engl. J. Med. 2009, 360, 2255–2256. [Google Scholar]
- Edwards, A.O.; Swaroop, A.; Seddon, J.M. Geographic atrophy in age-Related macular degeneration and TLR3. N. Engl. J. Med. 2009, 360, 2255–2256. [Google Scholar]
- Liew, G.; Mitchell, P.; Wong, T.Y. Geographic atrophy in age-Related macular degeneration and TLR3. N. Engl. J. Med. 2009, 360, 2255–2256. [Google Scholar]
- Lewin, A.S. Geographic atrophy in age-Related macular degeneration and TLR3. N. Engl. J. Med. 2009, 360, 2255–2256. [Google Scholar] [CrossRef] [Green Version]
- Edwards, A.O.; Chen, D.; Fridley, B.L.; James, K.M.; Wu, Y.; Abecasis, G.; Swaroop, A.; Othman, M.; Branham, K.; Iyengar, S.K.; et al. Toll-like receptor polymorphisms and age-Related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1652–1659. [Google Scholar] [CrossRef]
- Cho, Y.; Wang, J.J.; Chew, E.Y.; Ferris, F.L., 3rd; Mitchell, P.; Chan, C.C.; Tuo, J. Toll-like receptor polymorphisms and age-related macular degeneration: Replication in three case-Control samples. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5614–5618. [Google Scholar] [CrossRef]
- Klein, M.L.; Ferris, F.L., 3rd; Francis, P.J.; Lindblad, A.S.; Chew, E.Y.; Hamon, S.C.; Ott, J. Progression of geographic atrophy and genotype in age-Related macular degeneration. Ophthalmology 2010, 117, 1554–1559. [Google Scholar] [CrossRef]
- Zhou, P.; Fan, L.; Yu, K.D.; Zhao, M.W.; Li, X.X. Toll-Like receptor 3 C1234T may protect against geographic atrophy through decreased dsRNA binding capacity. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 3489–3495. [Google Scholar] [CrossRef]
- Sharma, N.K.; Sharma, K.; Gupta, A.; Prabhakar, S.; Singh, R.; Gupta, P.K.; Anand, A. Does toll-like receptor-3 (TLR-3) have any role in Indian AMD phenotype? Mol. Cell. Biochem. 2014, 393, 1–8. [Google Scholar] [CrossRef]
- Ma, L.; Tang, F.Y.; Chu, W.K.; Young, A.L.; Brelen, M.E.; Pang, C.P.; Chen, L.J. Association of toll-like receptor 3 polymorphism rs3775291 with age-Related macular degeneration: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 19718. [Google Scholar] [CrossRef]
- Sng, C.C.; Cackett, P.D.; Yeo, I.Y.; Thalamuthu, A.; Venkatraman, A.; Venkataraman, D.; Koh, A.H.; Tai, E.S.; Wong, T.Y.; Aung, T.; et al. Toll-Like receptor 3 polymorphism rs3775291 is not associated with choroidal neovascularization or polypoidal choroidal vasculopathy in Chinese subjects. Ophthalmic Res. 2011, 45, 191–196. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, M.W.; Li, H.P.; Zeng, W.T.; Zhou, P.; Huang, L.Z.; Li, X.X.; Sun, Y.Y. Toll-Like receptor 3 polymorphism is not associated with neovascular age-related macular degeneration and polypoidal choroidal vasculopathy in the Chinese. Genet. Mol. Res. 2014, 13, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, J.E.; Pistilli, M.; Ying, G.S.; Maguire, M.G.; Daniel, E.; Martin, D.F. Growth of geographic atrophy in the comparison of age-related macular degeneration treatments trials. Ophthalmology 2015, 122, 809–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zareparsi, S.; Buraczynska, M.; Branham, K.E.; Shah, S.; Eng, D.; Li, M.; Pawar, H.; Yashar, B.M.; Moroi, S.E.; Lichter, P.R.; et al. Toll-like receptor 4 variant D299G is associated with susceptibility to age-Related macular degeneration. Hum. Mol. Genet. 2005, 14, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Despriet, D.D.; Bergen, A.A.; Merriam, J.E.; Zernant, J.; Barile, G.R.; Smith, R.T.; Barbazetto, I.A.; van Soest, S.; Bakker, A.; de Jong, P.T.; et al. Comprehensive analysis of the candidate genes CCL2, CCR2, and TLR4 in age-Related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2008, 49, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Kaur, I.; Hussain, A.; Hussain, N.; Das, T.; Pathangay, A.; Mathai, A.; Hussain, A.; Nutheti, R.; Nirmalan, P.K.; Chakrabarti, S. Analysis of CFH, TLR4, and APOE polymorphism in India suggests the Tyr402His variant of CFH to be a global marker for age-Related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3729–3735. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.C.; Guo, X.H.; Chen, X.; Yao, Y. Toll-Like receptor 4 gene polymorphisms rs4986790 and rs4986791 and age-Related macular degeneration susceptibility: A meta-Analysis. Ophthalmic Genet. 2020, 41, 31–35. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Huang, Y.Q.; Zhang, X.Y.; Zheng, P.F.; Li, P.; Chen, Y.; Shu, L. Association study of toll-Like receptors 4 polymorphisms and the risk of age-Related macular degeneration: A meta-Analysis. Ophthalmic Genet. 2020, 41, 579–584. [Google Scholar] [CrossRef]
- Ling, Y.; Xiong, F. Associations of TLR4 gene polymorphisms with the risk of age-Related macular degeneration in a Chinese Han population. Medicine 2019, 98, e15583. [Google Scholar] [CrossRef]
- Zhu, Y.; Liang, L.; Qian, D.; Yu, H.; Yang, P.; Lei, B.; Peng, H. Increase in peripheral blood mononuclear cell Toll-Like receptor 2/3 expression and reactivity to their ligands in a cohort of patients with wet age-related macular degeneration. Mol. Vis. 2013, 19, 1826–1833. [Google Scholar] [PubMed]
- Maloney, S.C.; Antecka, E.; Orellana, M.E.; Fernandes, B.F.; Odashiro, A.N.; Eghtedari, M.; Burnier, M.N., Jr. Choroidal neovascular membranes express toll-Like receptor 3. Ophthalmic Res. 2010, 44, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, T.; Sonoda, K.H.; Hijioka, K.; Sato, K.; Takeda, A.; Hasegawa, E.; Oshima, Y.; Ishibashi, T. Choroidal neovascularization enhanced by Chlamydia pneumoniae via Toll-Like receptor 2 in the retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4694–4702. [Google Scholar] [CrossRef] [Green Version]
- Will-Orrego, A.; Qiu, Y.; Fassbender, E.S.; Shen, S.; Aranda, J.; Kotagiri, N.; Maker, M.; Liao, S.M.; Jaffee, B.D.; Poor, S.H. Amount of Mononuclear Phagocyte Infiltrate Does Not Predict Area of Experimental Choroidal Neovascularization (CNV). J. Ocul. Pharmacol. Ther. Off. J. Assoc. Ocul. Pharmacol. Ther. 2018, 34, 489–499. [Google Scholar] [CrossRef]
- West, X.Z.; Malinin, N.L.; Merkulova, A.A.; Tischenko, M.; Kerr, B.A.; Borden, E.C.; Podrez, E.A.; Salomon, R.G.; Byzova, T.V. Oxidative stress induces angiogenesis by activating TLR2 with novel endogenous ligands. Nature 2010, 467, 972–976. [Google Scholar] [CrossRef]
- Wang, H.; Guo, J.; West, X.Z.; Bid, H.K.; Lu, L.; Hong, L.; Jang, G.-F.; Zhang, L.; Crabb, J.W.; Linetsky, M.; et al. Detection and biological activities of carboxyethylpyrrole ethanolamine phospholipids (CEP-EPs). Chem. Res. Toxicol. 2014, 27, 2015–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Meer, S.G.; Miyagi, M.; Rayborn, M.E.; Hollyfield, J.G.; Crabb, J.W.; Salomon, R.G. Carboxyethylpyrrole protein adducts and autoantibodies, biomarkers for age-Related macular degeneration. J. Biol. Chem. 2003, 278, 42027–42035. [Google Scholar] [CrossRef] [Green Version]
- Mulfaul, K.; Ozaki, E.; Fernando, N.; Brennan, K.; Chirco, K.R.; Connolly, E.; Greene, C.; Maminishkis, A.; Salomon, R.G.; Linetsky, M.; et al. Toll-Like Receptor 2 Facilitates Oxidative Damage-Induced Retinal Degeneration. Cell Rep. 2020, 30, 2209–2224.e2205. [Google Scholar] [CrossRef] [Green Version]
- Gounarides, J.; Cobb, J.S.; Zhou, J.; Cook, F.; Yang, X.; Yin, H.; Meredith, E.; Rao, C.; Huang, Q.; Xu, Y.; et al. Lack of involvement of CEP adducts in TLR activation and in angiogenesis. PLoS ONE 2014, 9, e111472. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.M.; Duffort, S.; Ivanov, D.; Wang, H.; Laird, J.M.; Salomon, R.G.; Cruz-Guilloty, F.; Perez, V.L. The oxidative stress product carboxyethylpyrrole potentiates TLR2/TLR1 inflammatory signaling in macrophages. PLoS ONE 2014, 9, e106421. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Ju, M.; Lee, K.Y.V.; Mackey, A.; Evangelista, M.; Iwata, D.; Adamson, P.; Lashkari, K.; Foxton, R.; Shima, D.; et al. A Proinflammatory Function of Toll-Like Receptor 2 in the Retinal Pigment Epithelium as a Novel Target for Reducing Choroidal Neovascularization in Age-Related Macular Degeneration. Am. J. Pathol. 2017, 187, 2208–2221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, Y.; Matsumoto, H.; Roh, M.; Giani, A.; Kataoka, K.; Morizane, Y.; Kayama, M.; Thanos, A.; Nakatake, S.; Notomi, S.; et al. Programmed necrosis, not apoptosis, is a key mediator of cell loss and DAMP-Mediated inflammation in dsRNA-induced retinal degeneration. Cell Death Differ. 2014, 21, 270–277. [Google Scholar] [CrossRef]
- Chintala, S.K.; Putris, N.; Geno, M. Activation of TLR3 promotes the degeneration of retinal ganglion cells by upregulating the protein levels of JNK3. Investig. Ophthalmol. Vis. Sci. 2015, 56, 505–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiose, S.; Chen, Y.; Okano, K.; Roy, S.; Kohno, H.; Tang, J.; Pearlman, E.; Maeda, T.; Palczewski, K.; Maeda, A. Toll-Like receptor 3 is required for development of retinopathy caused by impaired all-Trans-Retinal clearance in mice. J. Biol. Chem. 2011, 286, 15543–15555. [Google Scholar] [CrossRef] [Green Version]
- Klettner, A.; Koinzer, S.; Meyer, T.; Roider, J. Toll-Like receptor 3 activation in retinal pigment epithelium cells-Mitogen-Activated protein kinase pathways of cell death and vascular endothelial growth factor secretion. Acta Ophthalmol. 2013, 91, e211–e218. [Google Scholar] [CrossRef]
- Patel, A.K.; Hackam, A.S. A novel protective role for the innate immunity Toll-Like Receptor 3 (TLR3) in the retina via Stat3. Mol. Cell. Neurosci. 2014, 63, 38–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.K.; Hackam, A.S. Toll-like receptor 3 (TLR3) protects retinal pigmented epithelium (RPE) cells from oxidative stress through a STAT3-dependent mechanism. Mol. Immunol. 2013, 54, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Kleinman, M.E.; Yamada, K.; Takeda, A.; Chandrasekaran, V.; Nozaki, M.; Baffi, J.Z.; Albuquerque, R.J.; Yamasaki, S.; Itaya, M.; Pan, Y.; et al. Sequence- and target-independent angiogenesis suppression by siRNA via TLR3. Nature 2008, 452, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Ohno-Matsui, K.; Ichinose, S.; Sato, T.; Iwata, N.; Saido, T.C.; Hisatomi, T.; Mochizuki, M.; Morita, I. The potential role of amyloid beta in the pathogenesis of age-Related macular degeneration. J. Clin. Investig. 2005, 115, 2793–2800. [Google Scholar] [CrossRef]
- Chen, L.; Bai, Y.; Zhao, M.; Jiang, Y. TLR4 inhibitor attenuates amyloid-beta-induced angiogenic and inflammatory factors in ARPE-19 cells: Implications for age-Related macular degeneration. Mol. Med. Rep. 2016, 13, 3249–3256. [Google Scholar] [CrossRef] [Green Version]
- Javitt, N.B.; Javitt, J.C. The retinal oxysterol pathway: A unifying hypothesis for the cause of age-Related macular degeneration. Curr. Opin. Ophthalmol. 2009, 20, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.D.; Amaral, J.; Lee, J.W.; Rodriguez, I.R. 7-Ketocholesterol-induced inflammation signals mostly through the TLR4 receptor both in vitro and in vivo. PLoS ONE 2014, 9, e100985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohno, H.; Chen, Y.; Kevany, B.M.; Pearlman, E.; Miyagi, M.; Maeda, T.; Palczewski, K.; Maeda, A. Photoreceptor proteins initiate microglial activation via Toll-Like receptor 4 in retinal degeneration mediated by all-Trans-Retinal. J. Biol. Chem. 2013, 288, 15326–15341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowley, M.A.; Delgado, O.; Will-Orrego, A.; Buchanan, N.M.; Anderson, K.; Jaffee, B.D.; Dryja, T.P.; Liao, S.M. Induction of Ocular Complement Activation by Inflammatory Stimuli and Intraocular Inhibition of Complement Factor D in Animal Models. Investig. Ophthalmol. Vis. Sci. 2018, 59, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Takeda, A.; Yoshimura, T.; Oshima, Y.; Sonoda, K.H.; Ishibashi, T. IL-10 is significantly involved in HSP70-regulation of experimental subretinal fibrosis. PLoS ONE 2013, 8, e80288. [Google Scholar] [CrossRef]
- Kumar, M.V.; Nagineni, C.N.; Chin, M.S.; Hooks, J.J.; Detrick, B. Innate immunity in the retina: Toll-Like receptor (TLR) signaling in human retinal pigment epithelial cells. J. Neuroimmunol. 2004, 153, 7–15. [Google Scholar] [CrossRef]
- Duncan, R.S.; Rohowetz, L.; Vogt, A.; Koulen, P. Repeat exposure to polyinosinic:polycytidylic acid induces TLR3 expression via JAK-STAT signaling and synergistically potentiates NFκB-RelA signaling in ARPE-19 cells. Cell Signal. 2020, 66, 109494. [Google Scholar] [CrossRef]
- Wörnle, M.; Merkle, M.; Wolf, A.; Ribeiro, A.; Himmelein, S.; Kernt, M.; Kampik, A.; Eibl-Lindner, K.H. Inhibition of TLR3-Mediated proinflammatory effects by Alkylphosphocholines in human retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6536–6544. [Google Scholar] [CrossRef]
- Brosig, A.; Kuhrt, H.; Wiedemann, P.; Kohen, L.; Bringmann, A.; Hollborn, M. Gene expression regulation in retinal pigment epithelial cells induced by viral RNA and viral/bacterial DNA. Mol. Vis. 2015, 21, 1000–1016. [Google Scholar]
- Paimela, T.; Ryhänen, T.; Mannermaa, E.; Ojala, J.; Kalesnykas, G.; Salminen, A.; Kaarniranta, K. The effect of 17beta-Estradiol on IL-6 secretion and NF-Kappab DNA-Binding activity in human retinal pigment epithelial cells. Immunol. Lett. 2007, 110, 139–144. [Google Scholar] [CrossRef]
- Zhu, Y.; Dai, B.; Li, Y.; Peng, H. C5a and toll-Like receptor 4 crosstalk in retinal pigment epithelial cells. Mol. Vis. 2015, 21, 1122–1129. [Google Scholar]
- Terheyden, L.; Roider, J.; Klettner, A. Basolateral activation with TLR agonists induces polarized cytokine release and reduces barrier function in RPE in vitro. Graefe’s Arch. Clin. Exp. Ophthalmol. 2021, 259, 413–424. [Google Scholar] [CrossRef]
- Dietrich, L.; Lucius, R.; Roider, J.; Klettner, A. Interaction of inflammatorily activated retinal pigment epithelium with retinal microglia and neuronal cells. Exp. Eye Res. 2020, 199, 108167. [Google Scholar] [CrossRef] [PubMed]
- Hettich, C.; Wilker, S.; Mentlein, R.; Lucius, R.; Roider, J.; Klettner, A. The retinal pigment epithelium (RPE) induces FasL and reduces iNOS and Cox2 in primary monocytes. Graefe’s Arch. Clin. Exp. Ophthalmol. 2014, 252, 1747–1754. [Google Scholar] [CrossRef] [PubMed]
- Klettner, A.; Brinkmann, A.; Winkelmann, K.; Käckenmeister, T.; Hildebrandt, J.; Roider, J. Effect of long-Term inflammation on viability and function of RPE cells. Exp. Eye Res. 2020, 200, 108214. [Google Scholar] [CrossRef]
- Elner, S.G.; Petty, H.R.; Elner, V.M.; Yoshida, A.; Bian, Z.M.; Yang, D.; Kindzelskii, A.L. TLR4 mediates human retinal pigment epithelial endotoxin binding and cytokine expression. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4627–4633. [Google Scholar] [CrossRef]
- Chen, C.; Guo, D.; Lu, G. Wogonin protects human retinal pigment epithelium cells from LPS-Induced barrier dysfunction and inflammatory responses by regulating the TLR4/NF-κB signaling pathway. Mol. Med. Rep. 2017, 15, 2289–2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindzelskii, A.L.; Elner, V.M.; Elner, S.G.; Yang, D.; Hughes, B.A.; Petty, H.R. Toll-Like receptor 4 (TLR4) of retinal pigment epithelial cells participates in transmembrane signaling in response to photoreceptor outer segments. J. Gen. Physiol. 2004, 124, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Karlstetter, M.; Scholz, R.; Rutar, M.; Wong, W.T.; Provis, J.M.; Langmann, T. Retinal microglia: Just bystander or target for therapy? Prog. Retin. Eye Res. 2015, 45, 30–57. [Google Scholar] [CrossRef]
- Apte, R.S.; Richter, J.; Herndon, J.; Ferguson, T.A. Macrophages inhibit neovascularization in a murine model of age-Related macular degeneration. PLoS Med. 2006, 3, e310. [Google Scholar] [CrossRef] [Green Version]
- Klettner, A.; Hamann, T.; Schluter, K.; Lucius, R.; Roider, J. Retinal pigment epithelium cells alter the pro-inflammatory response of retinal microglia to TLR-3 stimulation. Acta Ophthalmol. 2014, 92, e621–e629. [Google Scholar] [CrossRef]
- Ghasemi, H.; Ghazanfari, T.; Yaraee, R.; Faghihzadeh, S.; Hassan, Z.M. Roles of IL-8 in ocular inflammations: A review. Ocul. Immunol. Inflamm. 2011, 19, 401–412. [Google Scholar] [CrossRef]
- Tezel, G. TNF-Alpha signaling in glaucomatous neurodegeneration. Prog. Brain Res. 2008, 173, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Jonas, J.B.; Tao, Y.; Neumaier, M.; Findeisen, P. Cytokine concentration in aqueous humour of eyes with exudative age-Related macular degeneration. Acta Ophthalmol. 2012, 90, e381–e388. [Google Scholar] [CrossRef] [PubMed]
- Miao, H.; Tao, Y.; Li, X.X. Inflammatory cytokines in aqueous humor of patients with choroidal neovascularization. Mol. Vis. 2012, 18, 574–580. [Google Scholar] [PubMed]
- Kleinman, M.E.; Kaneko, H.; Cho, W.G.; Dridi, S.; Fowler, B.J.; Blandford, A.D.; Albuquerque, R.J.; Hirano, Y.; Terasaki, H.; Kondo, M.; et al. Short-Interfering RNAs induce retinal degeneration via TLR3 and IRF3. Mol. Ther. J. Am. Soc. Gene Ther. 2012, 20, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tode, J.; Richert, E.; Koinzer, S.; Klettner, A.; von der Burchard, C.; Brinkmann, R.; Lucius, R.; Roider, J. Thermal Stimulation of the Retina Reduces Bruch’s Membrane Thickness in Age Related Macular Degeneration Mouse Models. Transl. Vis. Sci. Technol. 2018, 7, 2. [Google Scholar] [CrossRef]
- Dithmar, S.; Curcio, C.A.; Le, N.A.; Brown, S.; Grossniklaus, H.E. Ultrastructural changes in Bruch’s membrane of apolipoprotein E-Deficient mice. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2035–2042. [Google Scholar]
- Zhao, Z.; Chen, Y.; Wang, J.; Sternberg, P.; Freeman, M.L.; Grossniklaus, H.E.; Cai, J. Age-related retinopathy in NRF2-Deficient mice. PLoS ONE 2011, 6, e19456. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| TLR | Agonist | Effect | Reference |
|---|---|---|---|
| TLR2 | Chlamydia pneumonia | Secretion IL-6, VEGF | [65] |
| Expression MCP-1, IL-1ß, IL-8 | [73] | ||
| PAM2CSK4 | Expression CFB, C3 | [70] | |
| Expression/secretion IL-6, IL-1ß, IL-8, MCP1, TNFα | [73,94,95] | ||
| Decreased barrier function, reduced expression tight junction | [94] | ||
| TLR3 | Poly I:C | Expression/secretion IL-6, IL-1ß, IL-8, TNFα, MCP-1, sICAM-1, IFN-ß | [25,88,90,91,94,95,97] |
| Expression HIF-1α, Jam-1, ICAM-1, bFGF, C5, C9, CFB | [25,90,91] | ||
| VEGF | [77] | ||
| Activation of ERK1/2, p38, JNK | [77,91] | ||
| Reduced barrier function | [94] | ||
| Induced cell death | [43,74,76,77,97] | ||
| Poly I:C + paraquat | Protection from oxidative stress | [78,79] | |
| TLR4 | LPS | Expression/secretion IL-8, IL-6, TNFα, IL-1ß | [94,97,98,99] |
| Expression COX-2, iNOS | [99] | ||
| Reduced barrier function | [94,99] | ||
| Reduced cell viability | [97] | ||
| Reduced phagocytosis | [97] | ||
| Reduced expression RPE65 | [97] | ||
| TLR9 | CpG-DNA | Secretion IL-8 | [25] |
| Increased phagocytosis | [25] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klettner, A.; Roider, J. Retinal Pigment Epithelium Expressed Toll-like Receptors and Their Potential Role in Age-Related Macular Degeneration. Int. J. Mol. Sci. 2021, 22, 8387. https://doi.org/10.3390/ijms22168387
Klettner A, Roider J. Retinal Pigment Epithelium Expressed Toll-like Receptors and Their Potential Role in Age-Related Macular Degeneration. International Journal of Molecular Sciences. 2021; 22(16):8387. https://doi.org/10.3390/ijms22168387
Chicago/Turabian StyleKlettner, Alexa, and Johann Roider. 2021. "Retinal Pigment Epithelium Expressed Toll-like Receptors and Their Potential Role in Age-Related Macular Degeneration" International Journal of Molecular Sciences 22, no. 16: 8387. https://doi.org/10.3390/ijms22168387
APA StyleKlettner, A., & Roider, J. (2021). Retinal Pigment Epithelium Expressed Toll-like Receptors and Their Potential Role in Age-Related Macular Degeneration. International Journal of Molecular Sciences, 22(16), 8387. https://doi.org/10.3390/ijms22168387