
Abiotrophia defectiva DnaK Promotes Fibronectin-Mediated Adherence to HUVECs and Induces a Proinflammatory Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
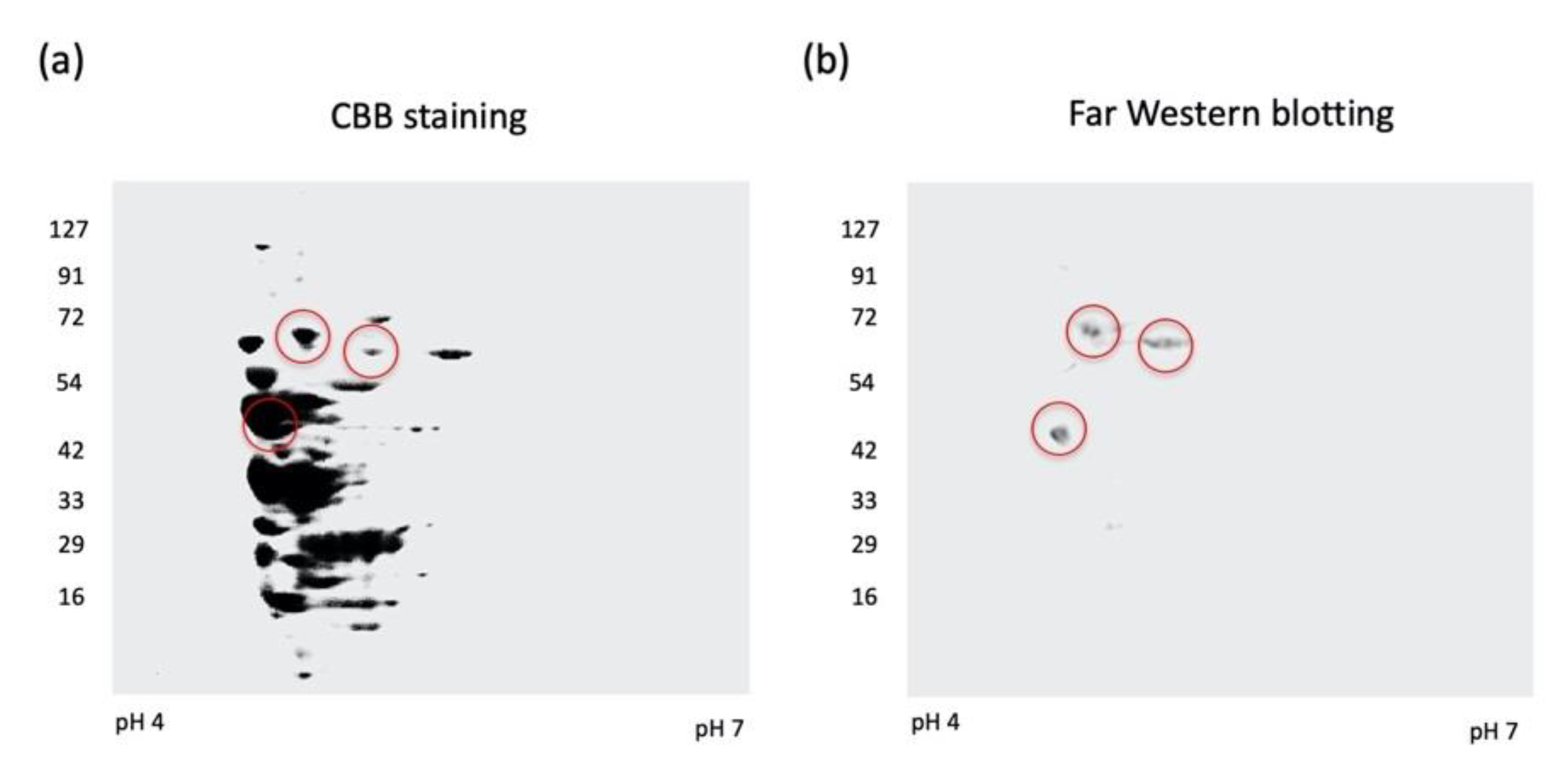
2.1. Identification of Fibronectin-Binding Molecules in A. defectiva Cells with Two-Dimensional Electrophoresis and Far-Western Blotting
2.2. Binding of rDnaK to Immobilized Fibronectin and the Effect of Anti-Dnak Antiserum on the Binding of A. defectiva to Immobilized Fibronectin or HUVECs
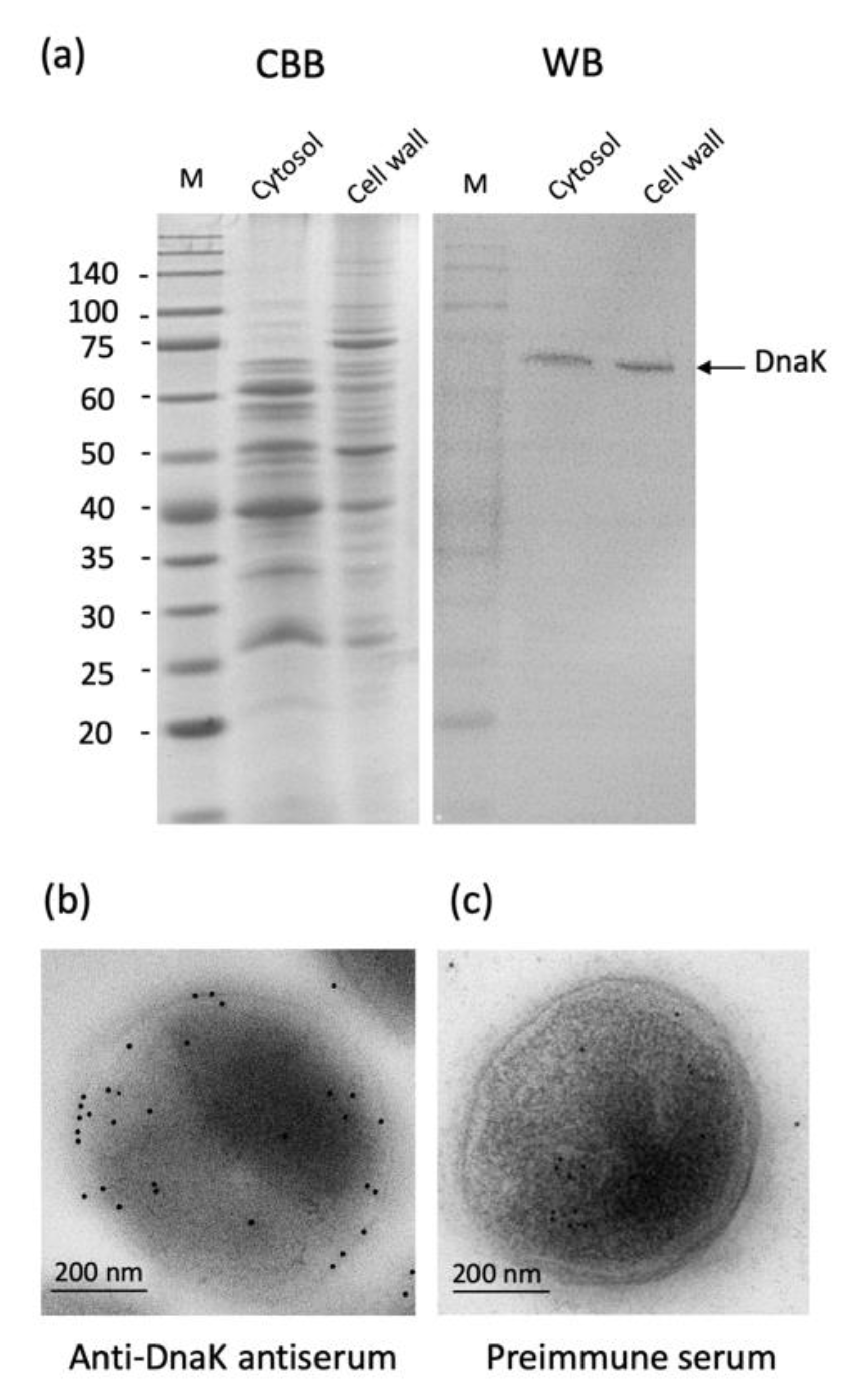
2.3. Localization of A. defectiva DnaK
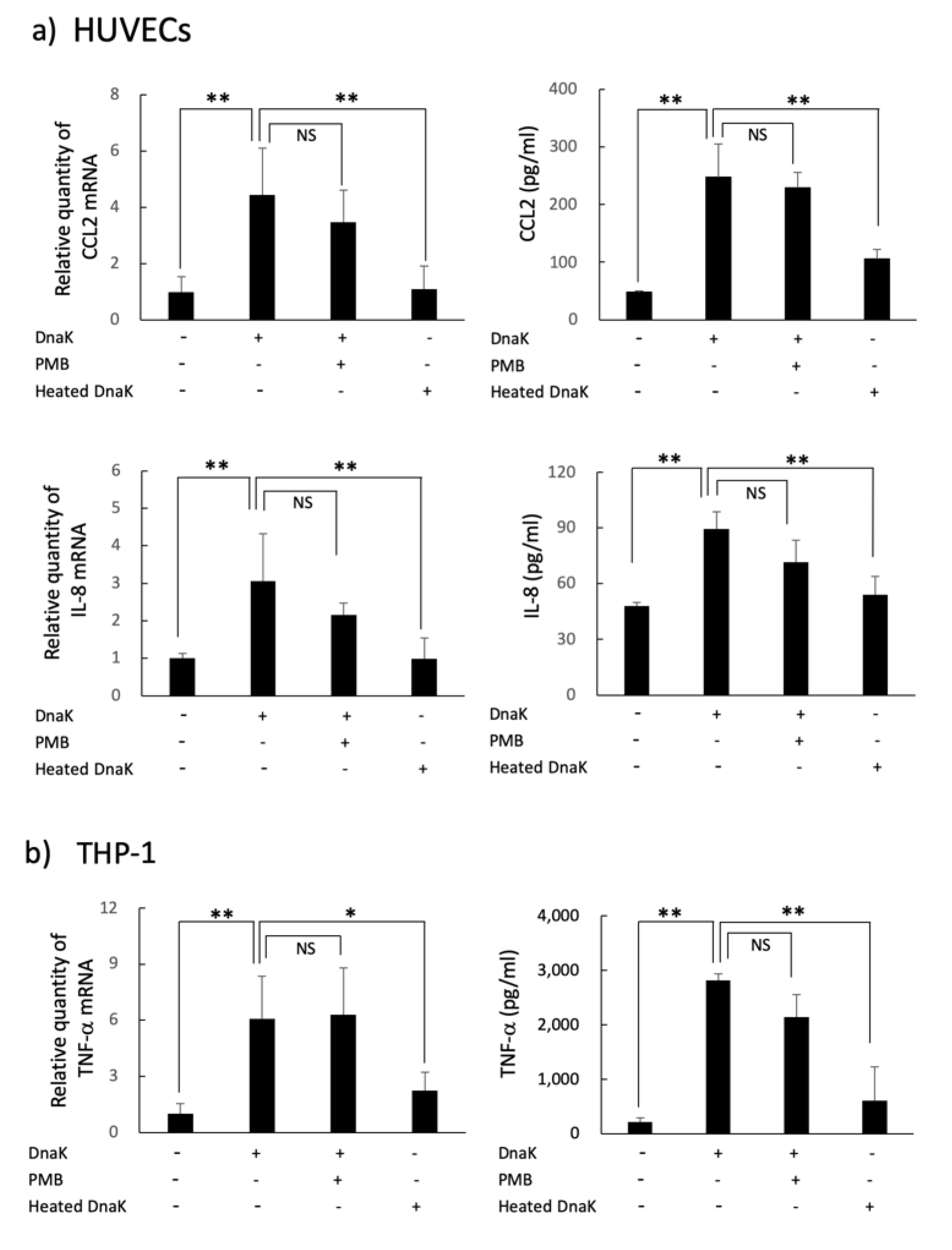
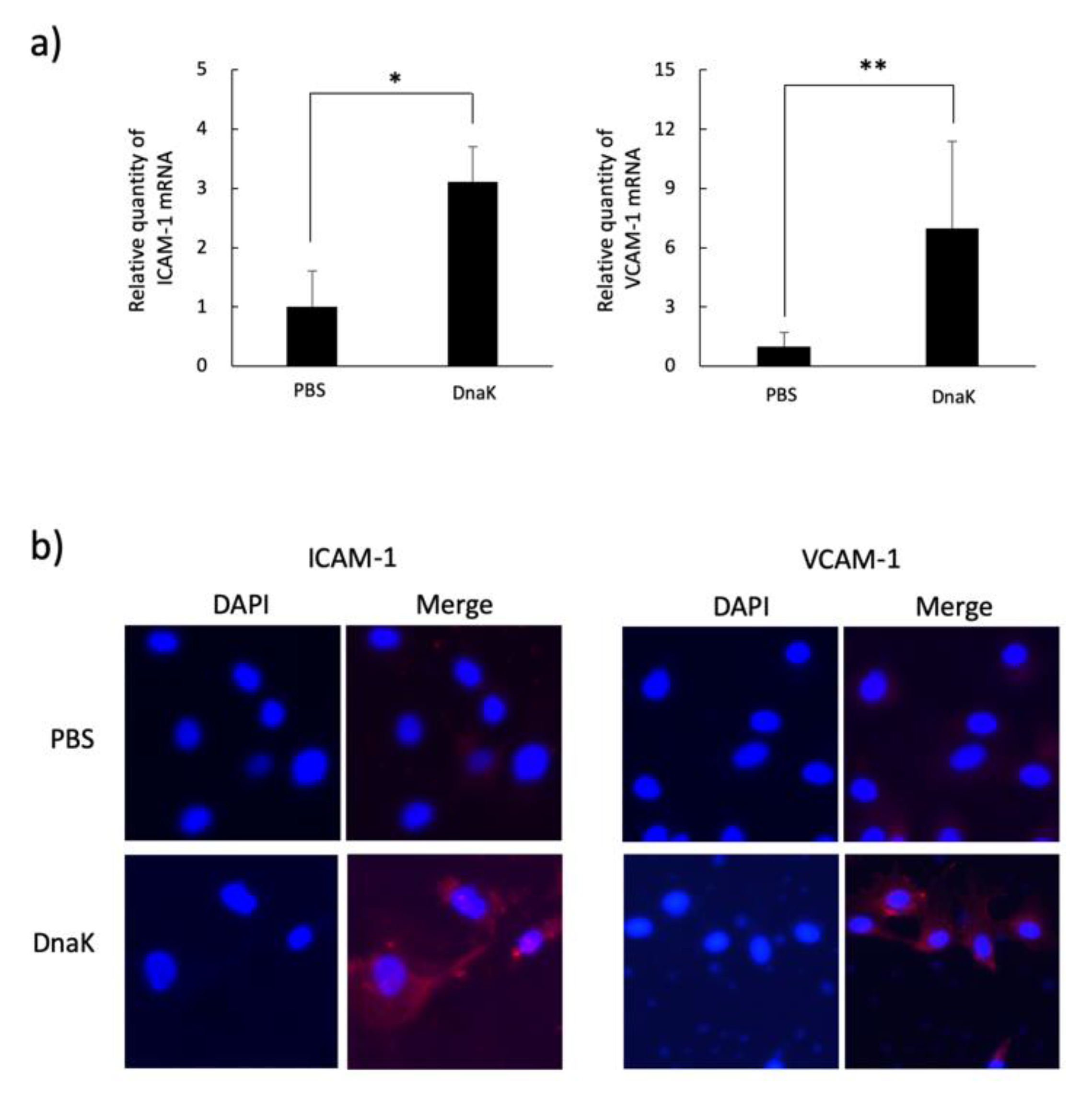
2.4. Cytokines and Adhesion Molecules Induction in HUVECs or THP-1 Cells Stimulated with A. defectiva rDnaK
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Cultured Cells and Culture Conditions
4.3. Two-Dimensional Electrophoresis and Far-Western Blotting
4.4. Identification of a 66-kDa Protein
4.5. Construction of Expression Vector for A. defectiva Dnak Protein and Expression and Purification of rDnak
4.6. Binding of rDnaK to Immobilized Fibronectin
4.7. Binding of A. defectiva to Immobilized Fibronectin or HUVECs
4.8. Localization of A. defectiva DnaK
4.9. Induction of Cytokines and Adhesion Molecules in HUVECs or THP-1 Cells Stimulated with A. defectiva rDnaK
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frenkel, A.; Hirsch, W. Spontaneous development of L forms of streptococci requiring secretions of other bacteria or sulphydryl compounds for normal growth. Nature 1961, 191, 728–730. [Google Scholar] [CrossRef]
- Takeshita, T.; Yasui, M.; Shibata, Y.; Furuta, M.; Saeki, Y.; Eshima, N.; Yamashita, Y. Dental plaque development on a hydroxyapatite disk in young adults observed by using a barcoded pyrosequencing approach. Sci. Rep. 2015, 5, 8136. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.A.; Levine, M.J.; Reddy, M.S.; Tabak, L.A.; Bergey, E.J. Preparation of a sialic acid-binding protein from Streptococcus mitis KS32AR. Infect. Immun. 1986, 53, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, H.F.; Demuth, D.R. Structure, function and immunogenicity of streptococcal antigen I/II polypeptides. Mol. Microbiol. 1997, 23, 183–190. [Google Scholar] [CrossRef]
- Sasaki, M.; Kodama, Y.; Shimoyama, Y.; Ishikawa, T.; Tajika, S.; Kimura, S. Abiotrophia defectiva adhere to saliva-coated hydroxyapatite beads via interactions between salivary proline-rich-proteins and bacterial glycerlaldehyde-3-phosphate dehydrogenase. Microbiol. Immunol. 2020, 64, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Shimoyama, Y.; Ishikawa, T.; Kodama, Y.; Tajika, S.; Kimura, S. Contribution of different adherent properties of Granulicatella adiacens and Abiotrophia defectiva to their associations with oral colonization and the risk of infective endocarditis. J. Oral Sci. 2020, 62, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Senn, L.; Entenza, J.M.; Prod’hom, G. Adherence of Abiotrophia defectiva and Granulicatella species to fibronectin: Is there a link with endovascular infections? FEMS Immunol. Med. Microbiol. 2006, 48, 215–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, R.A. The staphylococcal fibronectin receptor: Evidence for its importance in invasive infections. Rev. Infect. Dis. 1987, 9, S335–S340. [Google Scholar] [CrossRef] [PubMed]
- Hanski, E.; Caparon, M. Protein F, a fibronectin-binding protein, is an adhesin of the group A streptococcus pyogenes. Proc. Natl. Acad. Sci. USA 1992, 89, 6172–6176. [Google Scholar] [CrossRef] [Green Version]
- Xolalpa, W.; Vallecillo, A.J.; Lara, M.; Mendoza-Hernandez, G.; Comini, M.; Spallek, R.; Singh, M.; Espitia, C. Identification of novel bacterial plasminogen-binding proteins in the human pathogen Mycobacterium tuberculosis. Proteomics 2007, 7, 3332–3341. [Google Scholar] [CrossRef]
- Kalayoglu, M.V.; Morrison, R.P.I.; Morrison, S.G.; Yuan, Y.; Byrne, G.I. Chlamydial virulence determinants in atherogenesis: The role of chlamydial lipopolysaccharide and heat shock protein 60 in macrophage-lipoprotein interactions. J. Infect. Dis. 2000, 181, S483–S489. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, S.; Yamaguchi, H.; Osaki, T.; Taguchi, H. A virulence factor of Helicobacter pylori: Role of heat shock protein in mucosal inflammation after H. pylori infection. J. Clin. Gastroenterol. 1998, 27, S35–S39. [Google Scholar] [CrossRef]
- Wang, Y.; Kelly, C.G.; Karttunen, J.T.; Whittall, T.; Lehner, P.J.; Duncan, L.; MacAry, P.; Younson, J.S.; Singh, M.; Oehlmann, W.; et al. CD40 is a cellular receptor mediating mycobacterial heat shock protein 70 stimulation of CC-chemokines. Immunity 2001, 15, 971–983. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yokota, K.; Ayada, K.; Yamamoto, Y.; Okada, T.; Shen, L.; Oguma, K. Helicobacter pylori heat-shock protein 60 induces interleukin-8 via a toll-like receptor (TLR) 2 and mitogen-activated protein (MAP) kinase pathway in human monocytes. J. Med. Microbiol. 2007, 56, 154–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabona, P.; Reddi, K.; Khan, S.; Nair, S.P.; Crean, S.J.; Meghji, S.; Wilson, M.; Preuss, M.; Miller, A.D.; Poole, S.; et al. Homogeneous Escherichia coli chaperonin 60 induces IL-1 and IL-6 gene expression in human monocytes by a mechanism independent of protein conformation. J. Immunol. 1998, 161, 1414–1421. [Google Scholar] [PubMed]
- Roux, M.E.; Lecoq, D.; Meyer, D.; Dosne, A.M. Requirement of prestimulated THP-1 monocytic cells for endothelial cell activation. Involvement of TNF alpha. Blood Coagul. Fibrinolysis 1997, 8, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chu, C.F.; Wang, C.N.; Wu, H.T.; Bi, K.W.; Pang, J.H.; Huang, S.T. The anti-atherosclerotic effect of tanshinone IIA is associated with the inhibition of TNF-α-induced VCAM-1, ICAM-1 and CX3CL1 expression. Phytomedicine 2014, 21, 207–216. [Google Scholar] [CrossRef]
- Guimaraes, A.O.; Cao, Y.; Hong, K.; Mayba, O.; Peck, M.C.; Gutierrez, J.; Ruffin, F.; Carrasco-Triguero, M.; Dinoso, J.B.; Clemenzi-Allen, A.; et al. A prognostic model of persistent bacteremia and mortality in complicated Staphylococcus aureus bloodstream infection. Clin. Infect. Dis. 2019, 68, 1502–1511. [Google Scholar] [CrossRef]
- Schwarz-Linek, U.; Höök, M.; Potts, J.R. The molecular basis of fibronectin-mediated bacterial adherence to host cells. Mol. Microbiol. 2007, 52, 631–641. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Terao, Y.; Kawabata, S. Pleiotropic virulence factor–Streptococcus pyogenes fibronectin-binding proteins. Cell Microbiol. 2013, 15, 503–511. [Google Scholar] [CrossRef]
- Herman-Bausier, P.; El-Kirat-Chatel, S.; Foster, T.J.; Geoghegan, J.A.; Dufrêne, Y.F. Staphylococcus aureus fibronectin-binding protein A mediates cell-cell adhesion through low-affinity homophilic bonds. mBio 2015, 6, e00413–e00415. [Google Scholar] [CrossRef] [Green Version]
- Kodama, Y.; Ishikawa, T.; Shimoyama, Y.; Sasaki, D.; Kimura, S.; Sasaki, M. The fibronectin-binding protein homologue Fbp62 of Streptococcus anginosus is a potent virulence factor. Microbiol. Immunol. 2018, 62, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Kodama, Y.; Shimoyama, Y.; Ishikawa, T.; Kimura, S.; Sasaki, M. Characterization and pathogenicity of fibronectin binding protein FbpI of Streptococcus intermedius. Arch. Microbiol. 2020, 202, 2071–2081. [Google Scholar] [CrossRef]
- Tart, R.C.; van de Rijn, I. Identification of the surface component of Streptococcus defectivus that mediates extracellular matrix adherence. Infect. Immun. 1993, 61, 4994–5000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senn, L.; Entenza, J.M.; Greub, G.; Jaton, K.; Wenger, A.; Bille, J.; Calandra, T.; Prod’hom, G. Bloodstream and endovascular infections due to Abiotrophia defectiva and Granulicatella species. BMC Infect. Dis. 2006, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayaraman, G.C.; Burne, R.A. DnaK expression in response to heat shock of Streptococcus mutans. FEMS Microbiol. Lett. 1995, 131, 255–261. [Google Scholar] [CrossRef]
- Hecker, M.; Schumann, W.; Völker, U. Heat-shock and general stress response in Bacillus subtilis. Mol. Microbiol. 1996, 19, 417–428. [Google Scholar] [CrossRef]
- Schaumburg, J.; Diekmann, O.; Hagendorff, P.; Bergmann, S.; Rohde, M.; Hammerschmidt, S.; Jänsch, L.; Wehland, J.; Kärst, U. The cell wall subproteome of Listeria monocytogenes. Proteomics 2004, 4, 2991–3006. [Google Scholar] [CrossRef] [PubMed]
- Floto, R.A.; MacAry, P.A.; Boname, J.M.; Mien, T.S.; Kampmann, B.; Hair, J.R.; Huey, O.S.; Houben, E.N.; Pieters, J.; Day, C.; et al. Dendritic cell stimulation by mycobacterial Hsp70 is mediated through CCR5. Science 2006, 314, 454–458. [Google Scholar] [CrossRef]
- Basu, D.; Khare, G.; Singh, S.; Tyagi, A.; Khosla, S.; Mande, S.C. A novel nucleoid-associated protein of Mycobacterium tuberculosis is a sequence homolog of GroEL. Nucleic Acids Res. 2009, 37, 4944–4954. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Henderson, B.; Lund, P.A.; Tormay, P.; Ahmed, M.T.; Gurcha, S.S.; Besra, G.S.; Coates, A.R. A Mycobacterium tuberculosis mutant lacking the groEL homologue cpn60.1 is viable but fails to induce an inflammatory response in animal models of infection. Infect. Immun. 2008, 76, 1535–1546. [Google Scholar] [CrossRef] [Green Version]
- Kol, A.; Sukhova, G.K.; Lichtman, A.H.; Libby, P. Chlamydial heat shock protein 60 localizes in human atheroma and regulates macrophage tumor necrosis factor-alpha and matrix metalloproteinase expression. Circulation 1998, 98, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Kuppner, M.C.; van Meir, E.; Hamou, M.F.; de Tribolet, N. Cytokine regulation of intercellular adhesion molecule-1 (ICAM-1) expression on human glioblastoma cells. Clin. Exp. Immunol. 1990, 81, 142–148. [Google Scholar] [CrossRef]
- McHale, J.F.; Harari, O.A.; Marshall, D.; Haskard, D.O. TNF-alpha and IL-1 sequentially induce endothelial ICAM-1 and VCAM-1 expression in MRL/lpr lupus-prone mice. J. Immunol. 1999, 163, 3993–4000. [Google Scholar]
- Klemke, M.; Weschenfelder, T.; Konstandin, M.H.; Samstag, Y. High affinity interaction of integrin alpha4beta1 (VLA-4) and vascular cell adhesion molecule 1 (VCAM-1) enhances migration of human melanoma cells across activated endothelial cell layers. J. Cell Physiol. 2007, 212, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Lee, Y.; Yu, H.; Ha, U.H. HSP70-homolog DnaK of Pseudomonas aeruginosa increases the production of IL-27 through expression of EBI3 via TLR4-dependent NF-κB and TLR4-independent Akt signaling. Int. J. Mol. Sci. 2020, 21, 9194. [Google Scholar] [CrossRef]
- Ashtekar, A.R.; Zhang, P.; Katz, J.; Deivanayagam, C.C.; Rallabhandi, P.; Vogel, S.N.; Michalek, S.M. TLR4-mediated activation of dendritic cells by the heat shock protein DnaK from Francisella tularensis. J. Leukoc. Biol. 2008, 84, 1434–1446. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Kelly, C.G.; Singh, M.; Wang, Y.; Kelly, C.G.; Singh, M.; McGowan, E.G.; Carrara, A.S.; Bergmeier, L.A.; Lehner, T. Stimulation of Th1-polarizing cytokines, C-C chemokines, maturation of dendritic cells, and adjuvant function by the peptide binding fragment of heat shock protein 70. J. Immunol. 2002, 169, 2422–2429. [Google Scholar] [CrossRef] [Green Version]
- Rivera, J.; Cordero, R.J.; Nakouzi, A.S.; Frases, S.; Nicola, A.; Casadevall, A. Bacillus anthracis produces membrane-derived vesicles containing biologically active toxins. Proc. Natl. Acad. Sci. USA 2010, 107, 19002–19007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veith, P.D.; Chen, Y.Y.; Gorasia, D.G.; Chen, D.; Glew, M.D.; O’Brien-Simpson, N.M.; Cecil, J.D.; Holden, J.A.; Reynolds, E.C. Porphyromonas gingivalis outer membrane vesicles exclusively contain outer membrane and periplasmic proteins and carry a cargo enriched with virulence factors. J. Proteome Res. 2014, 13, 2420–2432. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, N.; Shimizu, H.; Fujimoto, K.; Watanabe, K.; Yamasaki, T.; Hatano, N.; Tamai, E.; Katayama, S.; Hitsumoto, Y. Expression of glyceraldehyde-3-phosphate dehydrogenase on the surface of Clostridium perfringens cells. Anaerobe 2018, 51, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y.; Ishikawa, T.; Kodama, Y.; Kimura, S.; Sasaki, M. Tyrosine tRNA synthetase as a novel extracellular immunomodulatory protein in Streptococcus anginosus. FEMS Microbiol. Lett. 2020, 367, fnaa153. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.R.; McNab, R.; Millsap, K.W.; Rohde, M.; Hammerschmidt, S.; Mawdsley, J.L.; Jenkinson, H.F. The pavA gene of Streptococcus pneumoniae encodes a fibronectin-binding protein that is essential for virulence. Mol. Microbiol. 2001, 41, 1395–1408. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, M.; Shimoyama, Y.; Kodama, Y.; Ishikawa, T. Abiotrophia defectiva DnaK Promotes Fibronectin-Mediated Adherence to HUVECs and Induces a Proinflammatory Response. Int. J. Mol. Sci. 2021, 22, 8528. https://doi.org/10.3390/ijms22168528
Sasaki M, Shimoyama Y, Kodama Y, Ishikawa T. Abiotrophia defectiva DnaK Promotes Fibronectin-Mediated Adherence to HUVECs and Induces a Proinflammatory Response. International Journal of Molecular Sciences. 2021; 22(16):8528. https://doi.org/10.3390/ijms22168528
Chicago/Turabian StyleSasaki, Minoru, Yu Shimoyama, Yoshitoyo Kodama, and Taichi Ishikawa. 2021. "Abiotrophia defectiva DnaK Promotes Fibronectin-Mediated Adherence to HUVECs and Induces a Proinflammatory Response" International Journal of Molecular Sciences 22, no. 16: 8528. https://doi.org/10.3390/ijms22168528
APA StyleSasaki, M., Shimoyama, Y., Kodama, Y., & Ishikawa, T. (2021). Abiotrophia defectiva DnaK Promotes Fibronectin-Mediated Adherence to HUVECs and Induces a Proinflammatory Response. International Journal of Molecular Sciences, 22(16), 8528. https://doi.org/10.3390/ijms22168528