1. Introduction
Radiotherapy is a conventional and effective treatment for neoplasm in a clinical context. More than 50 percent of tumor patients receive radiotherapy for curative or palliative purposes [
1]. During the radiotherapy of patients with abdominal or pelvic tumors, the small and large bowel is one of the crucial organs at risk during radiotherapy delivered to the abdomen and/or pelvis. A course of clinical radiotherapy usually lasts 4–6 weeks and the average exposure dose is about 50 Gy. As high dose of ionizing radiation (IR) can kill intestinal epithelial cells, intestinal absorption and barrier function may be destroyed and result in acute intestinal injury [
2]. The major clinical manifestations of intestinal injury include abdominal pain, diarrhea, and water electrolyte imbalance. In severe cases, intestinal fistula, intestinal stenosis, intestinal obstruction, may occur and even be life-threatening. Thus, IR-caused intestinal injury limits the effective radiation dosage used to eradicate tumors and adversely affects the curative effect.
Symptomatic treatment is commonly used to alleviate intestinal injury clinically. Actually, the main effective method to avoid bowel injury is to avoid intestine irradiation with better radiotherapy techniques (IMRT, IGRT, online adaptive RT). Radioprotective action such as use of radioprotectors, radiation mitigators, or therapeutic agents is usually undertaken during radiotherapy. A radioprotector as the earliest radiation countermeasure taken is usually given before radiation; thus, it is far superior to other agents in terms of therapeutic effect. Many natural and synthetic radioprotective agents have been applied for years, including ammonia and sulfur compounds, polyphenols, superoxide dismutase, cytokine, Chinese herbal medicine, etc. [
3]. However, there are still some problems need to be solved, such as toxicity, stability, effectivity, and administration route. The best known radioprotector, amifostine, a kind of thiol compound, is the only clinically approved synthetic radioprotector [
4]. However, amifostine may cause some side effects, including hypotension, nausea, vomiting, hot flashes, mild somnolence, and hypocalcemia, which limit its utility in clinical radiotherapy. To date, an effective and convenient radioprotective agent available to prevent or reduce radiation damage on the body has not been developed yet. It is necessary to explore an optimal radioprotector for individuals suffering from radiotherapy for tumors.
Melatonin (
N-acetyl-5-methoxytryptamine) is a neurohormone secreted by the pineal gland in the brain of mammals and humans. It was originally discovered to participate in the control of circadian rhythms and possibly sleep processes in diurnal species [
5]. Thereafter, melatonin was reported to be involved in the regulation of lots of biological processes, including anti-aging, antioxidation, scavenging free radical, regulation of the immune system, and prevention of tumors [
6]. Recent studies have indicated its effectiveness in resisting radiation in both in vitro and in vivo experiments [
7]. Melatonin can not only safely be self-administered orally, sublingually, or via an intranasal spray, but can also be used in the short term or long term with little to no significant side effects. It is worth noting that melatonin was successfully used on radiation accident victims and workers in the radiation leak from the Fukushima Daiichi Nuclear Power Plant. Therefore, Das et al. inferred that melatonin may be a desirable candidate as a countermeasure against IR exposure [
8].
To date, there have been a lot of reports about the potential of melatonin to protect against radiation. However, the literature on radiation-caused intestinal injury is limited. In this study, the effects of melatonin on intestinal injury caused by abdominal irradiation (ABI) from mice was first evaluated. Next, microarray analysis to investigate the mechanisms behind the protective effect of melatonin was performed. We explored the gene expression profiles mediated by melatonin following radiation. The microarray data were subjected to gene ontology (GO) annotation and ingenuity pathway analysis (IPA). Our data provided fundamental research clues to the radioprotective effects of melatonin and further in-depth study is necessary.
3. Discussion
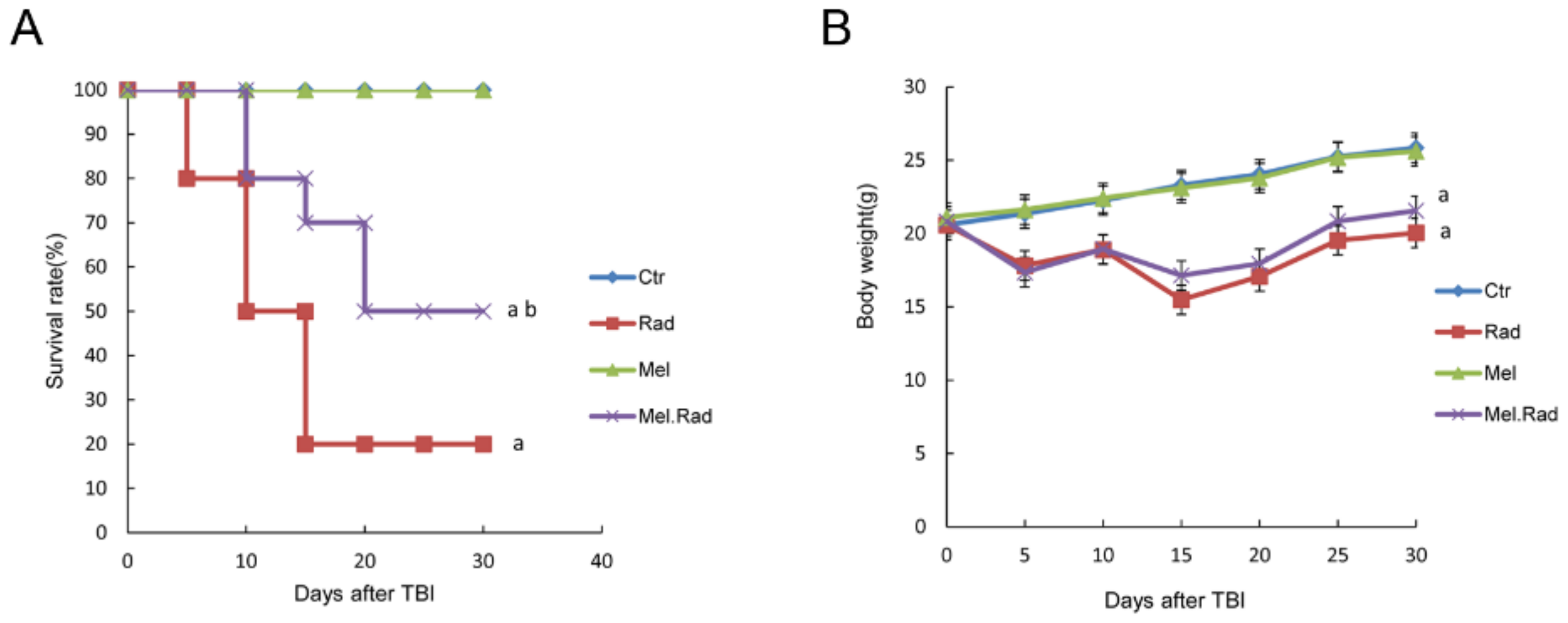
In the study, the possible effects of melatonin on radiation damage in mice were first investigated. Survival rate is regarded as an important parameter indicating radiation effects. We found that melatonin promoted the survival rate of mice irradiated with a fatal dose of TBI, suggesting the radioprotective effect of melatonin on the body.
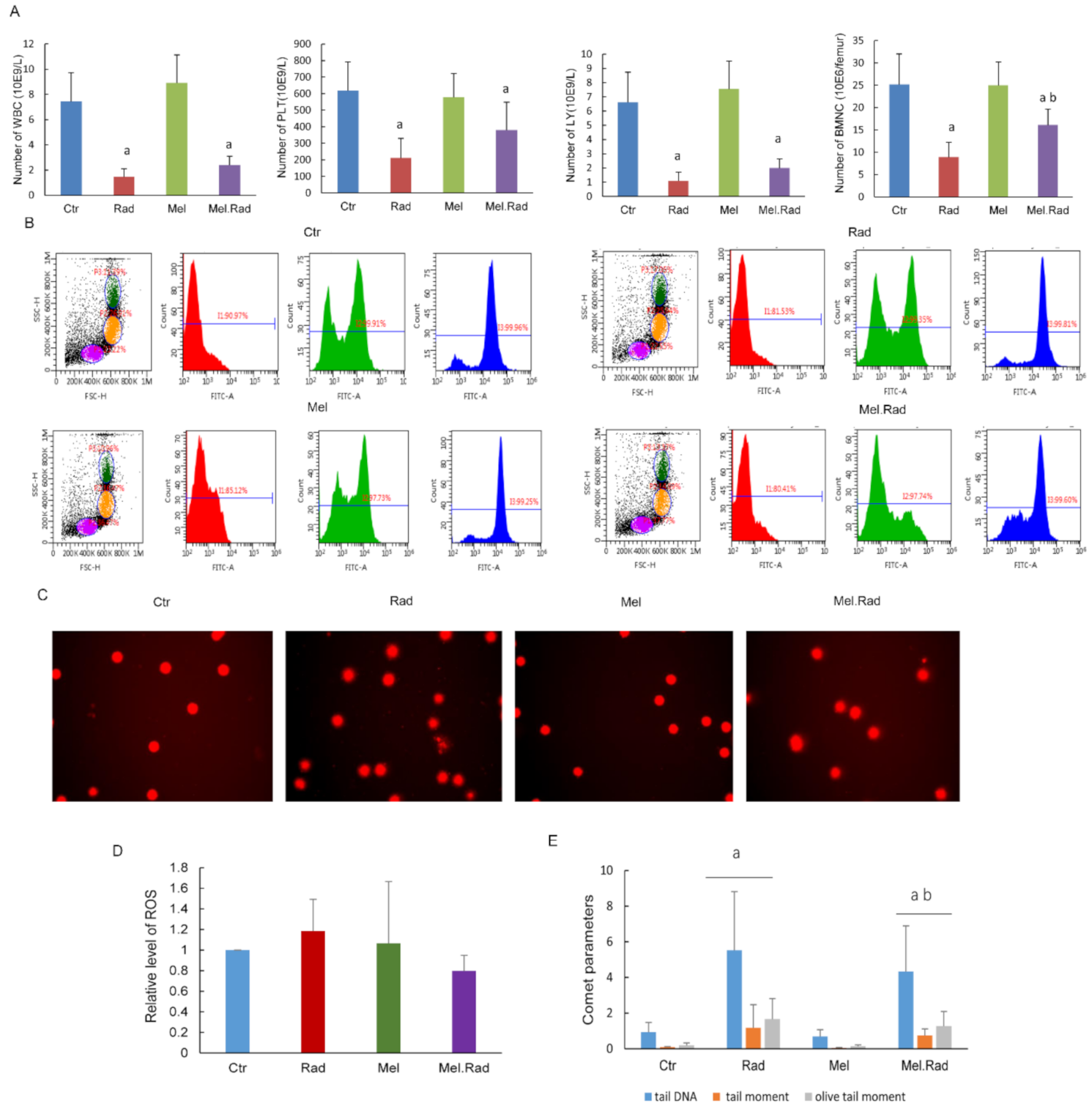
The hematopoietic system is extremely sensitive to radiation during radiotherapy, which may result in impaired hematopoietic function [
12]. In the study, melatonin treatment prior to radiation increased the counts of whole blood cells and bone marrow nucleated cells. DNA is the primary target for the lethal effects of radiation. ROS induced by radiation may destroy the structure and function of DNA, resulting in oxidative damage to cells [
13]. Because of its small size and high lipophilicity, melatonin can easily enter cellular membranes, reaching its highest concentration in the nucleus, and thus protecting the DNA of the nucleus from oxidative injury caused by IR [
14]. We found that melatonin decreased the generation of ROS in bone marrow and alleviated radiation-caused DNA damage of lymphocytes in whole blood. The data suggest that melatonin may alleviate radiation-caused damage on hematopoietic cells by promoting antioxidant activity. It has been well established that mice irradiated with fatal doses of radiation that survive depend on the restoration of the hematopoietic system [
15]. Therefore, the survival rate and the body weight of the melatonin-treated mice prior to radiation was improved in the study.
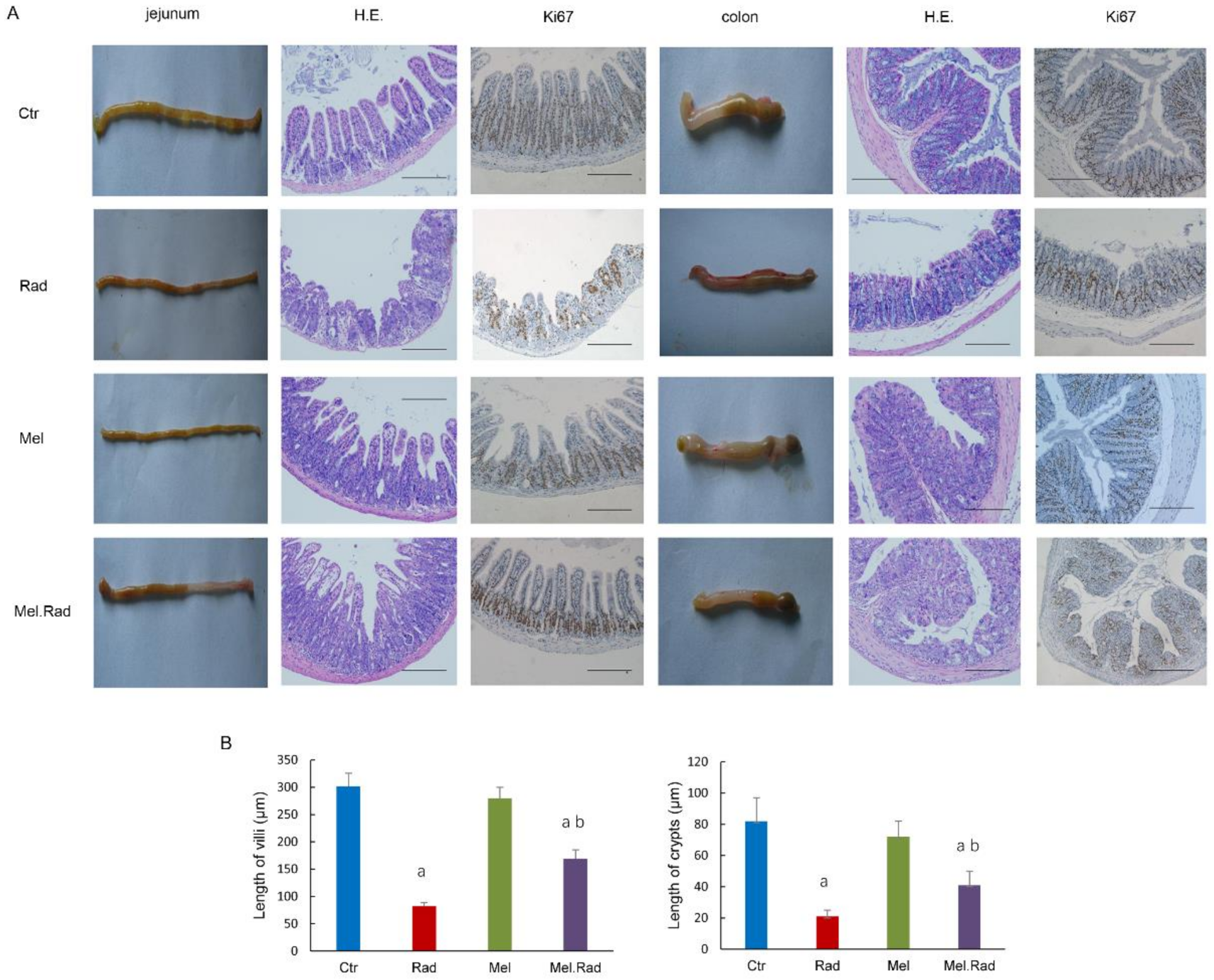
To disclose the effect of melatonin on radiation-caused intestinal injury, the intestines were evaluated by pathological histology. The results indicate that the impaired intestinal villus and severe loss of crypts caused by radiation were recovered by melatonin treatment. Thus, melatonin facilitated the recovery of absorption and barrier functions of the intestines and alleviated the intestinal injury caused by radiation.
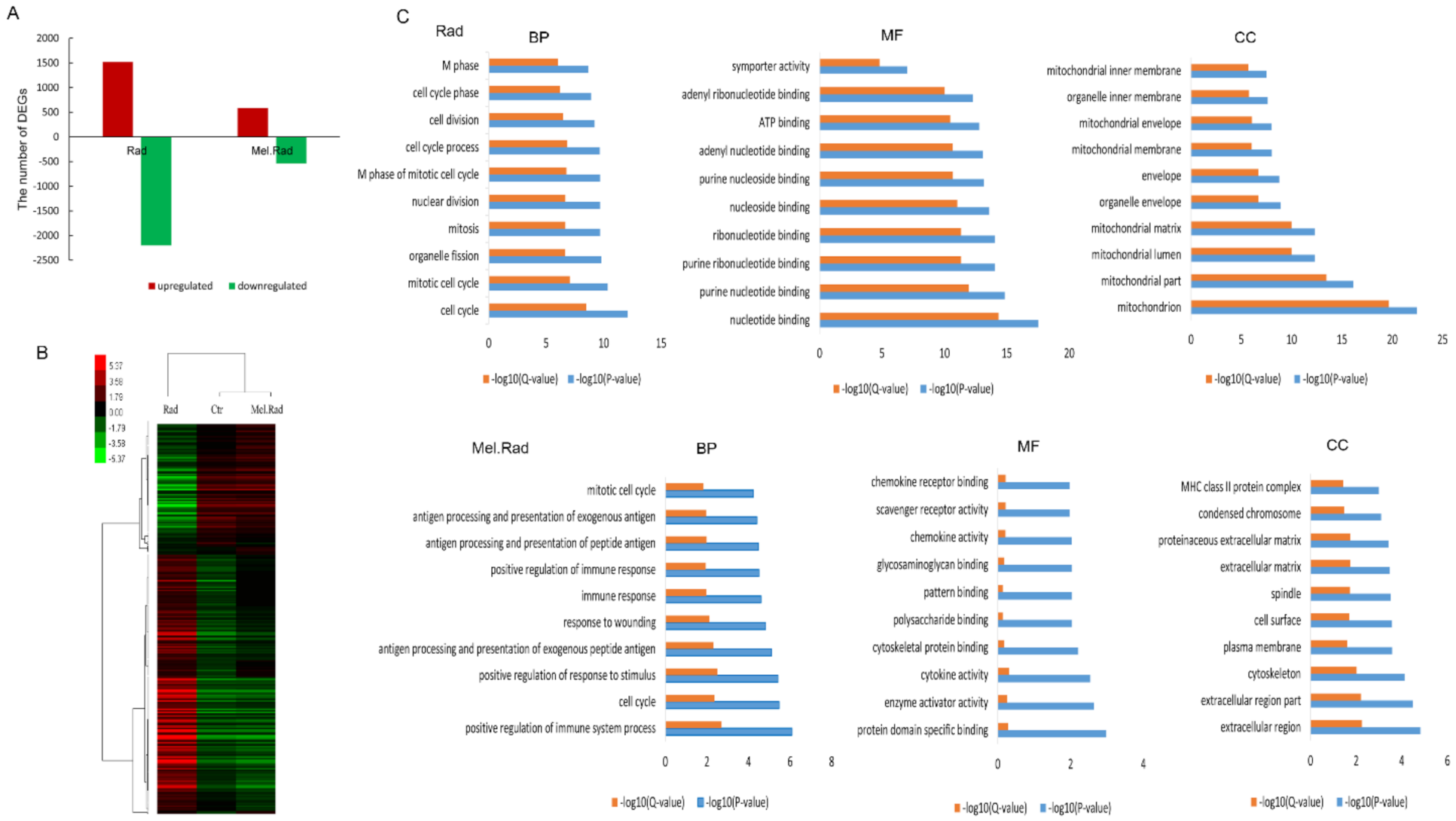
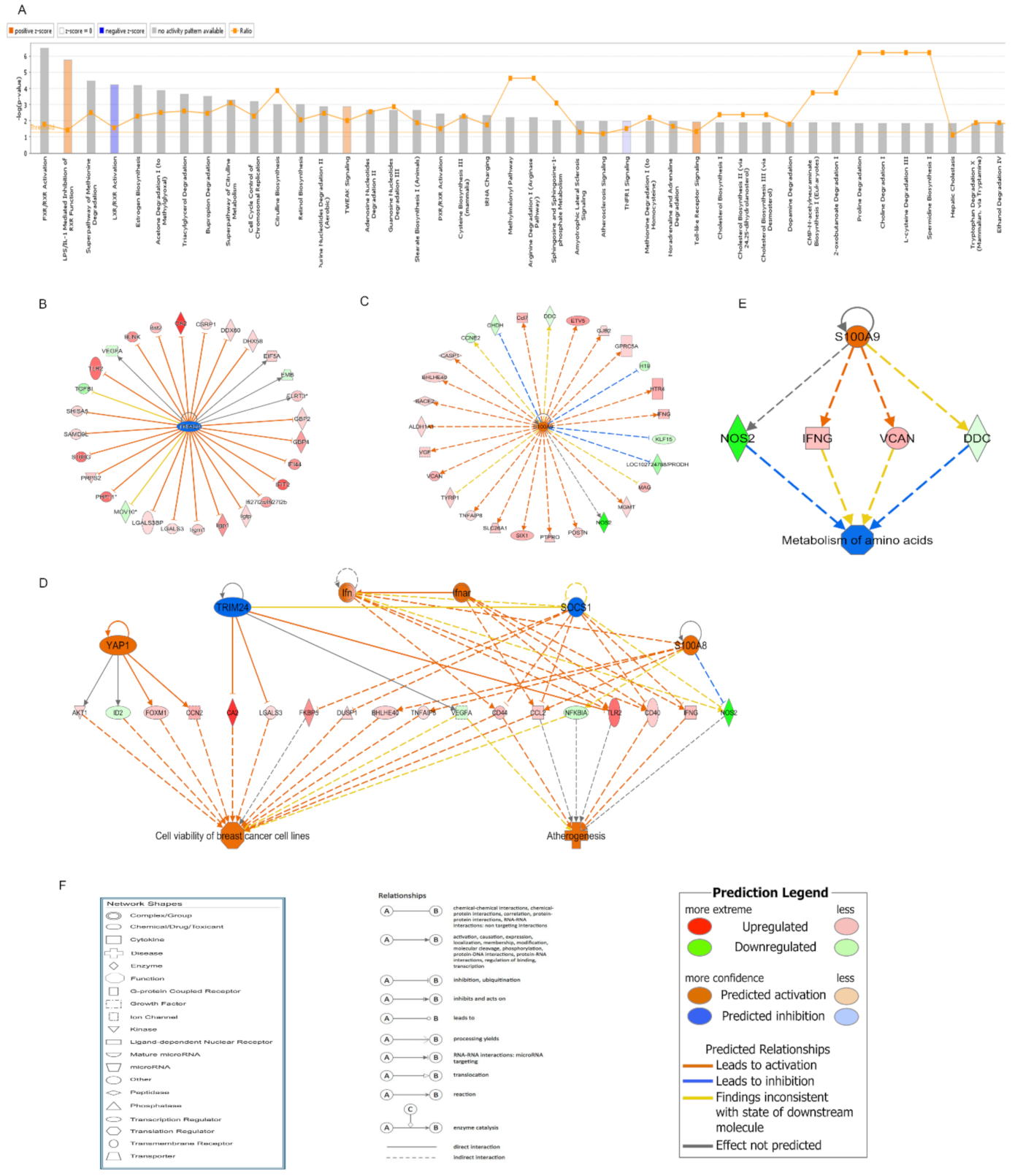
In exploring the mechanisms of the radioprotective function of melatonin on intestinal injury, melatonin treatment before radiation resulted in significant changes in DNA expression patterns of the small intestines. GO analysis clearly showed that the significantly enriched biological process mainly included the immune process in the mice treated with melatonin. Many studies have shown that melatonin has a positive effect on immunity [
16]. In the IPA analysis of canonical pathways, we found that the most activated pathways affected by melatonin treatment prior to radiation included LPS/IL-1-mediated inhibition of RXR function and Toll-like receptor signaling. Lipopolysaccharide (LPS), a principal component of the outer membrane of Gram-negative bacteria, strongly stimulates host innate immune response [
17]. Toll-like receptors (TLRs) are usually expressed in innate immune cells and non-immune cells such as epithelial cells and fibroblasts, and react to the membrane constituents of Gram-positive or Gram-negative bacteria [
18]. LPS recognition as a constituent of bacterial outer membrane by TLRs evokes rapid activation of innate immunity. The two activated pathways were involved in innate immune responses, suggesting that the activation of innate immunity plays an important role in the protective effect of melatonin against IR.
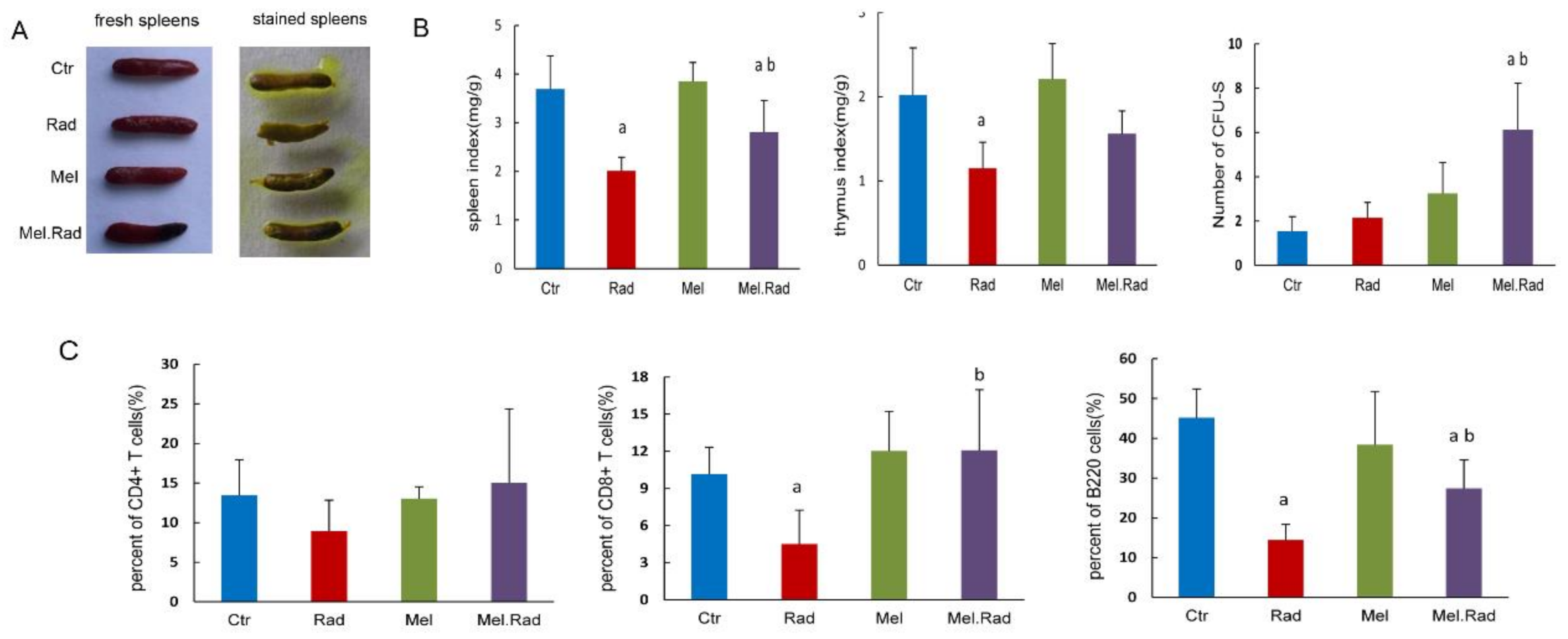
To determine the effect of melatonin on innate immunity, we examined the change in immune organs and immune cells following melatonin treatment. The spleen is the largest lymphoid organ in the human body and the colony-forming ability of the spleen was used to evaluate the immune function following exposure to high doses of IR [
15]. We found that melatonin administration resulted in an increase in the CFU-S and spleen/thymus coefficient. A similar effect of melatonin treatment on CD4
+ T-cell, CD8
+ T-cell, and B-cell recovery in blood was also observed in the irradiated mice. The data indicated that melatonin treatment motivated the recovery of immunity following IR and immune regulation was related to the protective effect of melatonin on intestinal injury. It was reported that a significantly enriched biological process was involved in innate and adaptive immunity in male mice treated with sex-matched fecal microbiota transplantation to alleviate intestinal injury [
19], which was consistent with our results.
In further analysis of the upstream regulatory network and regulator effects network affected by melatonin treatment before radiation, TRIM24 was forecasted to be the most significantly inhibited upstream regulator. TRIM24 is a member of the tripartite motif family, which plays an important role in regulation of innate immunity, autophagy, and carcinogenesis [
20]. TRIM24 was reported to regulate the activity of the retinoic acid receptor and the tumor suppressor p53 [
21]. Diseases such as differentiated thyroid carcinoma, hepatocellular carcinomas, and cerebellar agenesis were associated with TRIM24. In the study, the regulatory network of TRIM24 activated atherogenesis and cell viability in breast cancer cell lines. In addition, we found that S100A9 was forecasted to be the most significantly activated upstream regulator. S100A9 is a calcium- and zinc-binding protein. Usually, S100A9 and S100A8 are secreted in a heterodimeric form by activating immunocytes such as monocytes, granulocytes, and neutrophils [
22], which are involved in the regulation of inflammatory processes and immune response [
23]. In the study, the regulatory network of S100A9 inhibited the metabolism of amino acids.
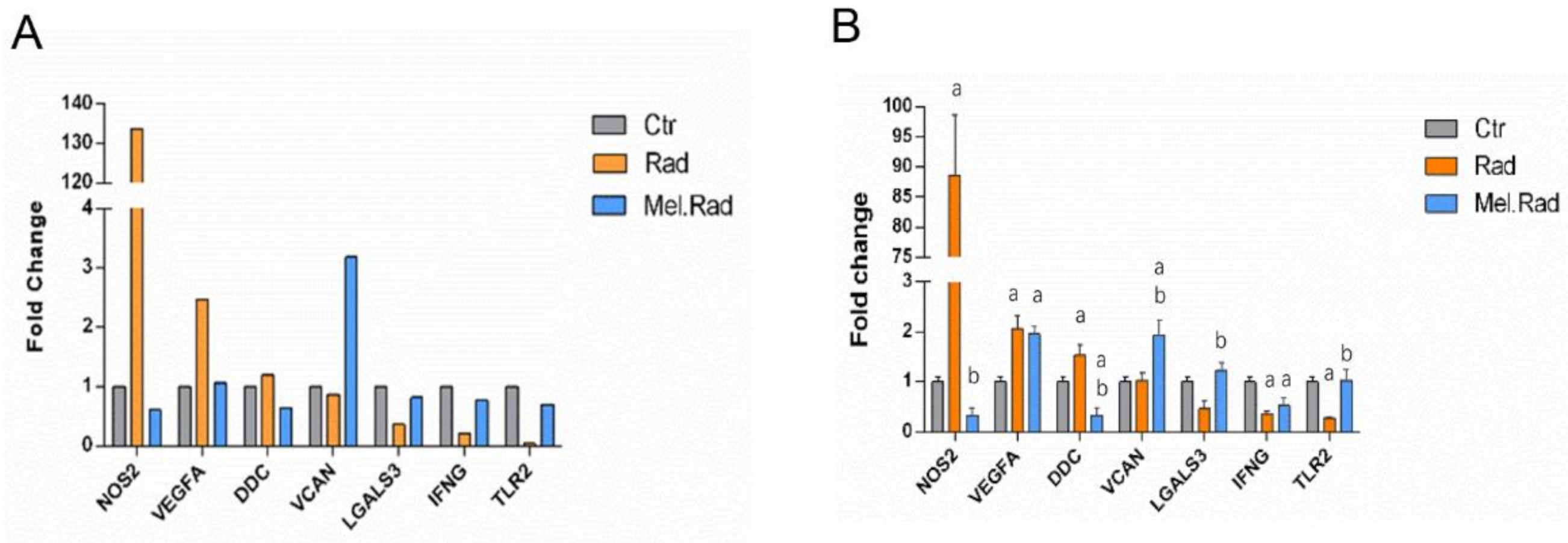
In the qRT-PCR validation of the candidate DEGs involved in the regulatory network of TRIM24 and S100A9, there was an obvious increase and decrease in the mRNA levels of TLR2 and NOS2, respectively, in the small intestines of mice treated with melatonin. TLR2 is expressed on many cells such as macrophages, dendritic cells, and monocytes that are responsive to peptides from bacteria. TLR2 participated in the two signaling pathways, Toll-like receptor signaling and LPS-mediated signaling [
24]. The increased TLR2 level in the Mel.Rad mice denoted that immune response was activated following melatonin treatment. NOS2, a nitric oxide synthase, resulted in inflammation by increasing the synthesis of proinflammatory mediators such as IL6 and IL8 [
25]. The decreased NOS2 level in the Mel.Rad mice suggested that inflammation was inhibited by melatonin’s immune defense.
We preliminarily explored the mechanisms of the radioprotective effect of melatonin on intestinal injury, which were associated with activation of the immune response. However, the relationship between the two activated pathways involved in immunity, LPS/IL-1-mediated inhibition of RXR function, and Toll-like receptor signaling was not investigated in this study. So far, the roles of TRIM24 and S100A9 in the regulatory network associated with melatonin treatment following radiation have not been reported. How TRIM24 and S100A9 participate in the beneficial effect of melatonin are still not very clear. It is required to conduct in-depth experiments to facilitate the understanding of melatonin as a radioprotector for radiation enteropathy.
Studies on healthy male volunteers by oral administration of melatonin in doses of 1–300 mg [
26] or 1 g of melatonin daily for 30 days [
27] reported no evidence of toxicity. In addition, some randomized clinical trials indicated that administration of melatonin (20 mg/day) improved the quality of life of cancer patients and thus demonstrated a therapeutic role of melatonin [
28,
29]. Melatonin administration, either alone or combined with radiotherapy, resulted in a favorable outcome. Melatonin has the potential as a radioprotector in the radiotherapy for tumor patients.
In conclusion, the data provided a scene of gene expression of melatonin treatment before radiation and fundamental research clues to the protective effect of melatonin. Melatonin might promote immune response and thus protect the intestines from radiation damage. However, the associated mechanisms of activation of the immunity have not yet been elucidated thoroughly. It is necessary to conduct further intensive experiments to explore the underlying mechanisms of melatonin’s immune regulation on intestinal injury.
4. Materials and Methods
4.1. Mice
Male C57BL/6J mice were obtained from Vital River (Beijing, China). Mice 6–8 weeks of age and with 18 ± 2 g of body weight were used. The mice were raised under laboratory conditions of 23 ± 2 °C temperature, 50 ± 5% humidity, and light/dark 10/14 h. The mice were kept in the experimental animal center at the Institute of Radiation Medicine of the Chinese Academy of Medical Science. The mice were humanely euthanized through 100% CO2 inhalation, in accordance with the principles guidelines of the Institutional Animal Care and Ethics Committee guidelines.
4.2. Radiation Exposure in Mice
The mice were anaesthetized with chloral hydrate solution (0.3 g/kg body weight) by intraperitoneal injection. The mice were restrained in a perforated wooden container and radiation was delivered to the abdominal area with the other parts of the body covered by lead plate. The mice were irradiated with a 137Cesium γ irradiator (Atomic Energy of Canada Inc., Chalk River, ON, Canada) at a rate of 0.883 Gy/min.
4.3. Administration of Melatonin
Melatonin (
N-acetyl-5-methoxytryptamine) was purchased from Sigma-Aldrich (St. Louis, MO, USA). The melatonin was dissolved in 50 μL absolute ethanol and diluted with isotonic NaCl solution to a final ethanol concentration of 10%. The mice were treated with 10 mg/kg body weight of melatonin by intraperitoneal injection. The dose of melatonin was selected based upon our previous study [
30].
4.4. Experimental Design
The animals were randomly split up into four groups: (i) Ctrl group, mice received no treatment. (ii) Rad group, mice were irradiated with a certain dose of gamma irradiation once. (iii) Mel group, mice were administered with melatonin by intraperitoneal injection. (iv) Mel.Rad group, mice were administered melatonin by intraperitoneal injection. One hour following the last administration, the mice were irradiated with a certain dose of gamma irradiation once. Throughout the experiment, melatonin was administered between 4 p.m. and 5 p.m. Melatonin is regarded as being at its lowest natural concentration in the blood during this time period.
4.5. Analysis of 30-Day Survival Rate
The mice were treated based on the grouping method. The mice were observed for up to 30 days following a fatal dose of 7.2 Gy TBI. The number of surviving animals and the body weight of the surviving animals was examined. The survival curve of the animals was drawn by the Kaplan–Meier method.
4.6. Pathological Analysis
The intestines of the mice were dissected and immersed in 4% neutral buffered formalin for 24 h. The paraffin-embedded sections were sliced and dyed with hematoxylin and eosin staining. The slides were observed with a microscope (BX51, Olympus, Shinjuku, Tokyo, Japan).
4.7. Immunohistochemical Analysis
Paraffin-embedded slides were dewaxed and rehydrated with xylene and ethanol, respectively, and put in 3% hydrogen peroxide solution for 10 min to block endogenous peroxidase. The slides were immersed in Tris/EDTA (pH 9.0) and incubated for 15 min at 98 °C to retrieve antigens. The slides were incubated with serum for 30 min at room temperature to block nonspecific antigen binding sites. Mouse monoclonal anti-Ki67 antibody was diluted (Abcam Biotechnology, Cambridge, UK, No. ab8191, 1:1000 dilution), added to the slides, and incubated overnight at 4 °C. Secondary antibody HRP goat anti-mouse (IgG) antibody (Abcam Biotechnology, Cambridge, UK, No. ab205719) was added and incubated at 37 °C for 2 h.
4.8. Peripheral Blood Cell and Bone-Marrow Cell Counts
A volume of 0.2 mL of whole blood was drawn from the orbital sinuses of mice by a micropipette coated with an anticoagulant. The cells, including white blood cells (WBC), platelets (PLT), and lymphocytes (LY), were counted with the hemocytometer (Sysmex pocH-100i; Sysmex Corporation, Kobe, Japan). Bone marrow nucleated cells (BMNCs) were collected from the femurs as per previous literature and counted [
31].
4.9. Reactive Oxygen Species (ROS) Level Assay
BMNCs were collected from the femurs. The levels of ROS were detected by ROS assay kit 520 nm (Invitrogen, Carlsbad, CA, USA) using a flow cytometer. A 1 × ROS assay stain solution (100 μL) was added to each sample and incubated for 60 min at 37 °C. Then the solution was washed twice with PBS solution and re-suspended in 500 μL PBS solution. A minimum of 10,000 cells for each sample were collected and analyzed using FlowJo 7.6.1 software (Tree-Star, Ashland, OR, USA).
4.10. Analysis of DNA Damage in Lymphocytes and Lymphocyte Phenotype
A volume of 0.2 mL of whole blood was drawn from the orbital sinuses of mice. Lymphocytes were isolated with the lymphocyte separation medium and a comet assay was carried out in alkaline conditions as described previously [
32]. The slides were observed using an ECLIPSE 90i fluorescence microscope (Nikon, Tokyo, Japan). A total of 100 comet images were collected for each sample with a digital imaging system (Cherry Hill, NJ, USA) and analyzed using the Comet Assay Software Project (CASP, Wroclaw, Poland) [
33]. The proportion of DNA in the tail DNA (TDNA), tail moment (TM), and olive tail moment (OTM) were calculated to record the degree of DNA damage. A lymphocyte phenotype assay was carried out by flow cytometry as described previously [
34].
4.11. cDNA Microarray Analysis
The small intestines of mice were harvested and immediately immersed in RNA stabilization reagent (Qiegen, Shanghai, China). Total RNA was drawn with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and reverse transcribed to complementary DNA using the Quantscript RT kit (Tiangen Biotech, Beijing, China). The aminoallyl-RNA probes were labelled with Cy5 hybridized for 16 h at 50 °C to a mouse oligo microarray (Mouse Whole Genome OneArray MOA2.1; Phalanx Biotech Group, Hsinchu, Taiwan). All arrays were scanned using Agilent Microarray Scanner G2505C at a 10 μm resolution. Raw data were obtained from the scanned images and analyzed with Rosetta Resolver 7.2 software (Rosetta Biosoftware Technologies, San Diego, CA, USA). The PLIER default protocol was used to normalize the data.
4.12. GO Annotation and IPA Analysis
DEGs with at least 2-fold change in expression level with a p-value (differentially expressed) less than 0.05 were regarded to be significantly upregulated or downregulated. To gain insight into the levels of DEGs identified by cDNA microarray, GO annotation analysis was performed, including CC, MF, and BP. The GO terms were considered to be significantly enriched when the p-value (differentially expressed) was less than 0.05. Microarray data were analyzed by IPA (Ingenuity Systems, Redwood City, CA, USA) to ascertain the canonical pathways, upstream regulation, and regulator effect based on the whole DEGs involved in and previous literature.
4.13. qRT-PCR
Total RNA was isolated from the small intestines with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and reverse transcribed to complementary DNA was synthesized with an RNA PCR kit. The qPCR experiment was conducted using a SYBR Green kit (Takara Bio Inc., Dalian, China). The primer sequences were as follows: LGALS3: 5′-AGACAGCTTTTCGCTTAACGA-3′ (forward) and 5′-GGGTAGGCACTAGGAGGAGC-3′ (reverse), TLR2: 5′-GCAAACGCTGTTCTGCTCAG-3′ (forward) and 5′-AGGCGTCTCCCTCTATTGTATT-3′ (reverse), VEGFA: 5′-GAGGTCAAGGCTTTTGAAGGC-3′ (forward) and 5′-CTGTCCTGGTATTGAGGGTGG-3′ (reverse), DDC: 5′-TAGCTGACTATCTGGATGGCAT-3′ (forward) and 5′-GTCCTCGTATGTTTCTGGCTC-3′ (reverse), IFNG: 5′-TCCTCGCCAGACTCGTTTTC-3′ (forward) and 5′-GTCTTGGGTCATTGCTGGAAG-3′ (reverse), NOS2: 5′-GTTCTCAGCCCAACAATACAAGA-3′ (forward) and 5′-GTGGACGGGTCGATGTCAC-3′(reverse), VCAN: 5′-TTTTACCCGAGTTACCAGACTCA-3′ (forward) and 5′-GGAGTAGTTGTTACATCCGTTGC-3′ (reverse), and GAPDH: 5′-AGGTCGGTGTGAACGGATTTG-3′ (forward) and 5′-TGTAGACCATGTAGTTGAGGTCA-3′ (reverse). The primers were synthesized by Sangon Biotech (Shanghai, China).
4.14. Calculation of CFU-S and Spleen/Thymus Coefficient
The spleens and thymuses were dissected out from the abdominal cavity after the mice were euthanized. The spleens were stained using picric acid for 24 h and observed under a macroscope to count CFU-S. One nodule on the spleen surface that was visible to the naked eye was considered one CFU-S [
35]. The spleen/thymus coefficient was obtained by dividing the weight of a single spleen/thymus (mg) by the body weight (g).
4.15. Statistical Analysis
Each experiment was performed at least three times. Data are shown as mean ± SD. The 30-day survival rate was compared using the Kaplan–Meier method with a log-rank test. Statistical differences between groups were analyzed by one-way ANOVA followed by the Bonferroni post hoc test. A p-value less than 0.05 was considered to be statistically significant.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






