
Integrative Role of 14-3-3ε in Sleep Regulation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Stocks
2.2. Behavioral Assays
2.3. Immunofluorescence
2.4. Total RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR
2.5. Western Blot Analysis and Co-Immunoprecipitation
2.6. Chromatin Immunoprecipitation Assay
2.7. Calculation of Axonal Cross
2.8. Proteomic Screen and RNA-Seq
2.9. Statistical Analysis
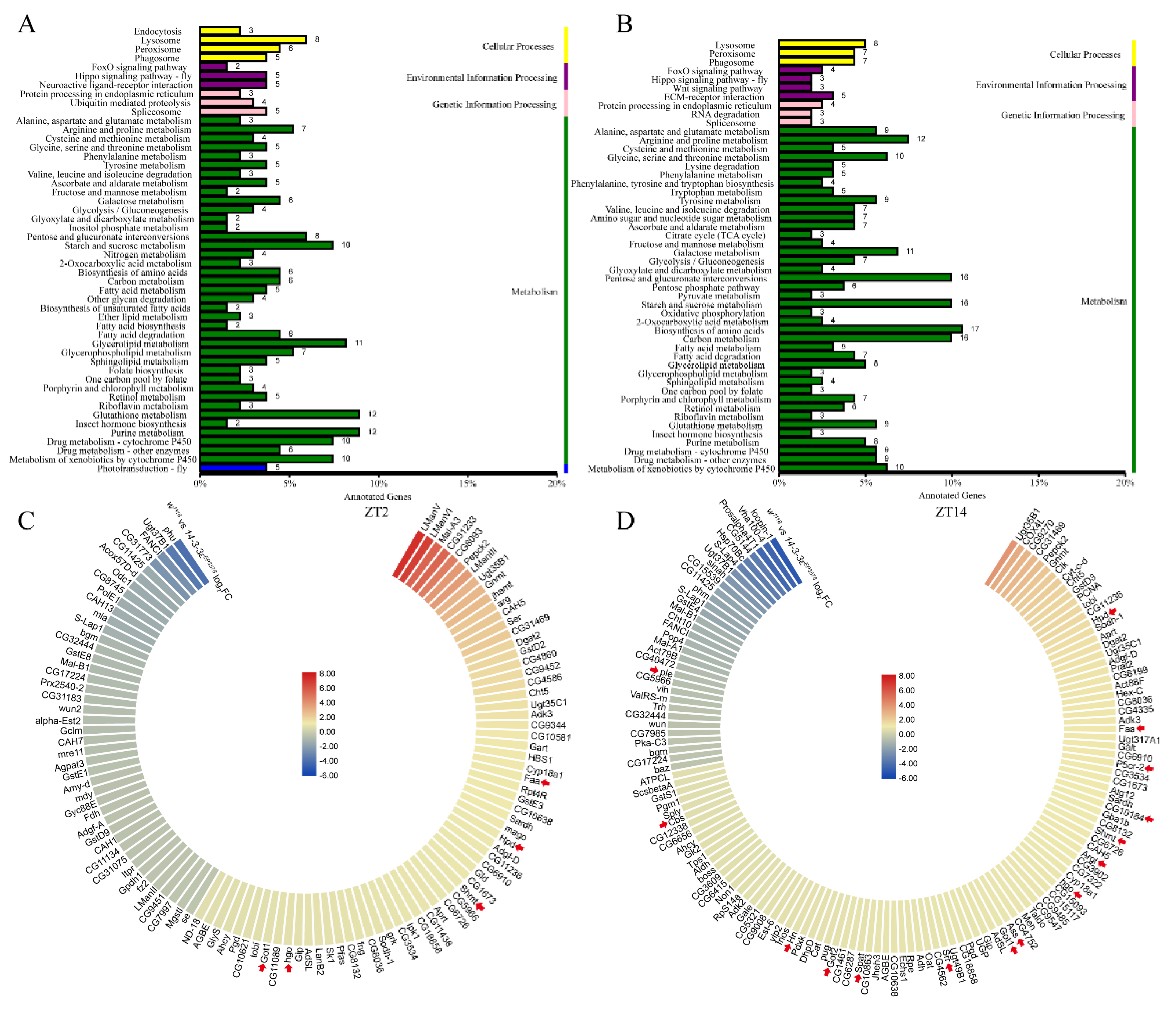
3. Results
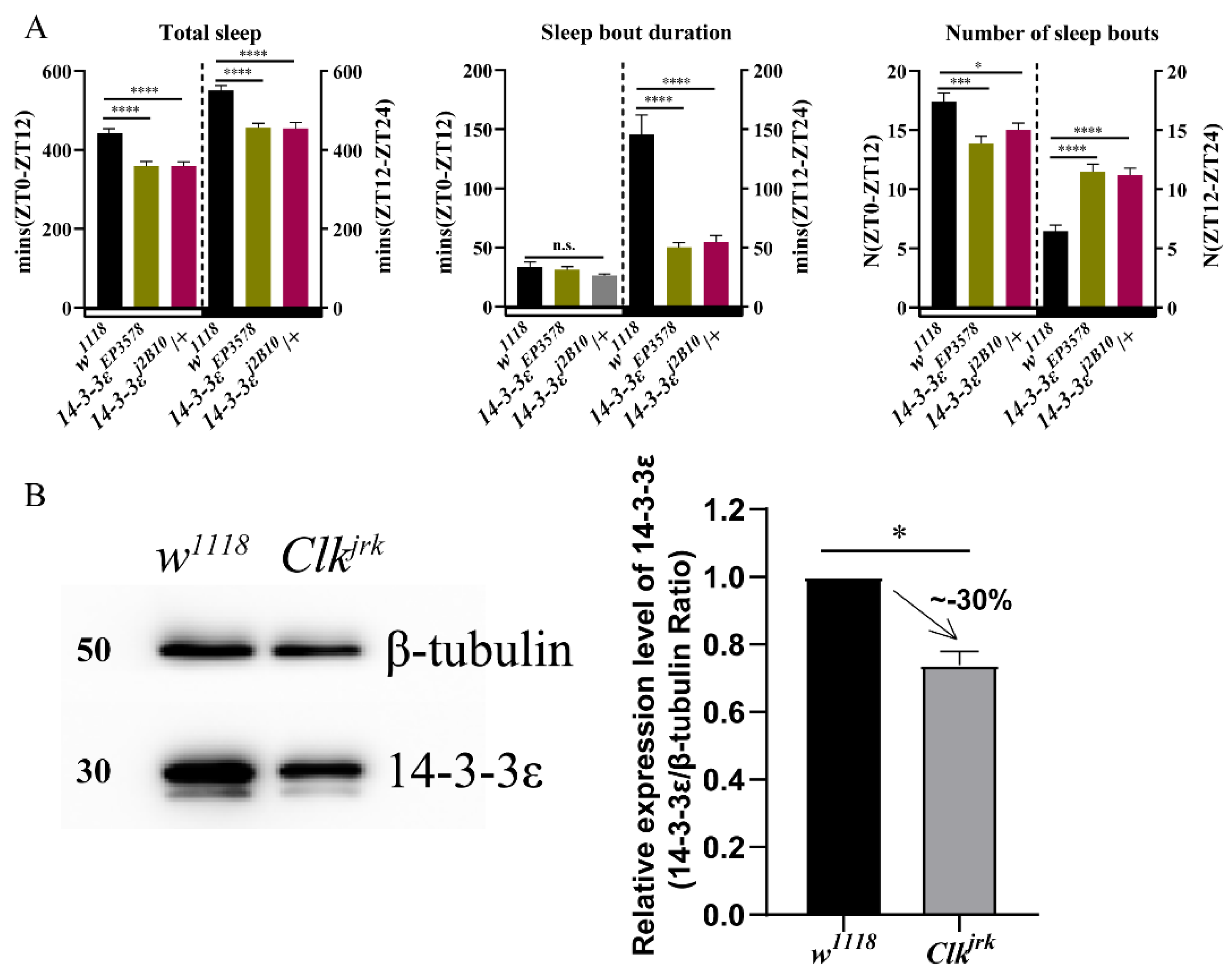
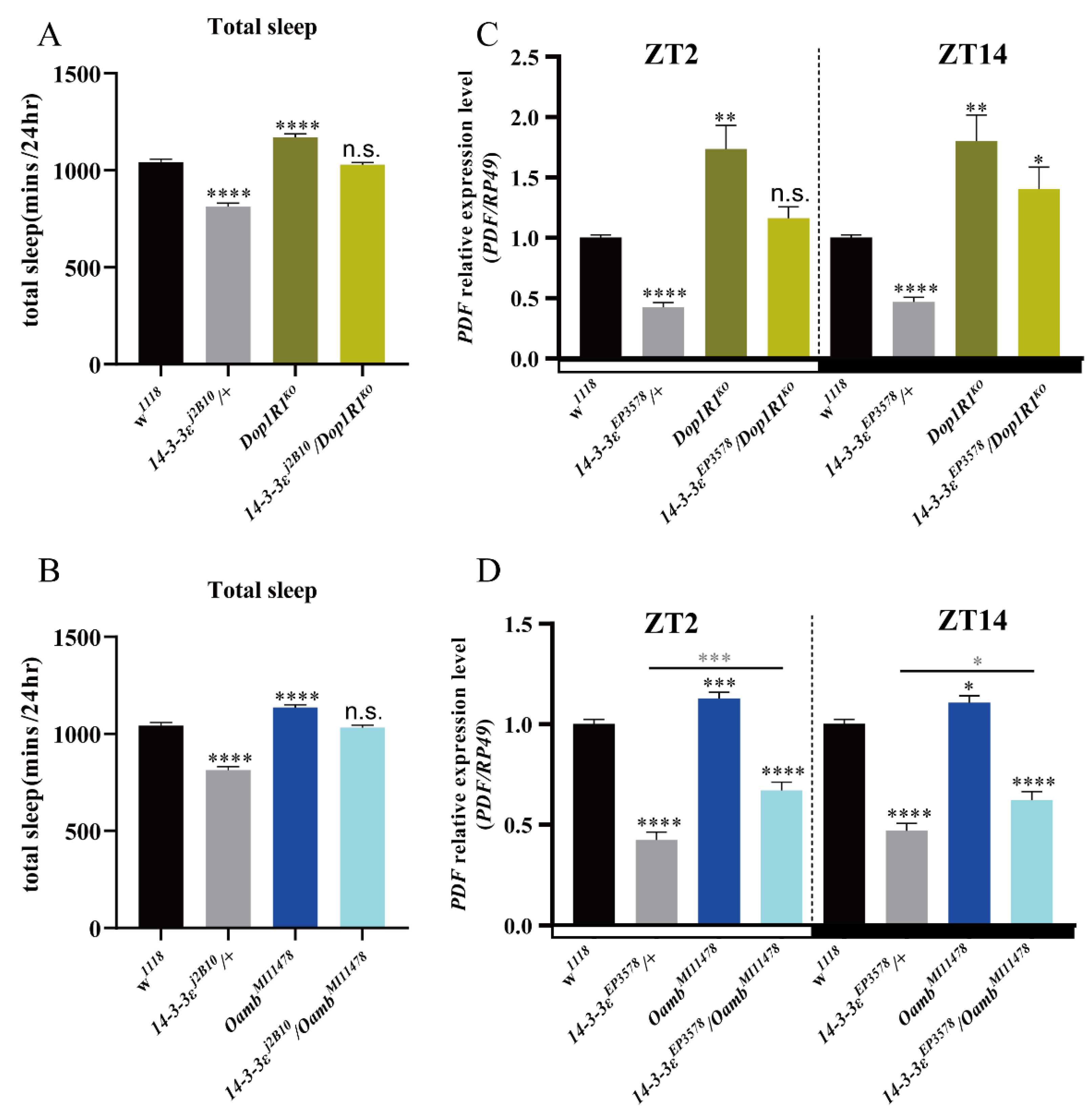
3.1. 14-3-3ε Regulates Sleep Independent of the Circadian Rhythm
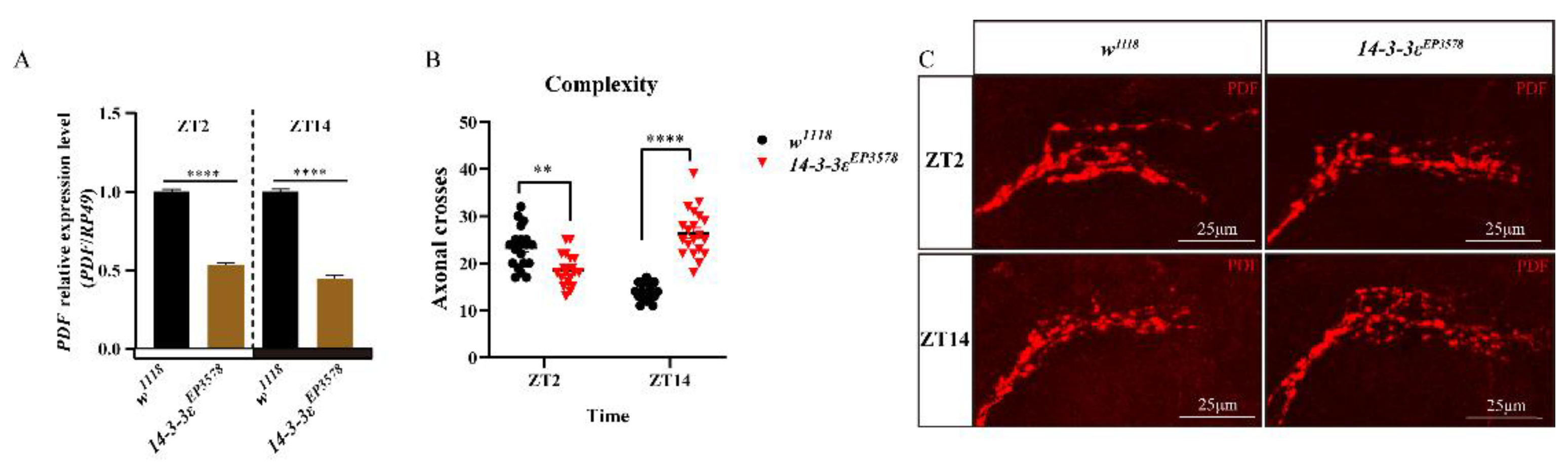
3.2. 14-3-3ε Regulates Sleep Factor PDF
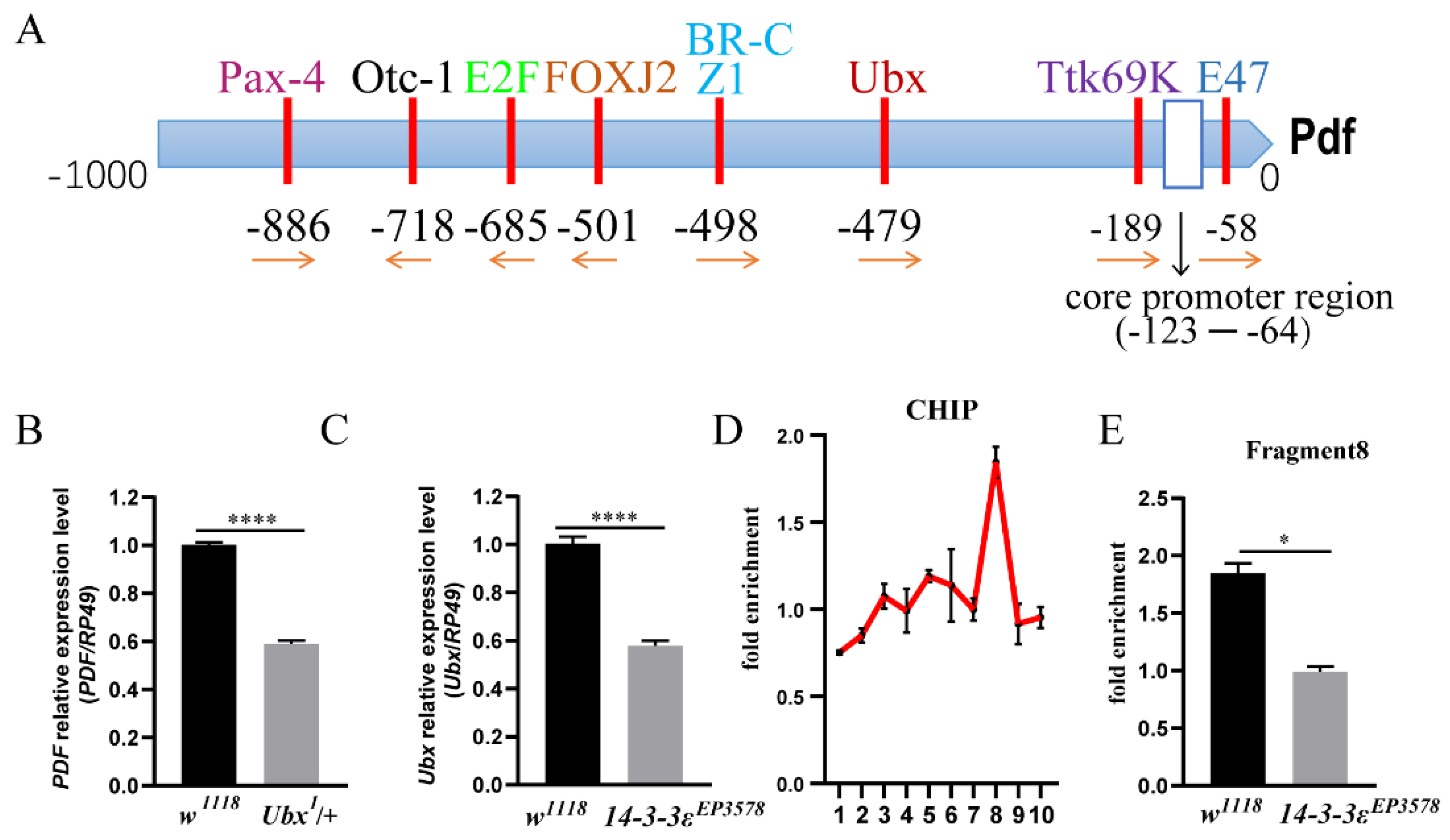
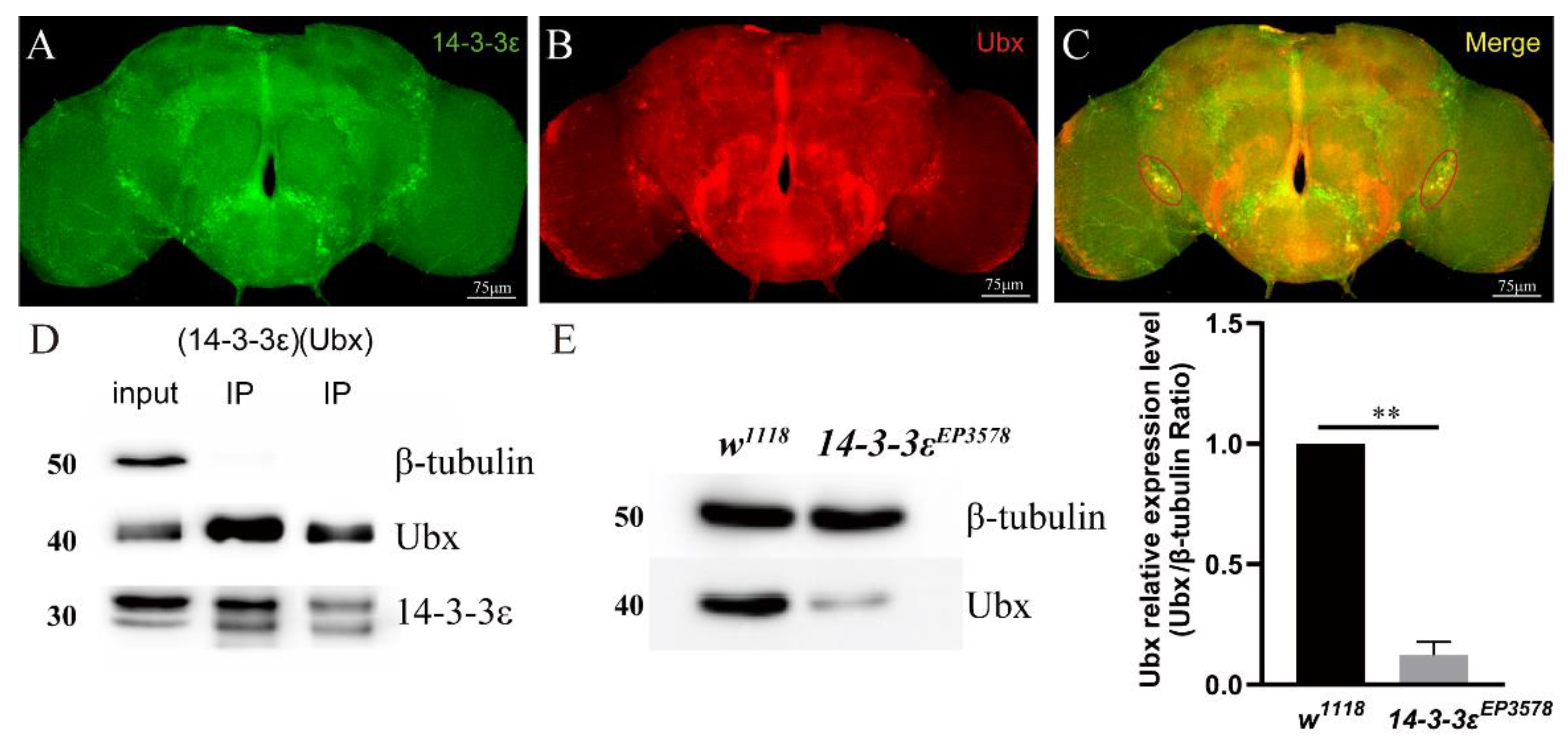
3.3. 14-3-3ε Directly Acts on Ubx to Regulate PDF Transcription
3.4. 14-3-3ε Regulates Sleep by Impacting Neurotransmitters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deboer, T. Sleep homeostasis and the circadian clock: Do the circadian pacemaker and the sleep homeostat influence each other’s functioning? Neurobiol. Sleep Circadian Rhythms 2018, 5, 68–77. [Google Scholar] [CrossRef]
- Franken, P.; Dijk, D.-J. Circadian clock genes and sleep homeostasis. Eur. J. Neurosci. 2009, 29, 1820–1829. [Google Scholar] [CrossRef]
- Szmyd, B.; Rogut, M.; Białasiewicz, P.; Gabryelska, A. The impact of glucocorticoids and statins on sleep quality. Sleep Med. Rev. 2020, 55, 101380. [Google Scholar] [CrossRef]
- Allada, R.; Siegel, J.M. Unearthing the Phylogenetic Roots of Sleep. Curr. Biol. 2008, 18, R670–R679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendricks, J.C.; Finn, S.M.; Panckeri, K.A.; Chavkin, J.; Williams, J.A.; Sehgal, A.; Pack, A. Rest in Drosophila Is a Sleep-like State. Neuron 2000, 25, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Shaw, P.J.; Cirelli, C.; Greenspan, R.J.; Tononi, G. Correlates of Sleep and Waking in Drosophila melanogaster. Science 2000, 287, 1834–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, B.Y.; Kilman, V.L.; Keath, J.R.; Pitman, J.L.; Allada, R. The GABA(A) receptor RDL acts in peptidergic PDF neurons to promote sleep in Drosophila. Curr. Biol. 2009, 19, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Cerullo, I.; Chen, X.; Rosbash, M. PDF neuron firing phase-shifts key circadian activity neurons in Drosophila. Elife 2014, 3, e02780. [Google Scholar] [CrossRef] [PubMed]
- Heekeren, H.; Marrett, S.; Bandettini, P.A.; Ungerleider, L.G. A general mechanism for perceptual decision-making in the human brain. Nature 2004, 431, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Shafer, O.T. The Drosophila Circadian Clock Is a Variably Coupled Network of Multiple Peptidergic Units. Science 2014, 343, 1516–1520. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Yu, J.; Jung, H.J.; Abruzzi, K.C.; Luo, W.; Griffith, L.C.; Rosbash, M. Circadian neuron feedback controls the Dro-sophila sleep-activity profile. Nature 2016, 536, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.D.L.P.; Berni, J.; Ceriani, M.F. Circadian Remodeling of Neuronal Circuits Involved in Rhythmic Behavior. PLoS Biol. 2008, 6, e69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Du, J.; Wei, L.; Zhao, Z. AKH-FOXO pathway regulates starvation-induced sleep loss through remodeling of the small ventral lateral neuron dorsal projections. PLoS Genet. 2020, 16, e1009181. [Google Scholar] [CrossRef]
- Nian, X.; Chen, W.; Bai, W.; Zhao, Z.; Zhang, Y. miR-263b Controls Circadian Behavior and the Structural Plasticity of Pacemaker Neurons by Regulating the LIM-Only Protein Beadex. Cells 2019, 8, 923. [Google Scholar] [CrossRef] [Green Version]
- Morrison, D.K. The 14–3-3 proteins: Integrators of diverse signaling cues that impact cell fate and cancer development. Trends Cell Biol. 2009, 19, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14–3-3 protein family in health, disease, and drug development. Drug Discov. Today 2016, 21, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Pan, D. The Hippo Signaling Pathway in Development and Cancer. Dev. Cell 2010, 19, 491–505. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Li, L.; Lei, Q.-Y.; Guan, K.-L. The Hippo-YAP pathway in organ size control and tumorigenesis: An updated version. Genes. Dev. 2010, 24, 862–874. [Google Scholar] [CrossRef] [Green Version]
- Ren, F.; Zhang, L.; Jiang, J. Hippo signaling regulates Yorkie nuclear localization and activity through 14–3-3 dependent and independent mechanisms. Dev. Biol. 2010, 337, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, T.; Isobe, T.; Okuyama, T.; Yamauchi, T.; Fujisawa, H. Brain 14–3-3 Protein Is an Activator Protein That Activates Tryptophan 5-Monooxygenase and Tyrosine 3-Monooxygenase in the Presence of Ca-2+, Calmodulin-Dependent Protein Ki-nase-Ii. FEBS Lett. 1987, 219, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, Y. 14–3-3 Proteins in Glutamatergic Synapses. Neural Plast. 2018, 2018, 1–6. [Google Scholar] [CrossRef]
- Deng, B.; Li, Q.; Liu, X.; Cao, Y.; Li, B.; Qian, Y.; Xu, R.; Mao, R.; Zhou, E.; Zhang, W.; et al. Chemoconnectomics: Mapping Chemical Transmission in Drosophila. Neuron 2019, 101, 876–893.e4. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Shi, W.; Li, L.; Zheng, Z.; Li, T.; Bai, W.; Zhao, Z. Regulation of sleep by the short neuropeptide F (sNPF) in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2013, 43, 809–819. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, F.; Liu, J.; Wang, D.; Yan, Y.; Ji, S.; Zan, J.; Zhou, J. The co-chaperone Cdc37 regulates the rabies virus phospho-protein stability by targeting to Hsp90AA1 machinery. Sci. Rep. 2016, 6, 27123. [Google Scholar] [CrossRef]
- Liu, X.; Li, H.; Liu, Q.; Niu, Y.; Hu, Q.; Deng, H.; Cha, J.; Wang, Y.; Liu, Y.; He, Q. Role for Protein Kinase A in the Neurospora Circadian Clock by Regulating White Collar-Independent frequency Transcription through Phosphorylation of RCM-1. Mol. Cell. Biol. 2015, 35, 2088–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Zhang, Y.F.; Xue, Y.B.; Zhao, X.Y.; Zhao, X.G.; Wei, Y.; Li, Z.; Zhang, Y.; Zhao, Z.W. Diurnal protein oscillation profiles in Drosophila head. FEBS Lett. 2018, 592, 3736–3749. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Haynes, P.; Pírez, N.; Harrington, K.; Guo, F.; Pollack, J.; Hong, P.; Griffith, L.; Rosbash, M. Imaging analysis of clock neurons reveals light buffers the wake-promoting effect of dopamine. Nat. Neurosci. 2011, 14, 889–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Griffith, L.; Rosbash, M. Light-arousal and circadian photoreception circuits intersect at the large PDF cells of the Drosophila brain. Proc. Natl. Acad. Sci. USA 2008, 105, 19587–19594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisky, K.M.; Agosto, J.; Pulver, S.R.; Shang, Y.; Kuklin, E.; Hodge, J.J.; Kang, K.; Liu, X.; Garrity, P.A.; Rosbash, M.; et al. PDF cells are a GABA-responsive wake-promoting component of the Drosophila sleep circuit. Neuron 2008, 60, 672–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Mignot, E. Genetics of Sleep and Sleep Disorders. Cell 2011, 146, 194–207. [Google Scholar] [CrossRef] [Green Version]
- Mochizuki, T.; Crocker, A.; McCormack, S.; Yanagisawa, M.; Sakurai, T.; Scammell, T.E. Behavioral State Instability in Orexin Knock-Out Mice. J. Neurosci. 2004, 24, 6291–6300. [Google Scholar] [CrossRef]
- Aton, S.J.; Colwell, C.S.; Harmar, A.J.; Waschek, J.; Herzog, E.D. Vasoactive intestinal polypeptide mediates circadian rhyth-micity and synchrony in mammalian clock neurons. Nat. Neurosci. 2005, 8, 476–483. [Google Scholar] [CrossRef]
- Vosko, A.M.; Schroeder, A.; Loh, D.; Colwell, C.S. Vasoactive intestinal peptide and the mammalian circadian system. Gen. Comp. Endocrinol. 2007, 152, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.L.; Kaufman, T.C. Hox genes and the evolution of the arthropod body plan1. Evol. Dev. 2002, 4, 459–499. [Google Scholar] [CrossRef]
- Singh, S.; Sanchez-Herrero, E.; Shashidhara, L.S. Critical role for Fat/Hippo and IIS/Akt pathways downstream of Ultrabi-thorax during haltere specification in Drosophila. Mech. Dev. 2015, 138, 198–209. [Google Scholar] [CrossRef]
- Gupta, R.P.; Bajpai, A.; Sinha, P. Selector genes display tumor cooperation and inhibition in Drosophila epithelium in a de-velopmental context-dependent manner. Biol. Open 2017, 6, 1581–1591. [Google Scholar] [CrossRef] [Green Version]
- Domsch, K.; Carnesecchi, J.; Disela, V.; Friedrich, J.; Trost, N.; Ermakova, O.; Polychronidou, M.; Lohmann, I. The Hox tran-scription factor Ubx stabilizes lineage commitment by suppressing cellular plasticity in Drosophila. Elife 2019, 8, e42675. [Google Scholar] [CrossRef] [PubMed]
- Domsch, K.; Schroder, J.; Janeschik, M.; Schaub, C.; Lohmann, I. The Hox Transcription Factor Ubx Ensures Somatic Myo-genesis by Suppressing the Mesodermal Master Regulator Twist. Cell Rep. 2021, 34, 108577. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Cui, L.; Zeng, Y.; Song, W.; Gaur, U.; Yang, M. 14–3-3 Proteins Are on the Crossroads of Cancer, Aging, and Age-Related Neurodegenerative Disease. Int. J. Mol. Sci. 2019, 20, 3518. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Huang, H.R.; Li, W.D.; Jin, X.J.; Zeng, J.; Liu, Y.W.; Gu, Y.; Sun, X.G.; Wen, G.; Ding, Y.Q.; et al. Nuclear local-ization of 14–3-3epsilon inversely correlates with poor long-term survival of patients with colorectal cancer. J. Surg. Oncol. 2012, 106, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Gu, H.; Li, J.; Xu, M.; Liu, T.; Shen, Y.; Chen, B.; Zhang, G. RKIP and 14–3-3ε Exert an Opposite Effect on Human Gastric Cancer Cells SGC7901 by Regulating the ERK/MAPK Pathway Differently. Dig. Dis. Sci. 2012, 58, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Yan, L.; Gu, H.; Mu, Y.; Tong, G.; Zhang, G. 14-3-3ε functions as an oncogene in SGC7901 gastric cancer cells through involvement of cyclin E and p27kip1. Mol. Med. Rep. 2014, 10, 3145–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Fang, X.; Fang, H.; Feng, Y.; Chen, F.; Xia, Q. ATPR-induced G(0)/G(1) phase arrest in gastric cancer cells by regulating the binding of 14-3-3ε and filamin A. Cancer Med. 2018, 7, 3373–3384. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.Y.; Ghelani, D.; Yeh, S.; Wu, K.K. Nonsteroidal anti-inflammatory drugs induce colorectal cancer cell apoptosis by suppressing 14-3-3epsilon. Cancer Res. 2007, 67, 3185–3191. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.K.; Liou, J.Y. Cyclooxygenase Inhibitors Induce Colon Cancer Cell Apoptosis Via PPAR delta -> 14-3-3 epsilon Pathway. Methods Mol. Biol. 2009, 512, 295–307. [Google Scholar]
- Soucise, A.; Vaughn, C.; Thompson, C.L.; Millen, A.E.; Freudenheim, J.L.; Wactawski-Wende, J.; Phipps, A.I.; Hale, L.; Qi, L.H.; Ochs-Balcom, H.M. Sleep quality, duration, and breast cancer aggressiveness. Breast Cancer Res. Tr. 2017, 164, 169–178. [Google Scholar] [CrossRef]
- Gabryelska, A.; Szmyd, B.; Szemraj, J.; Stawski, R.; Sochal, M.; Białasiewicz, P. Patients with obstructive sleep apnea present with chronic upregulation of serum HIF-1α protein. J. Clin. Sleep Med. 2020, 16, 1761–1768. [Google Scholar] [CrossRef]
- Gabryelska, A.; Szmyd, B.; Panek, M.; Szemraj, J.; Kuna, P.; Białasiewicz, P. Serum Hypoxia-Inducible Factor-1α protein level as a diagnostic marker of obstructive sleep apnea. Pol. Arch. Intern. Med. 2019, 130, 158–160. [Google Scholar] [CrossRef] [Green Version]
- Gabryelska, A.; Stawski, R.; Sochal, M.; Szmyd, B.; Białasiewicz, P. Influence of one-night CPAP therapy on the changes of HIF-1α protein in OSA patients: A pilot study. J. Sleep Res. 2020, 29, e12995. [Google Scholar] [CrossRef]
- Gabryelska, A.; Karuga, F.F.; Szmyd, B.; Białasiewicz, P. HIF-1α as a Mediator of Insulin Resistance, T2DM, and Its Compli-cations: Potential Links With Obstructive Sleep Apnea. Front. Physiol. 2020, 11, 1035. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Li, N.; Yao, X.; Zhou, L. Potential inflammatory markers in obstructive sleep apnea-hypopnea syndrome. Bosn. J. Basic Med. Sci. 2016, 17, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Gabryelska, A.; Sochal, M.; Turkiewicz, S.; Białasiewicz, P. Relationship between HIF-1 and Circadian Clock Proteins in Ob-structive Sleep Apnea Patients-Preliminary Study. J. Clin. Med. 2020, 9, 1599. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Du, J.; Zhao, Z. Integrative Role of 14-3-3ε in Sleep Regulation. Int. J. Mol. Sci. 2021, 22, 9748. https://doi.org/10.3390/ijms22189748
Wei Y, Du J, Zhao Z. Integrative Role of 14-3-3ε in Sleep Regulation. International Journal of Molecular Sciences. 2021; 22(18):9748. https://doi.org/10.3390/ijms22189748
Chicago/Turabian StyleWei, Yu, Juan Du, and Zhangwu Zhao. 2021. "Integrative Role of 14-3-3ε in Sleep Regulation" International Journal of Molecular Sciences 22, no. 18: 9748. https://doi.org/10.3390/ijms22189748
APA StyleWei, Y., Du, J., & Zhao, Z. (2021). Integrative Role of 14-3-3ε in Sleep Regulation. International Journal of Molecular Sciences, 22(18), 9748. https://doi.org/10.3390/ijms22189748