
A Journey on the Skin Microbiome: Pitfalls and Opportunities

 , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. References Analyzed and Methodologies of Study
3. The Skin Microbiota
3.1. Resident and Transient Skin Microbiota
3.2. Insight into the Core Microbiota of Derma and Adnexal Structures
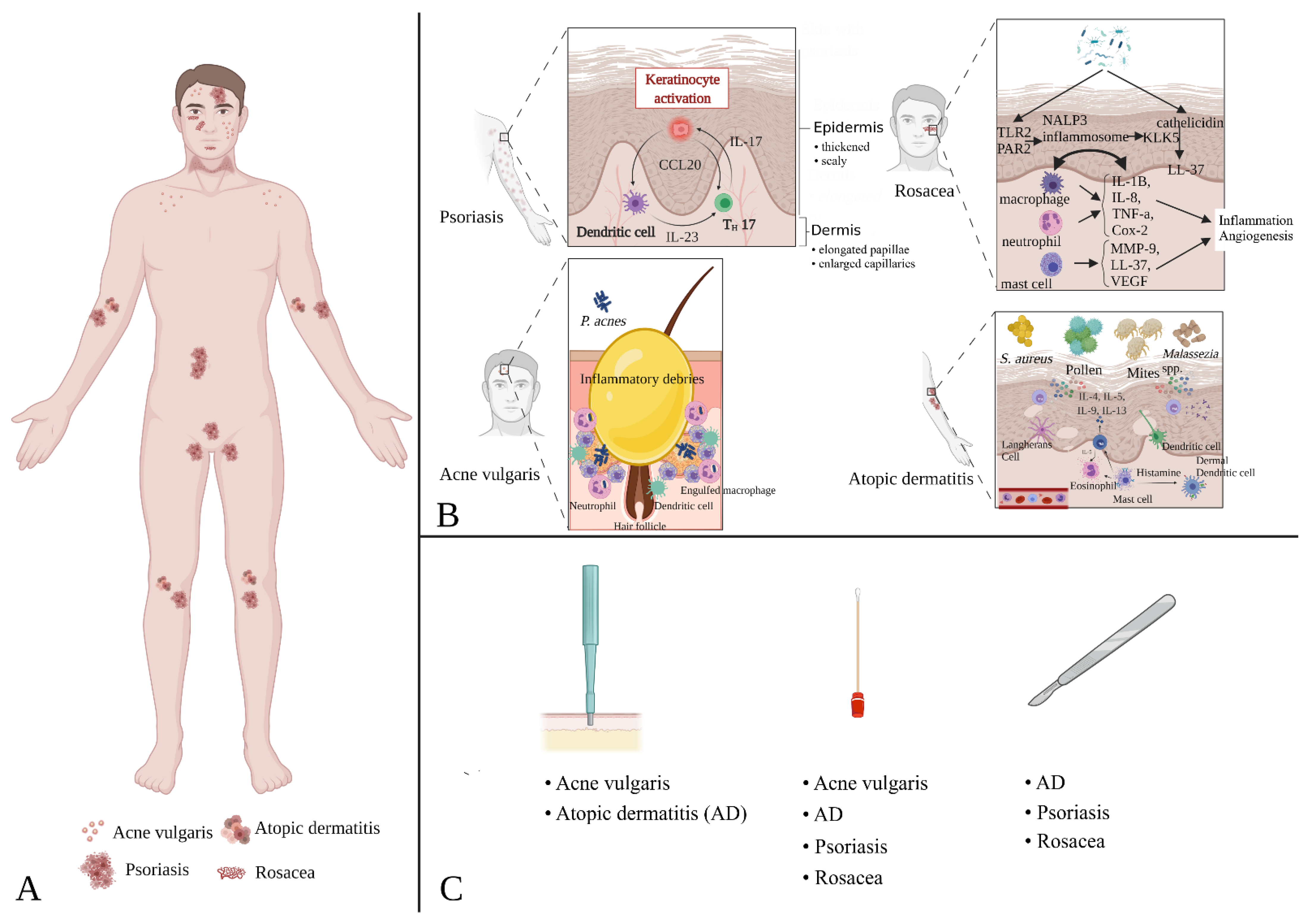
3.3. Dysbiosis of the Skin Microbiota in Specific Diseases
3.3.1. Acne Vulgaris
3.3.2. Atopic Dermatitis
3.3.3. Psoriasis
3.3.4. Rosacea
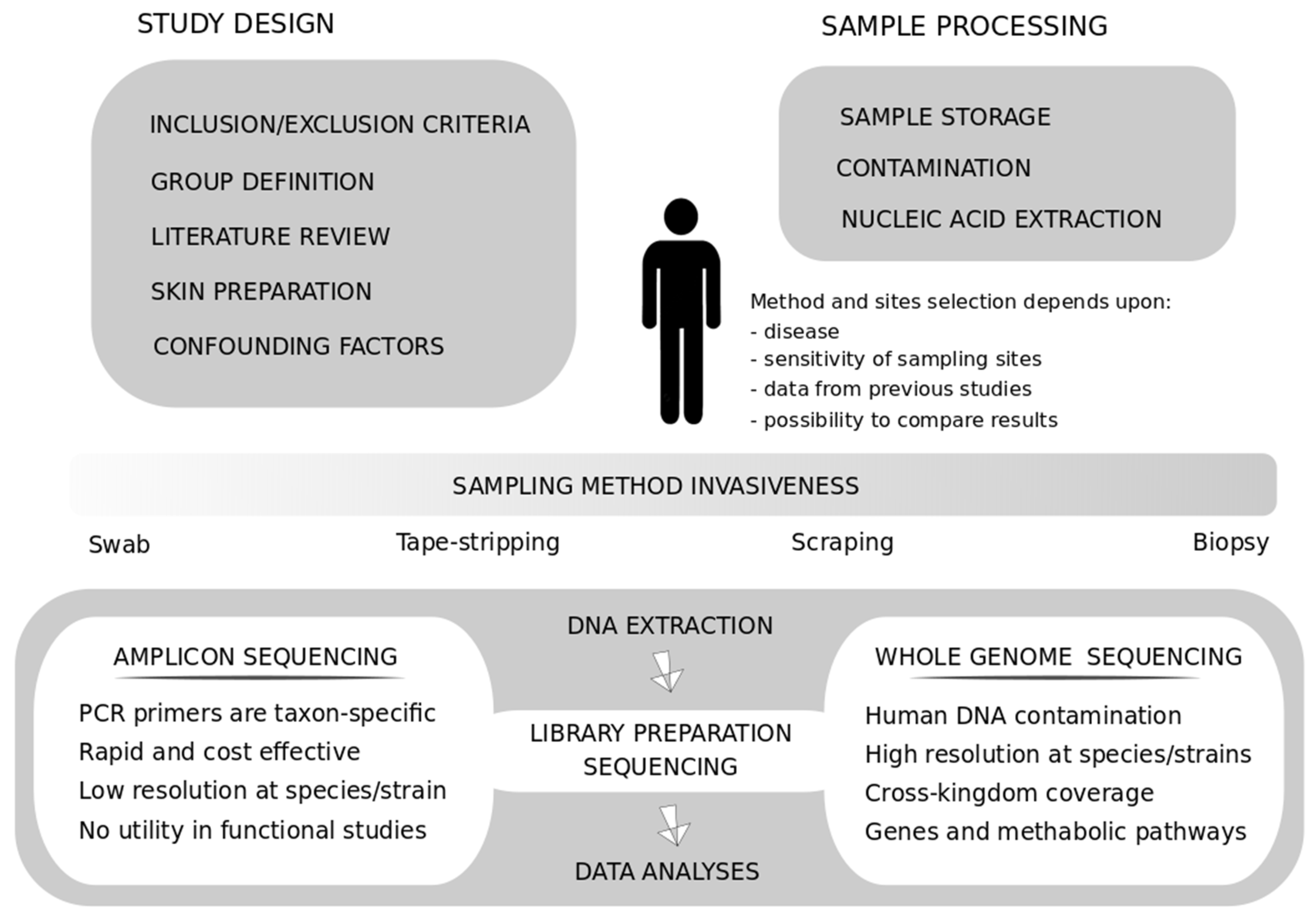
4. Methodological Considerations on NGS Techniques
5. Skin Sampling Procedures: Standardization and Reproducibility among Studies
Towards the Optimization of a Skin Disease-Based Sampling
6. Topical and Oral Probiotics in Skin Health and Diseases: State of the Art
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belkaid, Y.; Harrison, O.J. Homeostatic Immunity and the Microbiota. Immunity 2017, 46, 562–576. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Balato, A.; Cacciapuoti, S.; Di Caprio, R.; Marasca, C.; Masarà, A.; Raimondo, A.; Fabbrocini, G. Human Microbiome: Composition and Role in Inflammatory Skin Diseases. Arch. Immunol. Ther. Exp. 2019, 67, 1–18. [Google Scholar] [CrossRef]
- Nelson, K.; Weinstock, G.; Highlander, S.; Worley, K.; Creasy, H.; Wortman, J.; Rusch, D.; Mitreva, M.; Sodergren, E.; Chinwalla, A.; et al. A Catalog of Reference Genomes from the Human Microbiome. Science 2010, 328, 994–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, L.; Creasy, H.; Fettweis, J.; Lloyd-Price, J.; Mahurkar, A.; Zhou, W.; Buck, G.A.; Snyder, M.P.; Strauss, J.; Weinstock, G.M.; et al. The Integrative Human Microbiome Project. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xia, J.; Jiang, L.; Tan, Y.; An, Y.; Zhu, X.; Ruan, J.; Chen, Z.; Zhen, H.; Ma, Y.; et al. Characterization of the Human Skin Resistome and Identification of Two Microbiota Cutotypes. Microbiome 2021, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Panelli, S.; Bandi, C.; Zuccotti, G.; Perini, M.; D’Auria, E. What Pediatricians Should Know Before Studying Gut Microbiota. J. Clin. Med. 2019, 8, 1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panelli, S.; Epis, S.; Cococcioni, L.; Perini, M.; Paroni, M.; Bandi, C.; Drago, L.; Zuccotti, G.V. Inflammatory Bowel Diseases, the Hygiene Hypothesis and the Other Side of the Microbiota: Parasites and Fungi. Pharmacol. Res. 2020, 159, 104962. [Google Scholar] [CrossRef]
- Poussin, C.; Sierro, N.; Boué, S.; Battey, J.; Scotti, E.; Belcastro, V.; Peitsch, M.C.; Ivanov, N.V.; Hoeng, J. Interrogating the Microbiome: Experimental and Computational Considerations in Support of Study Reproducibility. Drug Discov. Today 2018, 23, 1644–1657. [Google Scholar] [CrossRef]
- Bay, L.; Barnes, C.J.; Fritz, B.G.; Thorsen, J.; Restrup, M.E.M.; Rasmussen, L.; Sørensen, J.K.; Hesselvig, A.B.; Odgaard, A.; Hansen, A.J.; et al. Universal Dermal Microbiome in Human Skin. mBio 2020, 11, e02945-19. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Byrd, A.L.; Deming, C.; Conlan, S.; Kong, H.H.; Segre, J.A. Biogeography and Individuality Shape Function in the Human Skin Metagenome. Nature 2014, 514, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; Green, E.D.; et al. Topographical and Temporal Diversity of the Human Skin Microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A.; Kong, H.H.; Renaud, G.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Wolfsberg, T.G.; Turner, M.L.; Segre, J.A. A Diversity Profile of the Human Skin Microbiota. Genome Res. 2008, 18, 1043–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grogan, M.D.; Bartow-McKenney, C.; Flowers, L.; Knight, S.A.B.; Uberoi, A.; Grice, E.A. Research Techniques Made Simple: Profiling the Skin Microbiota. J. Investig. Dermatol. 2019, 139, 747–752.e1. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.H. Skin Microbiome: Genomics-Based Insights into the Diversity and Role of Skin Microbes. Trends Mol. Med. 2011, 17, 320–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, H.H.; Andersson, B.; Clavel, T.; Common, J.E.; Jackson, S.A.; Olson, N.D.; Segre, J.A.; Traidl-Hoffmann, C. Performing Skin Microbiome Research: A Method to the Madness. J. Investig. Dermatol. 2017, 137, 561–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ederveen, T.H.A.; Smits, J.P.H.; Boekhorst, J.; Schalkwijk, J.; van den Bogaard, E.H.; Zeeuwen, P.L.J.M. Skin Microbiota in Health and Disease: From Sequencing to Biology. J. Dermatol. 2020, 47, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Godlewska, U.; Brzoza, P.; Kwiecień, K.; Kwitniewski, M.; Cichy, J. Metagenomic Studies in Inflammatory Skin Diseases. Curr. Microbiol. 2020, 77, 3201–3212. [Google Scholar] [CrossRef] [PubMed]
- Khayyira, A.S.; Rosdina, A.E.; Irianti, M.I.; Malik, A. Simultaneous Profiling and Cultivation of the Skin Microbiome of Healthy Young Adult Skin for the Development of Therapeutic Agents. Heliyon 2020, 6, e03700. [Google Scholar] [CrossRef]
- Meisel, J.S.; Hannigan, G.D.; Tyldsley, A.S.; SanMiguel, A.J.; Hodkinson, B.P.; Zheng, Q.; Grice, E.A. Skin Microbiome Surveys Are Strongly Influenced by Experimental Design. J. Investig. Dermatol. 2016, 136, 947–956. [Google Scholar] [CrossRef]
- Reiger, M.; Traidl-Hoffmann, C.; Neumann, A.U. The Skin Microbiome as a Clinical Biomarker in Atopic Eczema: Promises, Navigation, and Pitfalls. J. Allergy Clin. Immunol. 2020, 145, 93–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, S.S.; Pourang, A.; Sivamani, R.K. A Review of next Generation Sequencing Technologies Used in the Evaluation of the Skin Microbiome: What a Time to Be Alive. Dermatol. Online J. 2019, 25, 9. [Google Scholar]
- Gallo, R.L. Human Skin is the Largest Epithelial Surface for Interaction with Microbes. J. Investig. Dermatol. 2017, 137, 1213–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grice, E.A.; Segre, J.A. The Skin Microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D Metabolism and Function in the Skin. Mol. Cell. Endocrinol. 2011, 347, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin Microbiota–Host Interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef]
- Logan, A.C.; Jacka, F.N.; Prescott, S.L. Immune-Microbiota Interactions: Dysbiosis as a Global Health Issue. Curr. Allergy Asthma Rep. 2016, 16, 13. [Google Scholar] [CrossRef]
- Prescott, S.L.; Larcombe, D.-L.; Logan, A.C.; West, C.; Burks, W.; Caraballo, L.; Levin, M.; Etten, E.V.; Horwitz, P.; Kozyrskyj, A.; et al. The Skin Microbiome: Impact of Modern Environments on Skin Ecology, Barrier Integrity, and Systemic Immune Programming. World Allergy Organ. J. 2017, 10, 29. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mitra, R.; Maitra, A.; Gupta, S.; Kumaran, S.; Chakrabortty, A.; Majumder, P.P. Sebum and Hydration Levels in Specific Regions of Human Face Significantly Predict the Nature and Diversity of Facial Skin Microbiome. Sci. Rep. 2016, 6, 36062. [Google Scholar] [CrossRef]
- Dréno, B.; Araviiskaia, E.; Berardesca, E.; Gontijo, G.; Sanchez Viera, M.; Xiang, L.F.; Martin, R.; Bieber, T. Microbiome in Healthy Skin, Update for Dermatologists. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 2038–2047. [Google Scholar] [CrossRef]
- Méric, G.; Mageiros, L.; Pensar, J.; Laabei, M.; Yahara, K.; Pascoe, B.; Kittiwan, N.; Tadee, P.; Post, V.; Lamble, S.; et al. Disease-Associated Genotypes of the Commensal Skin Bacterium Staphylococcus Epidermidis. Nat. Commun. 2018, 9, 5034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Staphylococcus Epidermidis–the “Accidental” Pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.J.C.; Abildgaard, J.T.; Backes, J.; Hawkins, R.J. Preventing Infection in Shoulder Surgery. J. Shoulder Elbow Surg. 2018, 27, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Speer, C.P. The Role of Staphylococcus Epidermidis in Neonatal Sepsis: Guarding Angel or Pathogenic Devil? Int. J. Med. Microbiol. 2014, 304, 513–520. [Google Scholar] [CrossRef]
- Szczuka, E.; Telega, K.; Kaznowski, A. Biofilm Formation by Staphylococcus Hominis Strains Isolated from Human Clinical Specimens. Folia Microbiol. 2015, 60, 1–5. [Google Scholar] [CrossRef]
- Jeon, J.; Park, S.C.; Her, J.; Lee, J.W.; Han, J.-K.; Kim, Y.-K.; Kim, K.P.; Ban, C. Comparative Lipidomic Profiling of the Human Commensal Bacterium Propionibacterium Acnes and Its Extracellular Vesicles. RSC Adv. 2018, 8, 15241–15247. [Google Scholar] [CrossRef] [Green Version]
- Boisrenoult, P. Cutibacterium Acnes Prosthetic Joint Infection: Diagnosis and Treatment. Orthop. Traumatol. Surg. Res. 2018, 104, S19–S24. [Google Scholar] [CrossRef] [PubMed]
- Elston, M.J.; Dupaix, J.P.; Opanova, M.I.; Atkinson, R.E. Cutibacterium Acnes (Formerly Proprionibacterium Acnes) and Shoulder Surgery. Hawaii J. Health Soc. Welf. 2019, 78, 3–5. [Google Scholar]
- Freedman, M.; Aflatooni, J.O.; Foster, R.; Haggerty, P.G.; Derber, C.J. Persistent Cutibacterium (Formerly Propionibacterium) Acnes Bacteremia and Refractory Endocarditis in a Patient with Retained Implantable Pacemaker Leads. Case Rep. Infect. Dis. 2020, 2020, e8883907. [Google Scholar] [CrossRef]
- Moissl-Eichinger, C.; Probst, A.J.; Birarda, G.; Auerbach, A.; Koskinen, K.; Wolf, P.; Holman, H.-Y.N. Human Age and Skin Physiology Shape Diversity and Abundance of Archaea on Skin. Sci. Rep. 2017, 7, 4039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probst, A.J.; Auerbach, A.K.; Moissl-Eichinger, C. Archaea on Human Skin. PLoS ONE 2013, 8, e65388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litwin, D.; Chen, W.; Dzika, E.; Korycinska, J. Human Permanent Ectoparasites; Recent Advances on Biology and Clinical Significance of Demodex Mites: Narrative Review Article. Iran J. Parasitol. 2017, 12, 12–21. [Google Scholar] [PubMed]
- Czepita, D.; Kuźna-Grygiel, W.; Czepita, M.; Grobelny, A. Demodex Folliculorum and Demodex Brevis as a Cause of Chronic Marginal Blepharitis. Ann. Acad. Med. Stetin. 2007, 53, 63–67, Discussion 67. [Google Scholar] [PubMed]
- Liang, L.; Ding, X.; Tseng, S.C.G. High Prevalence of Demodex Brevis Infestation in Chalazia. Am. J. Ophthalmol. 2014, 157, 342–348.e1. [Google Scholar] [CrossRef]
- Moravvej, H.; Dehghan-Mangabadi, M.; Abbasian, M.-R.; Meshkat-Razavi, G. Association of Rosacea with Demodicosis. Arch. Iran Med. 2007, 10, 199–203. [Google Scholar]
- Foulongne, V.; Sauvage, V.; Hebert, C.; Dereure, O.; Cheval, J.; Gouilh, M.A.; Pariente, K.; Segondy, M.; Burguière, A.; Manuguerra, J.-C.; et al. Human Skin Microbiota: High Diversity of DNA Viruses Identified on the Human Skin by High Throughput Sequencing. PLoS ONE 2012, 7, e38499. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Madupu, R.; Karaoz, U.; Nossa, C.W.; Yang, L.; Yooseph, S.; Yachimski, P.S.; Brodie, E.L.; Nelson, K.E.; Pei, Z. Human Papillomavirus Community in Healthy Persons, Defined by Metagenomics Analysis of Human Microbiome Project Shotgun Sequencing Data Sets. J. Virol. 2014, 88, 4786–4797. [Google Scholar] [CrossRef] [Green Version]
- Verbanic, S.; Kim, C.Y.; Deacon, J.M.; Chen, I.A. Improved Single-Swab Sample Preparation for Recovering Bacterial and Phage DNA from Human Skin and Wound Microbiomes. BMC Microbiol. 2019, 19, 214. [Google Scholar] [CrossRef] [Green Version]
- Leung, M.H.Y.; Tong, X.; Bøifot, K.O.; Bezdan, D.; Butler, D.J.; Danko, D.C.; Gohli, J.; Green, D.C.; Hernandez, M.T.; Kelly, F.J.; et al. Characterization of the Public Transit Air Microbiome and Resistome Reveals Geographical Specificity. Microbiome 2021, 9, 112. [Google Scholar] [CrossRef]
- Prussin, A.J.; Marr, L.C. Sources of Airborne Microorganisms in the Built Environment. Microbiome 2015, 3, 78. [Google Scholar] [CrossRef] [Green Version]
- Shibagaki, N.; Suda, W.; Clavaud, C.; Bastien, P.; Takayasu, L.; Iioka, E.; Kurokawa, R.; Yamashita, N.; Hattori, Y.; Shindo, C.; et al. Aging-Related Changes in the Diversity of Women’s Skin Microbiomes Associated with Oral Bacteria. Sci. Rep. 2017, 7, 10567. [Google Scholar] [CrossRef] [PubMed]
- Jugé, R.; Rouaud-Tinguely, P.; Breugnot, J.; Servaes, K.; Grimaldi, C.; Roth, M.-P.; Coppin, H.; Closs, B. Shift in Skin Microbiota of Western European Women across Aging. J. Appl. Microbiol. 2018, 125, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bai, X.; Peng, T.; Yi, X.; Luo, L.; Yang, J.; Liu, J.; Wang, Y.; He, T.; Wang, X.; et al. New Insights Into the Skin Microbial Communities and Skin Aging. Front. Microbiol. 2020, 11, 565549. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, J.J.; Myeong, N.R.; Kim, T.; Kim, D.; An, S.; Kim, H.; Park, T.; Jang, S.I.; Yeon, J.H.; et al. Segregation of Age-Related Skin Microbiome Characteristics by Functionality. Sci. Rep. 2019, 9, 16748. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Haiminen, N.; Carrieri, A.-P.; Hu, R.; Jiang, L.; Parida, L.; Russell, B.; Allaband, C.; Zarrinpar, A.; Vázquez-Baeza, Y.; et al. Human Skin, Oral, and Gut Microbiomes Predict Chronological Age. Msystems 2020, 5, e00630-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abadías-Granado, I.; Sánchez-Bernal, J.; Gilaberte, Y. The Microbiome and Aging. Plast. Aesthetic Res. 2021, 8, 27. [Google Scholar] [CrossRef]
- Boxberger, M.; Cenizo, V.; Cassir, N.; La Scola, B. Challenges in Exploring and Manipulating the Human Skin Microbiome. Microbiome 2021, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Dimitriu, P.A.; Iker, B.; Malik, K.; Leung, H.; Mohn, W.W.; Hillebrand, G.G. New Insights into the Intrinsic and Extrinsic Factors That Shape the Human Skin Microbiome. mBio 2019, 10, e00839-19. [Google Scholar] [CrossRef] [Green Version]
- Staudinger, T.; Pipal, A.; Redl, B. Molecular Analysis of the Prevalent Microbiota of Human Male and Female Forehead Skin Compared to Forearm Skin and the Influence of Make-Up. J. Appl. Microbiol. 2011, 110, 1381–1389. [Google Scholar] [CrossRef]
- Lee, H.J.; Jeong, S.E.; Lee, S.; Kim, S.; Han, H.; Jeon, C.O. Effects of Cosmetics on the Skin Microbiome of Facial Cheeks with Different Hydration Levels. Microbiologyopen 2018, 7, e00557. [Google Scholar] [CrossRef] [PubMed]
- Fredricks, D.N. Microbial Ecology of Human Skin in Health and Disease. J. Investig. Dermatol. Symp. Proc. 2001, 6, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.B.; Byun, E.J.; Kim, H.S. Potential Role of the Microbiome in Acne: A Comprehensive Review. J. Clin. Med. 2019, 8, 987. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.H.; Segre, J.A. Skin Microbiome: Looking Back to Move Forward. J. Investig. Dermatol. 2012, 132, 933–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schommer, N.N.; Gallo, R.L. Structure and Function of the Human Skin Microbiome. Trends Microbiol. 2013, 21, 660–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woese, C.R.; Fox, G.E. Phylogenetic Structure of the Prokaryotic Domain: The Primary Kingdoms. Proc. Natl. Acad. Sci. USA 1977, 74, 5088–5090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial Community Variation in Human Body Habitats Across Space and Time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Costello, E.K.; Berg-Lyons, D.; Gonzalez, A.; Stombaugh, J.; Knights, D.; Gajer, P.; Ravel, J.; Fierer, N.; et al. Moving Pictures of the Human Microbiome. Genome Biol. 2011, 12, R50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.; Byrd, A.L.; Park, M.; Kong, H.H.; Segre, J.A. Temporal Stability of the Human Skin Microbiome. Cell 2016, 165, 854–866. [Google Scholar] [CrossRef] [Green Version]
- Lilly, H.A.; Lowbury, E.J. Transient Skin Flora: Their Removal by Cleansing or Disinfection in Relation to their Mode of Deposition. J. Clin. Pathol. 1978, 31, 919–922. [Google Scholar] [CrossRef] [Green Version]
- Mutters, R.; Warnes, S.L. The Method Used to Dry Washed Hands Affects the Number and Type of Transient and Residential Bacteria Remaining on the Skin. J. Hosp. Infect. 2019, 101, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.C.; Jiang, S.C. Alterations of the Human Skin Microbiome after Ocean Water Exposure. Mar. Pollut. Bull. 2019, 145, 595–603. [Google Scholar] [CrossRef]
- Wallen-Russell, C. The Role of Every-Day Cosmetics in Altering the Skin Microbiome: A Study Using Biodiversity. Cosmetics 2019, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, T.F.; Noe, J.C.; Patterson, A.T.; Lenz, B.L.; Beachkofsky, T.M. Temporal Shifts in the Collective Dermatologic Microbiome of Military Trainees. Clin. Cosmet. Investig. Dermatol. 2019, 12, 625–637. [Google Scholar] [CrossRef] [Green Version]
- Krishna, S.; Miller, L.S. Host-Pathogen Interactions between the Skin and Staphylococcus Aureus. Curr. Opin. Microbiol. 2012, 15, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Polak-Witka, K.; Rudnicka, L.; Blume-Peytavi, U.; Vogt, A. The Role of the Microbiome in Scalp Hair Follicle Biology and Disease. Exp. Dermatol. 2020, 29, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavaud, C.; Jourdain, R.; Bar-Hen, A.; Tichit, M.; Bouchier, C.; Pouradier, F.; El Rawadi, C.; Guillot, J.; Ménard-Szczebara, F.; Breton, L.; et al. Dandruff Is Associated with Disequilibrium in the Proportion of the Major Bacterial and Fungal Populations Colonizing the Scalp. PLoS ONE 2013, 8, e58203. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Wang, Z.; Yuan, C.; Liu, X.; Yang, F.; Wang, T.; Wang, J.; Manabe, K.; Qin, O.; Wang, X.; et al. Dandruff Is Associated with the Conjoined Interactions between Host and Microorganisms. Sci. Rep. 2016, 6, 24877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkac, L.; Clarke, T.H.; Singh, H.; Greco, C.; Gomez, A.; Torralba, M.G.; Frank, B.; Nelson, K.E. Spatial and Environmental Variation of the Human Hair Microbiota. Sci. Rep. 2018, 8, 9017. [Google Scholar] [CrossRef] [Green Version]
- Mengist, A.; Aschale, Y.; Reta, A. Bacterial and Parasitic Assessment from Fingernails in Debre Markos, Northwest Ethiopia. Available online: https://www.hindawi.com/journals/cjidmm/2018/6532014/ (accessed on 30 June 2021).
- Micheal, A.O.; Elisabeth, P.T.; Adenike, K.K.; Olajumoke, A.E.; Adebiyi, A.; Olutope, O.S.; Ademola, A.D. Isolation and Molecular Characterization of Mycobiota and Other Microbiota from Fingernails. Int. J. Clin. Dev. Anat. 2021, 7, 1. [Google Scholar] [CrossRef]
- Wałaszek, M.Z.; Kołpa, M.; Różańska, A.; Jagiencarz-Starzec, B.; Wolak, Z.; Wójkowska-Mach, J. Nail Microbial Colonization Following Hand Disinfection: A Qualitative Pilot Study. J. Hosp. Infect. 2018, 100, 207–210. [Google Scholar] [CrossRef] [PubMed]
- De Pessemier, B.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut-Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef]
- Egert, M.; Simmering, R.; Riedel, C.U. The Association of the Skin Microbiota with Health, Immunity, and Disease. Clin. Pharmacol. Ther. 2017, 102, 62–69. [Google Scholar] [CrossRef]
- Forton, F. Demodex-Associated Folliculitis. Am. J. Dermatopathol. 1998, 20, 536–537. [Google Scholar] [CrossRef]
- Forton, F.M.N.; De Maertelaer, V. Papulopustular Rosacea and Rosacea-like Demodicosis: Two Phenotypes of the Same Disease? J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Jarmuda, S.; O’Reilly, N.; Żaba, R.; Jakubowicz, O.; Szkaradkiewicz, A.; Kavanagh, K. 2012 Potential Role of Demodex Mites and Bacteria in the Induction of Rosacea. J. Med. Microbiol. 2012, 61, 1504–1510. [Google Scholar] [CrossRef]
- Forton, F.; Germaux, M.-A.; Brasseur, T.; De Liever, A.; Laporte, M.; Mathys, C.; Sass, U.; Stene, J.-J.; Thibaut, S.; Tytgat, M.; et al. Demodicosis and Rosacea: Epidemiology and Significance in Daily Dermatologic Practice. J. Am. Acad. Dermatol. 2005, 52, 74–87. [Google Scholar] [CrossRef]
- Lacey, N.; Russell-Hallinan, A.; Zouboulis, C.C.; Powell, F.C. Demodex Mites Modulate Sebocyte Immune Reaction: Possible Role in the Pathogenesis of Rosacea. Br. J. Dermatol. 2018, 179, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-S.; Huang, Y.-C. Role of Demodex Mite Infestation in Rosacea: A Systematic Review and Meta-Analysis. J. Am. Acad. Dermatol. 2017, 77, 441–447.e6. [Google Scholar] [CrossRef] [PubMed]
- Yang, X. Relationship between Helicobacter Pylori and Rosacea: Review and Discussion. BMC Infect. Dis. 2018, 18, 318. [Google Scholar] [CrossRef] [Green Version]
- Lazaridou, E.; Korfitis, C.; Kemanetzi, C.; Sotiriou, E.; Apalla, Z.; Vakirlis, E.; Fotiadou, C.; Lallas, A.; Ioannides, D. Rosacea and Helicobacter Pylori: Links and Risks. Clin. Cosmet. Investig. Dermatol. 2017, 10, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfeld, M.; Gunasingam, N.; Leow, L.J.; Shirato, K.; Preda, V. Staphylococcus Epidermidis: A Possible Role in the Pustules of Rosacea. J. Am. Acad. Dermatol. 2011, 64, 49–52. [Google Scholar] [CrossRef]
- Fernandez-Obregon, A.; Patton, D.L. The Role of Chlamydia Pneumoniae in the Etiology of Acne Rosacea: Response to the Use of Oral Azithromycin. Cutis 2007, 79, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Obregon, A. Commentary on “The Role of Chlamydia Pneumoniae in the Etiology of Acne Rosacea: Response to the Use of Oral Azithromycin”. J. Infect. 2019, 2, 11–14. [Google Scholar] [CrossRef]
- Geoghegan, J.A.; Irvine, A.D.; Foster, T.J. Staphylococcus Aureus and Atopic Dermatitis: A Complex and Evolving Relationship. Trends Microbiol. 2018, 26, 484–497. [Google Scholar] [CrossRef]
- Iwamoto, K.; Moriwaki, M.; Miyake, R.; Hide, M. Staphylococcus Aureus in Atopic Dermatitis: Strain-Specific Cell Wall Proteins and Skin Immunity. Allergol. Int. 2019, 68, 309–315. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from Human Skin Commensal Bacteria Protect against Staphylococcus Aureus and are Deficient in Atopic Dermatitis. Sci. Transl. Med. 2017, 9, eaah4680. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.; Jaros, J.; Shi, V.Y. Staphylococcus Aureus in Atopic Dermatitis: Past, Present, and Future. Dermatitis 2020, 31, 247–258. [Google Scholar] [CrossRef]
- Hepburn, L.; Hijnen, D.J.; Sellman, B.R.; Mustelin, T.; Sleeman, M.A.; May, R.D.; Strickland, I. The Complex Biology and Contribution of Staphylococcus Aureus in Atopic Dermatitis, Current and Future Therapies. Br. J. Dermatol. 2017, 177, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.Y.; Leung, D.Y.M. Bacterial and Viral Infections in Atopic Dermatitis: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2016, 51, 329–337. [Google Scholar] [CrossRef]
- Saintive, S.; Abad, E.; de Ferreira, C.D.; Stambovsky, M.; Cavalcante, F.S.; Gonçalves, L.S.; Vidal, F.; Dos Santos, K.R. What is the Role of Staphylococcus Aureus and Herpes Virus Infections in the Pathogenesis of Atopic Dermatitis? Future Microbiol. 2017, 12, 1327–1334. [Google Scholar] [CrossRef]
- Rademaker, M.; Agnew, K.; Anagnostou, N.; Andrews, M.; Armour, K.; Baker, C.; Foley, P.; Gebauer, K.; Gupta, M.; Marshman, G.; et al. Psoriasis and Infection. A Clinical Practice Narrative. Australas J. Dermatol. 2019, 60, 91–98. [Google Scholar] [CrossRef]
- Ng, C.Y.; Huang, Y.H.; Chu, C.F.; Wu, T.C.; Liu, S.H. Risks for Staphylococcus Aureus Colonization in Patients with Psoriasis: A Systematic Review and Meta-Analysis. Br. J. Dermatol. 2017, 177, 967–977. [Google Scholar] [CrossRef]
- Queirós, N.; Torres, T. HIV-Associated Psoriasis. Actas Dermosifiliogr. (Engl. Ed.) 2018, 109, 303–311. [Google Scholar] [CrossRef]
- Thio, H.B. The Microbiome in Psoriasis and Psoriatic Arthritis: The Skin Perspective. J. Rheumatol. Suppl. 2018, 94, 30–31. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Issa, N.; Afifi, L.; Jeon, C.; Chang, H.W.; Liao, W. The Role of the Skin and Gut Microbiome in Psoriatic Disease. Curr. Dermatol. Rep. 2017, 6, 94–103. [Google Scholar] [CrossRef]
- de Jesús-Gil, C.; Sans-de San Nicolàs, L.; Ruiz-Romeu, E.; Ferran, M.; Soria-Martínez, L.; García-Jiménez, I.; Chiriac, A.; Casanova-Seuma, J.M.; Fernández-Armenteros, J.M.; Owens, S.; et al. Interplay between Humoral and CLA+ T Cell Response against Candida Albicans in Psoriasis. Int. J. Mol. Sci. 2021, 22, 1519. [Google Scholar] [CrossRef]
- Platsidaki, E.; Dessinioti, C. Recent Advances in Understanding Propionibacterium Acnes (Cutibacterium acnes) in Acne. F1000Res 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, A.; Mozafarpoor, S.; Bodaghabadi, M.; Mohamadi, M. The Potential of Probiotics for Treating Acne Vulgaris: A Review of Literature on Acne and Microbiota. Dermatol. Ther. 2020, 33, e13279. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, H. Acne, the Skin Microbiome, and Antibiotic Treatment. Am. J. Clin. Dermatol. 2019, 20, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Law, M.P.M.; Chuh, A.A.T.; Lee, A.; Molinari, N. Acne Prevalence and beyond: Acne Disability and Its Predictive Factors among Chinese Late Adolescents in Hong Kong. Clin. Exp. Dermatol. 2010, 35, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Lynn, D.D.; Umari, T.; Dunnick, C.A.; Dellavalle, R.P. The Epidemiology of Acne Vulgaris in Late Adolescence. Adolesc. Health Med. Ther. 2016, 7, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayel, K.; Attia, M.; Agamia, N.; Fadl, N. Acne Vulgaris: Prevalence, Severity, and Impact on Quality of Life and Self-Esteem among Egyptian Adolescents. J. Egypt. Public Health Assoc. 2020, 95, 30. [Google Scholar] [CrossRef] [PubMed]
- Heng, A.H.S.; Chew, F.T. Systematic Review of the Epidemiology of Acne Vulgaris. Sci. Rep. 2020, 10, 5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitz-Gibbon, S.; Tomida, S.; Chiu, B.-H.; Nguyen, L.; Du, C.; Liu, M.; Elashoff, D.; Erfe, M.C.; Loncaric, A.; Kim, J.; et al. Propionibacterium Acnes Strain Populations in the Human Skin Microbiome Associated with Acne. J. Investig. Dermatol. 2013, 133, 2152–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, D.; Shi, B.; Erfe, M.C.; Craft, N.; Li, H. Vitamin B12 Modulates the Transcriptome of the Skin Microbiota in Acne Pathogenesis. Sci. Transl. Med. 2015, 7, 293ra103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, S.; Barnard, E.; Dawson, T.L.; Li, H. The Role of the Skin Microbiota in Acne Pathophysiology. Br. J. Dermatol. 2019, 181, 691–699. [Google Scholar] [CrossRef]
- Bjerre, R.D.; Bandier, J.; Skov, L.; Engstrand, L.; Johansen, J.D. The Role of the Skin Microbiome in Atopic Dermatitis: A Systematic Review. Br. J. Dermatol. 2017, 177, 1272–1278. [Google Scholar] [CrossRef]
- Seite, S.; Bieber, T. Barrier Function and Microbiotic Dysbiosis in Atopic Dermatitis. Clin. Cosmet. Investig. Dermatol. 2015, 8, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Weidinger, S.; Novak, N. Atopic Dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Di Domenico, E.G.; Cavallo, I.; Capitanio, B.; Ascenzioni, F.; Pimpinelli, F.; Morrone, A.; Ensoli, F. Staphylococcus Aureus and the Cutaneous Microbiota Biofilms in the Pathogenesis of Atopic Dermatitis. Microorganisms 2019, 7, 301. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.R.; Gallo, R.L. Evidence That Human Skin Microbiome Dysbiosis Promotes Atopic Dermatitis. J. Investig. Dermatol. 2017, 137, 2460–2461. [Google Scholar] [CrossRef]
- Edslev, S.M.; Agner, T.; Andersen, P.S. Skin Microbiome in Atopic Dermatitis. Acta Derm. Venereol. 2020, 100, adv00164. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-Y.; Lee, E.; Park, Y.M.; Hong, S.-J. Microbiome in the Gut-Skin Axis in Atopic Dermatitis. Allergy Asthma Immunol. Res. 2018, 10, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Reddel, S.; Del Chierico, F.; Quagliariello, A.; Giancristoforo, S.; Vernocchi, P.; Russo, A.; Fiocchi, A.; Rossi, P.; Putignani, L.; El Hachem, M. Gut Microbiota Profile in Children Affected by Atopic Dermatitis and Evaluation of Intestinal Persistence of a Probiotic Mixture. Sci. Rep. 2019, 9, 4996. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Liang, H.; Wang, Y.; Miao, M.; Shi, T.; Yang, F.; Liu, E.; Yuan, W.; Ji, Z.-S.; Li, D.-K. Altered Gut Microbiota Composition Associated with Eczema in Infants. PLoS ONE 2016, 11, e0166026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalliomäki, M.; Kirjavainen, P.; Eerola, E.; Kero, P.; Salminen, S.; Isolauri, E. Distinct Patterns of Neonatal Gut Microflora in Infants in Whom Atopy Was and Was Not Developing. J. Allergy Clin. Immunol. 2001, 107, 129–134. [Google Scholar] [CrossRef]
- Panelli, S.; Schneider, L.; Comandatore, F.; Bandi, C.; Zuccotti, G.V.; D’Auria, E. Is There Life in the Meconium? A Challenging, Burning Question. Pharmacol. Res. 2018, 137, 148–149. [Google Scholar] [CrossRef]
- Perez-Muñoz, M.E.; Arrieta, M.-C.; Ramer-Tait, A.E.; Walter, J. A Critical Assessment of the “Sterile Womb” and “in Utero Colonization” Hypotheses: Implications for Research on the Pioneer Infant Microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Segre, J.A. The Human Microbiome: Our Second Genome. Annu. Rev. Genom. Hum. Genet. 2012, 13, 151–170. [Google Scholar] [CrossRef] [Green Version]
- Langley, R.G.B.; Krueger, G.G.; Griffiths, C.E.M. Psoriasis: Epidemiology, Clinical Features, and Quality of Life. Ann. Rheum. Dis. 2005, 64, ii18–ii23. [Google Scholar] [CrossRef] [Green Version]
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M. National, Regional, and Worldwide Epidemiology of Psoriasis: Systematic Analysis and Modelling Study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef]
- Schön, M.P.; Boehncke, W.-H. Psoriasis. Available online: https://www.nejm.org/doi/10.1056/NEJMra041320 (accessed on 10 February 2021).
- Gao, Z.; Tseng, C.; Strober, B.E.; Pei, Z.; Blaser, M.J. Substantial Alterations of the Cutaneous Bacterial Biota in Psoriatic Lesions. PLoS ONE 2008, 3, e2719. [Google Scholar] [CrossRef]
- Gether, L.; Overgaard, L.K.; Egeberg, A.; Thyssen, J.P. Incidence and Prevalence of Rosacea: A Systematic Review and Meta-Analysis. Br. J. Dermatol. 2018, 179, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Rainer, B.M.; Kang, S.; Chien, A.L. Rosacea: Epidemiology, Pathogenesis, and Treatment. Dermato-Endocrinology 2017, 9, e1361574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daou, H.; Paradiso, M.; Hennessy, K.; Seminario-Vidal, L. Rosacea and the Microbiome: A Systematic Review. Dermatol. Ther. (Heidelb.) 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Kim, H.S. Microbiota in Rosacea|SpringerLink. Available online: https://link.springer.com/article/10.1007/s40257-020-00546-8 (accessed on 11 February 2021).
- Murillo, N.; Aubert, J.; Raoult, D. Microbiota of Demodex Mites from Rosacea Patients and Controls. Microb. Pathog. 2014, 71–72, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Takahashi, M.; Matsutani, M.; Takada, N.; Noda, S.; Saijo, M. Obligate Intracellular Bacteria Diversity in Unfed Leptotrombidium scutellare Larvae Highlights Novel Bacterial Endosymbionts of Mites. Microbiol. Immunol. 2020, 64, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The Tick Microbiome: Why Non-Pathogenic Microorganisms Matter in Tick Biology and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 236. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, K.; Sielawa, H.; Chen, W.; Dzika, E. Endosymbiosis and Its Significance in Dermatology. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Clanner-Engelshofen, B.M.; French, L.E.; Reinholz, M. Corynebacterium kroppenstedtii Subsp. demodicis is the Endobacterium of Demodex folliculorum. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1043–1049. [Google Scholar] [CrossRef] [Green Version]
- Ogai, K.; Shibata, K.; Takahashi, N.; Ogura, K.; Okamoto, S.; Sugama, J. Amplicon-Based Skin Microbiome Profiles Collected by Tape Stripping with Different Adhesive Film Dressings: A Comparative Study. BMC Microbiol. 2021, 21, 54. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region as a Universal DNA Barcode Marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- De Filippis, F.; Laiola, M.; Blaiotta, G.; Ercolini, D. Different Amplicon Targets for Sequencing-Based Studies of Fungal Diversity. Appl. Environ. Microbiol. 2017, 83, e00905-17. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Freeman, A.F.; NISC Comparative Sequencing Program; Park, M.; Sokolic, R.; Candotti, F.; Holland, S.M.; Segre, J.A.; Kong, H.H. The Altered Landscape of the Human Skin Microbiome in Patients with Primary Immunodeficiencies. Genome Res. 2013, 23, 2103–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, E.; Tanaka, T.; Tajima, M.; Tsuboi, R.; Nishikawa, A.; Sugita, T. Characterization of the Skin Fungal Microbiota in Patients with Atopic Dermatitis and in Healthy Subjects. Microbiol. Immunol. 2011, 55, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.-P.; Warnow, T.; Pop, M.; White, B. A Perspective on 16S RRNA Operational Taxonomic Unit Clustering Using Sequence Similarity. NPJ Biofilms Microbiomes 2016, 2, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelino, M.; Eyre, S.; Moat, J.; Fox, G.; Martin, P.; Ho, P.; Upton, M.; Barton, A. Optimisation of Methods for Bacterial Skin Microbiome Investigation: Primer Selection and Comparison of the 454 versus MiSeq Platform. BMC Microbiol. 2017, 17, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palkova, L.; Tomova, A.; Repiska, G.; Babinska, K.; Bokor, B.; Mikula, I.; Minarik, G.; Ostatnikova, D.; Soltys, K. Evaluation of 16S RRNA Primer Sets for Characterisation of Microbiota in Paediatric Patients with Autism Spectrum Disorder. Sci. Rep. 2021, 11, 6781. [Google Scholar] [CrossRef]
- Pausan, M.R.; Csorba, C.; Singer, G.; Till, H.; Schöpf, V.; Santigli, E.; Klug, B.; Högenauer, C.; Blohs, M.; Moissl-Eichinger, C. Exploring the Archaeome: Detection of Archaeal Signatures in the Human Body. Front. Microbiol. 2019, 10, 2796. [Google Scholar] [CrossRef] [Green Version]
- Cococcioni, L.; Panelli, S.; Varotto-Boccazzi, I.; Carlo, D.D.; Pistone, D.; Leccese, G.; Zuccotti, G.V.; Comandatore, F. IBDs and the Pediatric Age: Their Peculiarities and the Involvement of the Microbiota. Dig. Liver Dis. 2021, 53, 17–25. [Google Scholar] [CrossRef]
- de Abreu, V.A.C.; Perdigão, J.; Almeida, S. Metagenomic Approaches to Analyze Antimicrobial Resistance: An Overview. Front. Genet. 2021, 11, 575592. [Google Scholar] [CrossRef]
- Cuscó, A.; Catozzi, C.; Viñes, J.; Sanchez, A.; Francino, O. Microbiota Profiling with Long Amplicons Using Nanopore Sequencing: Full-Length 16S RRNA Gene and the 16S-ITS-23S of the Rrn Operon. F1000Research 2019, 7, 1755. [Google Scholar] [CrossRef] [PubMed]
- Ciuffreda, L.; Rodríguez-Pérez, H.; Flores, C. Nanopore Sequencing and Its Application to the Study of Microbial Communities. Comput. Struct. Biotechnol. J. 2021, 19, 1497–1511. [Google Scholar] [CrossRef] [PubMed]
- Sevim, V.; Lee, J.; Egan, R.; Clum, A.; Hundley, H.; Lee, J.; Everroad, R.C.; Detweiler, A.M.; Bebout, B.M.; Pett-Ridge, J.; et al. Shotgun Metagenome Data of a Defined Mock Community Using Oxford Nanopore, PacBio and Illumina Technologies. Sci. Data 2019, 6, 285. [Google Scholar] [CrossRef] [Green Version]
- Toma, I.; Siegel, M.O.; Keiser, J.; Yakovleva, A.; Kim, A.; Davenport, L.; Devaney, J.; Hoffman, E.P.; Alsubail, R.; Crandall, K.A.; et al. Single-Molecule Long-Read 16S Sequencing to Characterize the Lung Microbiome from Mechanically Ventilated Patients with Suspected Pneumonia. J. Clin. Microbiol. 2014, 52, 3913–3921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, J.A.; Pan, Y.; Tooming-Klunderud, A.; Eijsink, V.G.H.; McHardy, A.C.; Nederbragt, A.J.; Pope, P.B. Improved Metagenome Assemblies and Taxonomic Binning Using Long-Read Circular Consensus Sequence Data. Sci. Rep. 2016, 6, 25373. [Google Scholar] [CrossRef] [Green Version]
- Brede, M.; Orton, T.; Pinior, B.; Roch, F.-F.; Dzieciol, M.; Zwirzitz, B.; Wagner, M.; Breves, G.; Wetzels, S.U. PacBio and Illumina MiSeq Amplicon Sequencing Confirm Full Recovery of the Bacterial Community After Subacute Ruminal Acidosis Challenge in the RUSITEC System. Front. Microbiol. 2020, 11, 1813. [Google Scholar] [CrossRef]
- Mahmoud, M.; Zywicki, M.; Twardowski, T.; Karlowski, W.M. Efficiency of PacBio Long Read Correction by 2nd Generation Illumina Sequencing. Genomics 2019, 111, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Dohm, J.C.; Peters, P.; Stralis-Pavese, N.; Himmelbauer, H. Benchmarking of Long-Read Correction Methods. NAR Genom. Bioinform. 2020, 2, lqaa037. [Google Scholar] [CrossRef] [PubMed]
- Lagier, J.-C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Levasseur, A.; Rolain, J.-M.; Fournier, P.-E.; Raoult, D. Culturing the Human Microbiota and Culturomics. Nat. Rev. Microbiol. 2018, 16, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Lagier, J.-C.; Edouard, S.; Pagnier, I.; Mediannikov, O.; Drancourt, M.; Raoult, D. Current and Past Strategies for Bacterial Culture in Clinical Microbiology. Clin. Microbiol. Rev. 2015, 28, 208–236. [Google Scholar] [CrossRef] [Green Version]
- Lagier, J.-C.; Khelaifia, S.; Alou, M.T.; Ndongo, S.; Dione, N.; Hugon, P.; Caputo, A.; Cadoret, F.; Traore, S.I.; Seck, E.H.; et al. Culture of Previously Uncultured Members of the Human Gut Microbiota by Culturomics. Nat. Microbiol. 2016, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Szymański, Ł.; Jęderka, K.; Cios, A.; Ciepelak, M.; Lewicka, A.; Stankiewicz, W.; Lewicki, S. A Simple Method for the Production of Human Skin Equivalent in 3D, Multi-Cell Culture. Int. J. Mol. Sci. 2020, 21, 4644. [Google Scholar] [CrossRef]
- Smandri, A.; Nordin, A.; Hwei, N.M.; Chin, K.-Y.; Abd Aziz, I.; Fauzi, M.B. Natural 3D-Printed Bioinks for Skin Regeneration and Wound Healing: A Systematic Review. Polymers 2020, 12, 1782. [Google Scholar] [CrossRef]
- Bjerre, R.D.; Hugerth, L.W.; Boulund, F.; Seifert, M.; Johansen, J.D.; Engstrand, L. Effects of Sampling Strategy and DNA Extraction on Human Skin Microbiome Investigations. Sci. Rep. 2019, 9, 17287. [Google Scholar] [CrossRef] [Green Version]
- Boulesnane, Y.; Leloup, J.; Lerch, T.Z.; Roynette, A.; Pensé-Lhéritier, A.-M.; Mielcarek, C.; Changey, F. Impact of Sampling and DNA Extraction Methods on Skin Microbiota Assessment. J. Microbiol. Methods 2020, 171, 105880. [Google Scholar] [CrossRef]
- Manus, M.B.; Kuthyar, S.; Perroni-Marañón, A.G.; la Mora, A.N.; Amato, K.R. Comparing Different Sample Collection and Storage Methods for Field-Based Skin Microbiome Research. Am. J. Hum. Biol. 2021, e23584. [Google Scholar] [CrossRef]
- Ogai, K.; Nagase, S.; Mukai, K.; Iuchi, T.; Mori, Y.; Matsue, M.; Sugitani, K.; Sugama, J.; Okamoto, S. A Comparison of Techniques for Collecting Skin Microbiome Samples: Swabbing Versus Tape-Stripping. Front. Microbiol. 2018, 9, 2362. [Google Scholar] [CrossRef] [PubMed]
- Dréno, B.; Martin, R.; Moyal, D.; Henley, J.B.; Khammari, A.; Seité, S. Skin Microbiome and Acne Vulgaris: Staphylococcus, a New Actor in Acne. Exp. Dermatol. 2017, 26, 798–803. [Google Scholar] [CrossRef] [Green Version]
- Churiso, G.; van Henten, S.; Cnops, L.; Pollmann, J.; Melkamu, R.; Lemma, M.; Kiflie, A.; Fikre, H.; van Griensven, J.; Adriaensen, W. Minimally Invasive Microbiopsies as an Improved Sampling Method for the Diagnosis of Cutaneous Leishmaniasis. Open Forum Infect. Dis. 2020, 7, ofaa364. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Maibach, H.I. Why Minimally Invasive Skin Sampling Techniques? A Bright Scientific Future. Cutan Ocul. Toxicol. 2011, 30, 1–6. [Google Scholar] [CrossRef]
- Prast-Nielsen, S.; Tobin, A.-M.; Adamzik, K.; Powles, A.; Hugerth, L.W.; Sweeney, C.; Kirby, B.; Engstrand, L.; Fry, L. Investigation of the Skin Microbiome: Swabs vs. Biopsies. Br. J. Dermatol. 2019, 181, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.A.; Connolly, J.; Hourihane, J.O.; Fallon, P.G.; McLean, W.H.I.; Murray, D.; Jo, J.-H.; Segre, J.A.; Kong, H.H.; Irvine, A.D. Skin Microbiome before Development of Atopic Dermatitis: Early Colonization with Commensal Staphylococci at 2 Months Is Associated with a Lower Risk of Atopic Dermatitis at 1 Year. J. Allergy Clin. Immunol. 2017, 139, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Shi, B.; Bangayan, N.J.; Curd, E.; Taylor, P.A.; Gallo, R.L.; Leung, D.Y.M.; Li, H. The Skin Microbiome is different in Pediatric versus Adult Atopic Dermatitis. J. Allergy Clin. Immunol. 2016, 138, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D.; Lee, K.; Afifi, L.; Fadrosh, D.; Leech, J.; et al. Alteration of the Cutaneous Microbiome in Psoriasis and Potential Role in Th17 Polarization. Microbiome 2018, 6, 154. [Google Scholar] [CrossRef]
- Drago, L.; De Grandi, R.; Altomare, G.; Pigatto, P.; Rossi, O.; Toscano, M. Skin Microbiota of First Cousins Affected by Psoriasis and Atopic Dermatitis. Clin. Mol. Allergy 2016, 14, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, C.; Chen, X.-Y.; Li, X.; Xue, F.; Chen, L.-H.; Liu, N.; Wang, B.; Wang, L.-Q.; Wang, X.-P.; Yang, H.; et al. Psoriatic Lesions Are Characterized by Higher Bacterial Load and Imbalance between Cutibacterium and Corynebacterium. J. Am. Acad. Dermatol. 2020, 82, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Fahlén, A.; Engstrand, L.; Baker, B.S.; Powles, A.; Fry, L. Comparison of Bacterial Microbiota in Skin Biopsies from Normal and Psoriatic Skin. Arch. Dermatol. Res. 2012, 304, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Forton, F.; Seys, B. Density of Demodex Folliculorum in Rosacea: A Case-Control Study Using Standardized Skin-Surface Biopsy. Br. J. Dermatol. 1993, 128, 650–659. [Google Scholar] [CrossRef]
- van Rensburg, J.J.; Lin, H.; Gao, X.; Toh, E.; Fortney, K.R.; Ellinger, S.; Zwickl, B.; Janowicz, D.M.; Katz, B.P.; Nelson, D.E.; et al. The Human Skin Microbiome Associates with the Outcome of and Is Influenced by Bacterial Infection. mBio 2015, 6, e01315-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, S.; Bouladoux, N.; Linehan, J.L.; Han, S.-J.; Harrison, O.J.; Wilhelm, C.; Conlan, S.; Himmelfarb, S.; Byrd, A.L.; Deming, C.; et al. Commensal–Dendritic-Cell Interaction Specifies a Unique Protective Skin Immune Signature. Nature 2015, 520, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Nakamizo, S.; Egawa, G.; Honda, T.; Nakajima, S.; Belkaid, Y.; Kabashima, K. Commensal Bacteria and Cutaneous Immunity. Semin. Immunopathol. 2015, 37, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Cogen, A.L.; Yamasaki, K.; Sanchez, K.M.; Dorschner, R.A.; Lai, Y.; MacLeod, D.T.; Torpey, J.W.; Otto, M.; Nizet, V.; Kim, J.E.; et al. Selective Antimicrobial Action Is Provided by Phenol-Soluble Modulins Derived from Staphylococcus Epidermidis, a Normal Resident of the Skin. J. Investig. Dermatol. 2010, 130, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanke, I.; Steffen, H.; Christ, C.; Krismer, B.; Götz, F.; Peschel, A.; Schaller, M.; Schittek, B. Skin Commensals Amplify the Innate Immune Response to Pathogens by Activation of Distinct Signaling Pathways. J. Investig. Dermatol. 2011, 131, 382–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callewaert, C.; Knödlseder, N.; Karoglan, A.; Güell, M.; Paetzold, B. Skin Microbiome Transplantation and Manipulation: Current State of the Art. Comput. Struct. Biotechnol. J. 2021, 19, 624–631. [Google Scholar] [CrossRef]
- Kober, M.-M.; Bowe, W.P. The Effect of Probiotics on Immune Regulation, Acne, and Photoaging. Int. J. Womens Dermatol. 2015, 1, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Lolou, V.; Panayiotidis, M.I. Functional Role of Probiotics and Prebiotics on Skin Health and Disease. Fermentation 2019, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Kimoto-Nira, H.; Aoki, R.; Sasaki, K.; Suzuki, C.; Mizumachi, K. Oral Intake of Heat-Killed Cells of Lactococcus lactis Strain H61 Promotes Skin Health in Women. J. Nutr. Sci. 2012, 1, e18. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.E.; Huh, C.S.; Ra, J.; Choi, I.D.; Jeong, J.W.; Kim, S.H.; Ryu, J.H.; Seo, Y.K.; Koh, J.S.; Lee, J.H.; et al. Clinical Evidence of Effects of Lactobacillus Plantarum HY7714 on Skin Aging: A Randomized, Double Blind, Placebo-Controlled Study. J. Microbiol. Biotechnol. 2015, 25, 2160–2168. [Google Scholar] [CrossRef]
- Drago, L.; Meroni, G.; Pistone, D.; Pasquale, L.; Milazzo, G.; Monica, F.; Aragona, S.; Ficano, L.; Vassallo, R.; Gastrobiota Group. Evaluation of Main Functional Dyspepsia Symptoms after Probiotic Administration in Patients Receiving Conventional Pharmacological Therapies. J. Int. Med. Res. 2021, 49. [Google Scholar] [CrossRef]
- Sfriso, R.; Egert, M.; Gempeler, M.; Voegeli, R.; Campiche, R. Revealing the Secret Life of Skin-with the Microbiome You Never Walk Alone. Int. J. Cosmet. Sci. 2020, 42, 116–126. [Google Scholar] [CrossRef]
- Rather, I.A.; Bajpai, V.K.; Kumar, S.; Lim, J.; Paek, W.K.; Park, Y.-H. Probiotics and Atopic Dermatitis: An Overview. Front. Microbiol. 2016, 7, 507. [Google Scholar] [CrossRef] [Green Version]
- Betsi, G.I.; Papadavid, E.; Falagas, M.E. Probiotics for the Treatment or Prevention of Atopic Dermatitis: A Review of the Evidence from Randomized Controlled Trials. Am. J. Clin. Dermatol. 2008, 9, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Foisy, M.; Boyle, R.J.; Chalmers, J.R.; Simpson, E.L.; Williams, H.C. Overview of Reviews The Prevention of Eczema in Infants and Children: An Overview of Cochrane and Non-Cochrane Reviews. Evid. Based Child. Health 2011, 6, 1322–1339. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-O.; Ah, Y.-M.; Yu, Y.M.; Choi, K.H.; Shin, W.-G.; Lee, J.-Y. Effects of Probiotics for the Treatment of Atopic Dermatitis: A Meta-Analysis of Randomized Controlled Trials. Ann. Allergy Asthma Immunol. 2014, 113, 217–226. [Google Scholar] [CrossRef]
- Lee, J.; Seto, D.; Bielory, L. Meta-Analysis of Clinical Trials of Probiotics for Prevention and Treatment of Pediatric Atopic Dermatitis. J. Allergy Clin. Immunol. 2008, 121, 116–121.e11. [Google Scholar] [CrossRef] [PubMed]
- West, C.E.; Jenmalm, M.C.; Prescott, S.L. The Gut Microbiota and Its Role in the Development of Allergic Disease: A Wider Perspective. Clin. Exp. Allergy 2015, 45, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestito, S.; D’Auria, E.; Baldassarre, M.E.; Salvatore, S.; Tallarico, V.; Stefanelli, E.; Tarsitano, F.; Concolino, D.; Pensabene, L. The Role of Prebiotics and Probiotics in Prevention of Allergic Diseases in Infants. Front. Pediatr. 2020, 8, 583946. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Trivedi, M.K.; Jha, A.; Lin, Y.-F.; Dimaano, L.; García-Romero, M.T. Synbiotics for Prevention and Treatment of Atopic Dermatitis: A Meta-Analysis of Randomized Clinical Trials. JAMA Pediatr. 2016, 170, 236–242. [Google Scholar] [CrossRef]
- Makrgeorgou, A.; Leonardi-Bee, J.; Bath-Hextall, F.J.; Murrell, D.F.; Tang, M.L.; Roberts, A.; Boyle, R.J. Probiotics for Treating Eczema. Cochrane Database Syst. Rev 2018, 11, CD006135. [Google Scholar] [CrossRef]
- Groeger, D.; O’Mahony, L.; Murphy, E.F.; Bourke, J.F.; Dinan, T.G.; Kiely, B.; Shanahan, F.; Quigley, E.M.M. Bifidobacterium Infantis 35624 Modulates Host Inflammatory Processes beyond the Gut. Gut Microbes 2013, 4, 325–339. [Google Scholar] [CrossRef] [Green Version]
- Navarro-López, V.; Martínez-Andrés, A.; Ramírez-Boscá, A.; Ruzafa-Costas, B.; Núñez-Delegido, E.; Carrión-Gutiérrez, M.A.; Prieto-Merino, D.; Codoñer-Cortés, F.; Ramón-Vidal, D.; Genovés-Martínez, S.; et al. Efficacy and Safety of Oral Administration of a Mixture of Probiotic Strains in Patients with Psoriasis: A Randomized Controlled Clinical Trial. Acta Derm. Venereol. 2019, 99, 1078–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Immune-Mediated Mechanisms of Action of Probiotics and Synbiotics in Treating Pediatric Intestinal Diseases. Nutrients 2018, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaza-Díaz, J.; Robles-Sánchez, C.; Abadía-Molina, F.; Sáez-Lara, M.J.; Vilchez-Padial, L.M.; Gil, Á.; Gómez-Llorente, C.; Fontana, L. Gene Expression Profiling in the Intestinal Mucosa of Obese Rats Administered Probiotic Bacteria. Sci. Data 2017, 4, 170186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and Postbiotic Activity in Health and Disease: Comparison on a Novel Polarised Ex-Vivo Organ Culture Model. Gut 2012, 61, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Moroi, M.; Uchi, S.; Nakamura, K.; Sato, S.; Shimizu, N.; Fujii, M.; Kumagai, T.; Saito, M.; Uchiyama, K.; Watanabe, T.; et al. Beneficial Effect of a Diet Containing Heat-Killed Lactobacillus Paracasei K71 on Adult Type Atopic Dermatitis. J. Dermatol. 2011, 38, 131–139. [Google Scholar] [CrossRef]
- D’Auria, E.; Panelli, S.; Lunardon, L.; Pajoro, M.; Paradiso, L.; Beretta, S.; Loretelli, C.; Tosi, D.; Perini, M.; Bedogni, G.; et al. Rice Flour Fermented with Lactobacillus Paracasei CBA L74 in the Treatment of Atopic Dermatitis in Infants: A Randomized, Double- Blind, Placebo- Controlled Trial. Pharmacol. Res. 2021, 163, 105284. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.-C.; Hung, C.-H.; Sy, L.B.; Lue, K.-H.; Shih, I.-H.; Yang, C.-Y.; Chen, L.-C.; Sun, H.-L.; Lee, M.-S.; Chambard, J.; et al. A Randomized, Double-Blind, Placebo-Controlled Trial Assessing the Oral Administration of a Heat-Treated Lactobacillus Paracasei Supplement in Infants with Atopic Dermatitis Receiving Topical Corticosteroid Therapy. Skin Pharmacol. Physiol. 2019, 32, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, D.R.; Feelisch, M. Soil bacteria, nitrite and the skin. In The Hygiene Hypothesis and Darwinian Medicine; Rook, G.A.W., Ed.; Progress in Inflammation Research; Birkhäuser: Basel, Switzerland, 2009; pp. 103–115. ISBN 978-3-7643-8903-1. [Google Scholar]
- Lee, N.Y.; Ibrahim, O.; Khetarpal, S.; Gaber, M.; Jamas, S.; Gryllos, I.; Dover, J.S. Dermal Microflora Restoration With Ammonia-Oxidizing Bacteria Nitrosomonas Eutropha in the Treatment of Keratosis Pilaris: A Randomized Clinical Trial. J. Drugs Dermatol. 2018, 17, 285–288. [Google Scholar]
- Navarro-López, V.; Núñez-Delegido, E.; Ruzafa-Costas, B.; Sánchez-Pellicer, P.; Agüera-Santos, J.; Navarro-Moratalla, L. Probiotics in the Therapeutic Arsenal of Dermatologists. Microorganisms 2021, 9, 1513. [Google Scholar] [CrossRef]
- Notay, M.; Saric-Bosanac, S.; Vaughn, A.R.; Dhaliwal, S.; Trivedi, M.; Reiter, P.N.; Rybak, I.; Li, C.C.; Weiss, L.B.; Ambrogio, L.; et al. The Use of Topical Nitrosomonas Eutropha for Cosmetic Improvement of Facial Wrinkles. J. Cosmet. Dermatol. 2020, 19, 689–693. [Google Scholar] [CrossRef]
- Maura, D.; Elmekki, N.; Goddard, C.A. The Ammonia Oxidizing Bacterium Nitrosomonas Eutropha Blocks T Helper 2 Cell Polarization via the Anti-Inflammatory Cytokine IL-10. Sci. Rep. 2021, 11, 14162. [Google Scholar] [CrossRef] [PubMed]
- Leonel, C.; Sena, I.F.G.; Silva, W.N.; Prazeres, P.H.D.M.; Fernandes, G.R.; Agresti, P.M.; Drumond, M.M.; Mintz, A.; Azevedo, V.A.C.; Birbrair, A. Staphylococcus Epidermidis Role in the Skin Microenvironment. J. Cell. Mol. Med. 2019, 23, 5949–5955. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.M.; Freire, M.O.; Gabrilska, R.A.; Rumbaugh, K.P.; Lemon, K.P. Staphylococcus Aureus Shifts toward Commensalism in Response to Corynebacterium Species. Front. Microbiol. 2016, 7, 1230. [Google Scholar] [CrossRef] [Green Version]
- Hendricks, A.J.; Mills, B.W.; Shi, V.Y. Skin Bacterial Transplant in Atopic Dermatitis: Knowns, Unknowns and Emerging Trends. J. Dermatol. Sci. 2019, 95, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knackstedt, R.; Knackstedt, T.; Gatherwright, J. The Role of Topical Probiotics in Skin Conditions: A Systematic Review of Animal and Human Studies and Implications for Future Therapies. Exp. Dermatol. 2020, 29, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Wan, P.; Chen, J. A Calm, Dispassionate Look at Skin Microbiota in Atopic Dermatitis: An Integrative Literature Review. Dermatol. Ther. (Heidelb.) 2020, 10, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsuji, T.; Hata, T.R.; Tong, Y.; Cheng, J.Y.; Shafiq, F.; Butcher, A.M.; Salem, S.S.; Brinton, S.L.; Spergel, A.K.R.; Johnson, K.; et al. Development of a Human Skin Commensal Microbe for Bacteriotherapy of Atopic Dermatitis and Use in a Phase 1 Randomized Clinical Trial. Nat. Med. 2021, 27, 700–709. [Google Scholar] [CrossRef]
- Di Marzio, L.; Centi, C.; Cinque, B.; Masci, S.; Giuliani, M.; Arcieri, A.; Zicari, L.; De Simone, C.; Cifone, M.G. Effect of the Lactic Acid Bacterium Streptococcus Thermophilus on Stratum Corneum Ceramide Levels and Signs and Symptoms of Atopic Dermatitis Patients. Exp. Dermatol. 2003, 12, 615–620. [Google Scholar] [CrossRef]
- Gueniche, A.; Knaudt, B.; Schuck, E.; Volz, T.; Bastien, P.; Martin, R.; Röcken, M.; Breton, L.; Biedermann, T. Effects of Nonpathogenic Gram-Negative Bacterium Vitreoscilla Filiformis Lysate on Atopic Dermatitis: A Prospective, Randomized, Double-Blind, Placebo-Controlled Clinical Study. Br. J. Dermatol. 2008, 159, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Myles, I.A.; Earland, N.J.; Anderson, E.D.; Moore, I.N.; Kieh, M.D.; Williams, K.W.; Saleem, A.; Fontecilla, N.M.; Welch, P.A.; Darnell, D.A.; et al. First-in-Human Topical Microbiome Transplantation with Roseomonas Mucosa for Atopic Dermatitis. JCI Insight 2018, 3, 120608. [Google Scholar] [CrossRef] [Green Version]
- Kimura, K.; Hagiya, H.; Nishi, I.; Yoshida, H.; Tomono, K. Roseomonas Mucosa Bacteremia in a Neutropenic Child: A Case Report and Literature Review. IDCases 2018, 14, e00469. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Ayibieke, A.; Saito, R.; Ogura, K.; Magara, Y.; Ueda, R.; Ogawa, H.; Hatakeyama, S. A Nosocomial Cluster of Roseomonas Mucosa Bacteremia Possibly Linked to Contaminated Hospital Environment. J. Infect. Chemother. 2020, 26, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Guo, X.; Guo, P.; Cui, Y.; Chen, Y. Roseomonas Mucosa Infective Endocarditis in Patient with Systemic Lupus Erythematosus: Case Report and Review of Literature. BMC Infect. Dis. 2019, 19, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, H.; Blanco, D.; McKeever, C.; Paz, N.; Vasquez, Y.N.; Quiring, J.; Enloe, C.; De León, E.; Stasko, N. Results of a Phase 2 Efficacy and Safety Study with SB204, an Investigational Topical Nitric Oxide-Releasing Drug for the Treatment of Acne Vulgaris. J. Clin. Aesthet. Dermatol. 2016, 9, 12–18. [Google Scholar]
- Kim, M.-J.; Kim, K.-P.; Choi, E.; Yim, J.-H.; Choi, C.; Yun, H.-S.; Ahn, H.-Y.; Oh, J.-Y.; Cho, Y. Effects of Lactobacillus Plantarum CJLP55 on Clinical Improvement, Skin Condition and Urine Bacterial Extracellular Vesicles in Patients with Acne Vulgaris: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2021, 13, 1368. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kuo, S.; Shu, M.; Yu, J.; Huang, S.; Dai, A.; Two, A.; Gallo, R.L.; Huang, C.-M. Staphylococcus Epidermidis in the Human Skin Microbiome Mediates Fermentation to Inhibit the Growth of Propionibacterium Acnes: Implications of Probiotics in Acne Vulgaris. Appl. Microbiol. Biotechnol. 2014, 98, 411–424. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.L.; Petrovski, S.; Dyson, Z.A.; Seviour, R.; Tucci, J. The Formulation of Bacteriophage in a Semi Solid Preparation for Control of Propionibacterium Acnes Growth. PLoS ONE 2016, 11, e0151184. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.R.; Maarouf, M.; Hendricks, A.J.; Lee, D.E.; Shi, V.Y. Topical Probiotics: The Unknowns behind Their Rising Popularity. Dermatol. Online J. 2019, 25. [Google Scholar] [CrossRef]
- Timm, C.M.; Loomis, K.; Stone, W.; Mehoke, T.; Brensinger, B.; Pellicore, M.; Staniczenko, P.P.A.; Charles, C.; Nayak, S.; Karig, D.K. Isolation and Characterization of Diverse Microbial Representatives from the Human Skin Microbiome. Microbiome 2020, 8, 58. [Google Scholar] [CrossRef]
- Uberoi, A.; Bartow-McKenney, C.; Zheng, Q.; Flowers, L.; Campbell, A.; Knight, S.A.B.; Chan, N.; Wei, M.; Lovins, V.; Bugayev, J.; et al. Commensal Microbiota Regulates Skin Barrier Function and Repair via Signaling through the Aryl Hydrocarbon Receptor. Cell Host Microbe 2021, 29, 1235–1248.e8. [Google Scholar] [CrossRef]
- Coates, M.; Lee, M.J.; Norton, D.; MacLeod, A.S. The Skin and Intestinal Microbiota and Their Specific Innate Immune Systems. Front. Immunol. 2019, 10, 2950. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The Gut-Skin Axis in Health and Disease: A Paradigm with Therapeutic Implications. BioEssays 2016, 38, 1167–1176. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Zhou, N.; McDonald, D.; Costello, E.K.; Knight, R. Forensic Identification Using Skin Bacterial Communities. Proc. Natl. Acad. Sci. USA 2010, 107, 6477–6481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meadow, J.F.; Altrichter, A.E.; Green, J.L. Mobile Phones Carry the Personal Microbiome of Their Owners. PeerJ 2014, 2, e447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szántó, M.; Dózsa, A.; Antal, D.; Szabó, K.; Kemény, L.; Bai, P. Targeting the Gut-Skin Axis—Probiotics as New Tools for Skin Disorder Management? Exp. Dermatol. 2019, 28, 1210–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Skin Disease | Microorganisms | References | Other Factors Implicated in the Pathology Moderately Associated (•) Highly Associated (•••) |
|---|---|---|---|
| Rosacea | Demodex folliculorum ↑ | [84,85,86,87,88,89] | Microbiome composition ••• |
| Helicobacter pylori ↑ | [90,91] | Solar exposure • | |
| Staphylococcus epidermidis ↑ | [86,92] | Dietary agents • | |
| Chlamydophyla pneumoniae ↑ | [93,94] | Drugs • | |
| Bacillus oleronius ↑ | [86] | Abnormalities of the cutaneous vascular and lymphatic system • | |
| Enhanced expression of toll-like receptor 2 in the epidermis and amplified inflammatory response ••• | |||
| Abnormalities of the sebaceous gland • | |||
| Dermal matrix degeneration • | |||
| Atopic dermatitis | Staphylococcus aureus ↑ | [95,96,97,98,99] | Food allergies • |
| herpes simplex virus ↑ | [100,101] | Irritants in contact with skin • (clothes, detergents, jewelry, ets) | |
| Staphylococcus epidermidis ↓ CoNS | [100] | Hormonal changes • | |
| Decrease in antimicrobial peptides • | |||
| Increased skin pH • | |||
| Th2 (cytokines such as IL-4 and IL-13) • | |||
| Psoriasis | Staphylococcus aureus ↑ | [102,103,104,105,106,107] | Genetics ••• |
| Streptococcus pyogenes ↑ | secondary colonization? | Hormonal changes • | |
| Human papillomavirus and endogenous retroviruses ↑ | Immune disorders • | ||
| Malassezia spp. ↑ | Alcohol consumption • | ||
| Candida albicans ↑ | Smoking • | ||
| Propionibacterium spp. ↓ | [105,106] | Stress • | |
| Acne vulgaris | Cutibacterium acnes ↑ | [108,109] | Hormonal changes ••• |
| Malassezia spp. ↑ | [110] | Medications (e.g. corticosteroids, lithium) • | |
| Diet • | |||
| Stress • |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pistone, D.; Meroni, G.; Panelli, S.; D’Auria, E.; Acunzo, M.; Pasala, A.R.; Zuccotti, G.V.; Bandi, C.; Drago, L. A Journey on the Skin Microbiome: Pitfalls and Opportunities. Int. J. Mol. Sci. 2021, 22, 9846. https://doi.org/10.3390/ijms22189846
Pistone D, Meroni G, Panelli S, D’Auria E, Acunzo M, Pasala AR, Zuccotti GV, Bandi C, Drago L. A Journey on the Skin Microbiome: Pitfalls and Opportunities. International Journal of Molecular Sciences. 2021; 22(18):9846. https://doi.org/10.3390/ijms22189846
Chicago/Turabian StylePistone, Dario, Gabriele Meroni, Simona Panelli, Enza D’Auria, Miriam Acunzo, Ajay Ratan Pasala, Gian Vincenzo Zuccotti, Claudio Bandi, and Lorenzo Drago. 2021. "A Journey on the Skin Microbiome: Pitfalls and Opportunities" International Journal of Molecular Sciences 22, no. 18: 9846. https://doi.org/10.3390/ijms22189846
APA StylePistone, D., Meroni, G., Panelli, S., D’Auria, E., Acunzo, M., Pasala, A. R., Zuccotti, G. V., Bandi, C., & Drago, L. (2021). A Journey on the Skin Microbiome: Pitfalls and Opportunities. International Journal of Molecular Sciences, 22(18), 9846. https://doi.org/10.3390/ijms22189846