Sweet Basil Has Distinct Synthases for Eugenol Biosynthesis in Glandular Trichomes and Roots with Different Regulatory Mechanisms

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
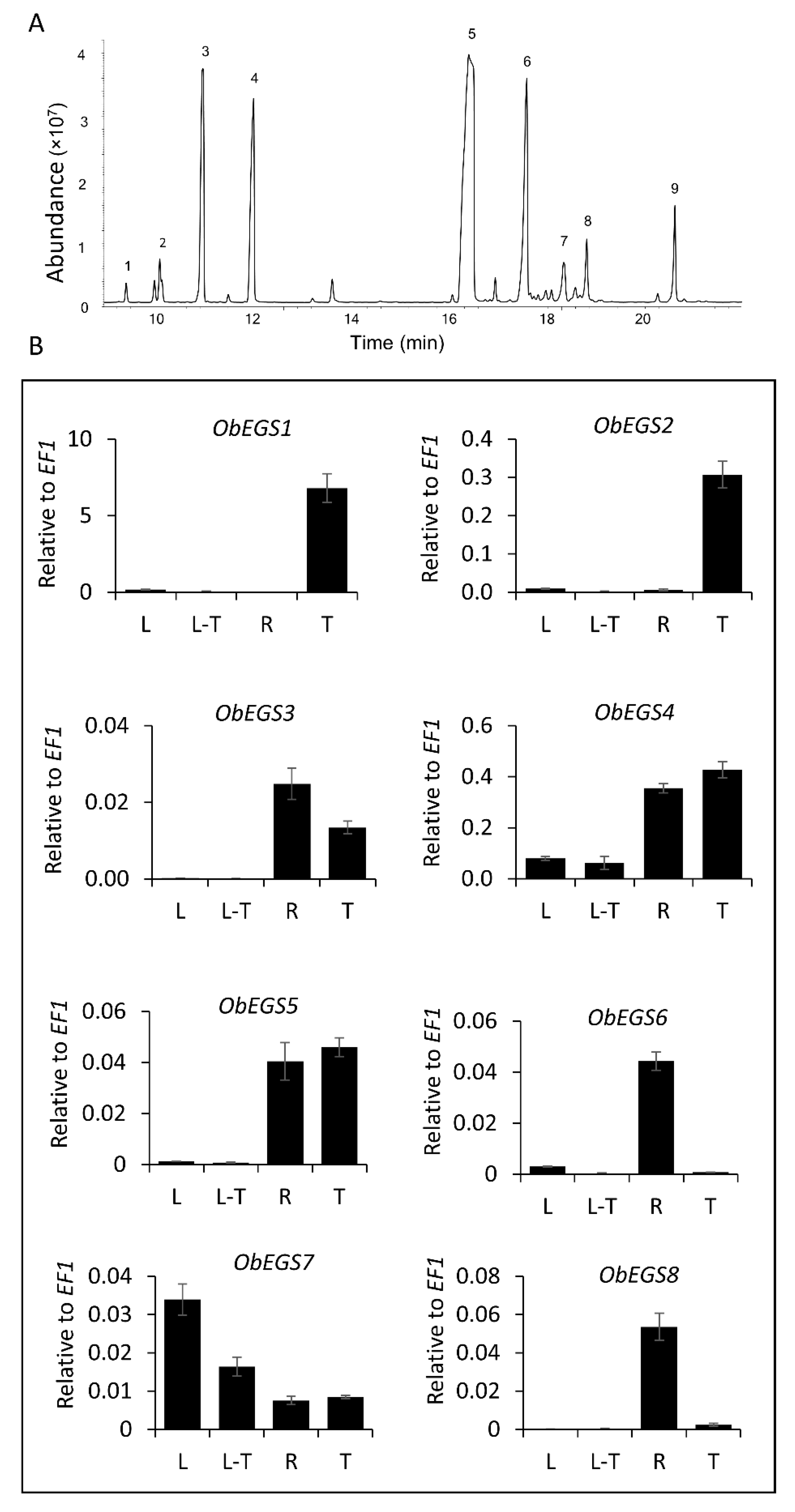
2.1. Identification of Eight Distinct Differentially Expressed PIP Family Reductases from Sweet Basil
2.2. Functional Characterization of EGSs
2.3. Transient Expression of EGSs in Tobacco Plants
2.4. Analysis of ObEGS1 and ObEGS4 Promoter Shows the Presence of Different Cis Regulatory Elements
2.5. Eugenol Was Not Observed in Roots of Aseptically-Grown Plants but Could Be Induced by Elicitors
2.6. PTMs Can Regulate Eugenol Synthase in the Roots of Sweet Basil
3. Discussion
4. Materials and Methods
4.1. Plant Material and RNA Isolation
4.2. Gene Amplification and Plasmid Construction
4.3. Quantitative Real Time PCR (qRT-PCR)
4.4. Subcellular Localization of ObEGSs
4.5. Promoter Analysis
4.6. Histology
4.7. Sweet Basil Transformation
4.8. In Vivo Assays in E. coli and Tobacco Leaves
4.9. Elicitor Treatment
4.10. GC-MS Analysis
4.11. HPLC Analysis
4.12. Total Protein Isolation
4.13. Phosphoprotein Enrichment and Ubiquitin Enrichment
4.14. Western Blot Analysis
4.15. Phylogenetic Analysis
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Ninkovic, V.; Rensing, M.; Dahlin, I.; Markovic, D. Who is my neighbor? Volatile cues in plant interactions. Plant Signal. Behav. 2019, 14, 1634993. [Google Scholar] [CrossRef]
- Selmar, D.; Kleinwächter, M. Stress Enhances the Synthesis of Secondary Plant Products: The Impact of Stress-Related Over-Reduction on the Accumulation of Natural Products. Plant Cell Physiol. 2013, 54, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [PubMed]
- Niinemets, Ü. Uncovering the hidden facets of drought stress: Secondary metabolites make the difference. Tree Physiol. 2015, 36, tpv128. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Griffin, T.S.; Kraner, D.; Schaffner, M.K.; Sharma, D.; Hazel, M.; Leitch, A.R.; Orians, C.M.; Han, W.; Stepp, J.R.; et al. Environmental Factors Variably Impact Tea Secondary Metabolites in the Context of Climate Change. Front. Plant Sci. 2019, 10, 939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, A.; Agrawal, L.; Misra, R.C.; Sharma, S.; Ghosh, S. Andrographis paniculata transcriptome provides molecular insights into tissue-specific accumulation of medicinal diterpenes. BMC Genom. 2015, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Xu, S.; Wang, N.; Xia, B.; Jiang, Y.; Wang, R. Transcriptome Analysis of Secondary Metabolism Pathway, Transcription Factors, and Transporters in Response to Methyl Jasmonate in Lycoris aurea. Front. Plant Sci. 2016, 7, 1971. [Google Scholar] [CrossRef] [Green Version]
- Kleinheinz, G.T.; Bagley, S.T.; John, W.P.S.; Rughani, J.R.; McGinnis, G.D. Characterization of Alpha-Pinene-Degrading Microorganisms and Application to a Bench-Scale Biofiltration System for VOC Degradation. Arch. Environ. Contam. Toxicol. 1999, 37, 151–157. [Google Scholar] [CrossRef]
- Rasmann, S.; Köllner, T.G.; Degenhardt, J.; Hiltpold, I.; Toepfer, S.; Kuhlmann, U.; Gershenzon, J.; Turlings, T.C.J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nat. Cell Biol. 2005, 434, 732–737. [Google Scholar] [CrossRef]
- Robert, C.A.M.; Erb, M.; Duployer, M.; Zwahlen, C.; Doyen, G.R.; Turlings, T.C.J. Herbivore-induced plant volatiles mediate host selection by a root herbivore. New Phytol. 2012, 194, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Wenke, K.; Kai, M.; Piechulla, B. Belowground volatiles facilitate interactions between plant roots and soil organisms. Planta 2010, 231, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Gang, D.R.; Wang, J.; Dudareva, N.; Nam, K.H.; Simon, J.E.; Lewinsohn, E.; Pichersky, E. An Investigation of the Storage and Biosynthesis of Phenylpropenes in Sweet Basil. Plant Physiol. 2001, 125, 539–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deschamps, C.; Simon, J.E. Phenylpropanoid Biosynthesis in Leaves and Glandular Trichomes of Basil (Ocimum basilicum L.). In Plant Secondary Metabolism Engineering. Methods in Molecular Biology (Methods and Protocols); Fett-Neto, A., Ed.; Humana Press: Totowa, NJ, USA, 2010; Volume 643, pp. 263–273. [Google Scholar]
- Lu, D.; Yuan, X.; Kim, S.-J.; Marques, J.V.; Chakravarthy, P.P.; Moinuddin, S.G.A.; Luchterhand, R.; Herman, B.; Davin, L.B.; Lewis, N.G. Eugenol specialty chemical production in transgenic poplar (Populus tremula × P. alba) field trials. Plant Biotechnol. J. 2017, 15, 970–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, T.; Kasahara, H.; Bedgar, D.L.; Youn, B.; Lawrence, P.K.; Gang, D.R.; Halls, S.C.; Park, H.; Hilsenbeck, J.L.; Davin, L.B.; et al. Crystal Structures of Pinoresinol-Lariciresinol and Phenylcoumaran Benzylic Ether Reductases and Their Relationship to Isoflavone Reductases. J. Biol. Chem. 2003, 278, 50714–50723. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; He, X.; Lin, J.; Shao, H.; Chang, Z.; Dixon, R.A. Crystal Structure of Isoflavone Reductase from Alfalfa (Medicago sativa L.). J. Mol. Biol. 2006, 358, 1341–1352. [Google Scholar] [CrossRef]
- Koeduka, T.; Fridman, E.; Gang, D.R.; Vassão, D.G.; Jackson, B.L.; Kish, C.M.; Orlova, I.; Spassova, S.M.; Lewis, N.G.; Noel, J.P.; et al. Eugenol and isoeugenol, characteristic aromatic constituents of spices, are biosynthesized via reduction of a coniferyl alcohol ester. Proc. Natl. Acad. Sci. USA 2006, 103, 10128–10133. [Google Scholar] [CrossRef] [Green Version]
- Dexter, R.; Qualley, A.; Kish, C.M.; Ma, C.J.; Koeduka, T.; Nagegowda, D.A.; Dudareva, N.; Pichersky, E.; Clark, D. Characterization of a petunia acetyltransferase involved in the biosynthesis of the floral volatile isoeugenol. Plant J. 2006, 49, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Dhar, N.; Sarangapani, S.; Reddy, V.A.; Kumar, N.; Panicker, D.; Jin, J.; Chua, N.; Sarojam, R.; Sreelatha, S.; Rajani, S. Characterization of a sweet basil acyltransferase involved in eugenol biosynthesis. J. Exp. Bot. 2020, 71, 3638–3652. [Google Scholar] [CrossRef]
- Gupta, A.K.; Schauvinhold, I.; Pichersky, E.; Schiestl, F.P. Eugenol synthase genes in floral scent variation in Gymnadenia species. Funct. Integr. Genom. 2014, 14, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Koeduka, T.; Louie, G.V.; Orlova, I.; Kish, C.M.; Ibdah, M.; Wilkerson, C.G.; Bowman, M.E.; Baiga, T.J.; Noel, J.P.; Dudareva, N.; et al. The multiple phenylpropene synthases in both Clarkia breweri and Petunia hybrida represent two distinct protein lineages. Plant J. 2008, 54, 362–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Baudino, S.; Caissard, J.-C.; Nicolè, F.; Zhang, H.; Tang, K.; Li, S.; Lu, S. Functional characterization of the eugenol synthase gene (RcEGS1) in rose. Plant Physiol. Biochem. 2018, 129, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Aragüez, I.; Osorio, S.; Hoffmann, T.; Rambla, J.L.; Medina-Escobar, N.; Granell, A.; Botella, J.R.; Schwab, W.; Valpuesta, V. Eugenol Production in Achenes and Receptacles of Strawberry Fruits Is Catalyzed by Synthases Exhibiting Distinct Kinetics. Plant Physiol. 2013, 163, 946–958. [Google Scholar] [CrossRef] [Green Version]
- Yahyaa, M.; Berim, A.; Nawade, B.; Ibdah, M.; Dudareva, N.; Ibdah, M. Biosynthesis of methyleugenol and methylisoeugenol in Daucus carota leaves: Characterization of eugenol/isoeugenol synthase and O-Methyltransferase. Phytochemistry 2019, 159, 179–189. [Google Scholar] [CrossRef]
- Lewis, N.G.; Davin, L.B.; Kim, S.J.; Vassão, D.G.; Patten, A.M.; Eichinger, D. Genes Encoding Chavicol/Eugenol Synthase from the Creosote Bush Larrea Tridentata. U.S. Patent 9,131,648, 15 September 2015. [Google Scholar]
- Anand, A.; Jayaramaiah, R.H.; Beedkar, S.D.; Singh, P.A.; Joshi, R.S.; Mulani, F.A.; Dholakia, B.B.; Punekar, S.A.; Gade, W.N.; Thulasiram, H.V.; et al. Comparative functional characterization of eugenol synthase from four different Ocimum species: Implications on eugenol accumulation. Biochim. Biophys. Acta BBA Proteins Proteom. 2016, 1864, 1539–1547. [Google Scholar] [CrossRef]
- Srivastava, S.; Conlan, X.A.; Adholeya, A.; Cahill, D.M. Elite hairy roots of Ocimum basilicum as a new source of rosmarinic acid and antioxidants. Plant Cell Tissue Organ Cult. PCTOC 2016, 126, 19–32. [Google Scholar] [CrossRef]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Mühlemann, J.K.; Woodworth, B.D.; Morgan, J.A.; Dudareva, N. The monolignol pathway contributes to the biosynthesis of volatile phenylpropenes in flowers. New Phytol. 2014, 204, 661–670. [Google Scholar] [CrossRef]
- Sibout, R.; Höfte, H. Plant Cell Biology: The ABC of Monolignol Transport. Curr. Biol. 2012, 22, R533–R535. [Google Scholar] [CrossRef] [Green Version]
- Prouse, M.B.; Campbell, M.M. The interaction between MYB proteins and their target DNA binding sites. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1819, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Koschmann, J.; Machens, F.; Becker, M.; Niemeyer, J.; Schulze, J.; Bülow, L.; Stahl, D.J.; Hehl, R. Integration of Bioinformatics and Synthetic Promoters Leads to the Discovery of Novel Elicitor-Responsive cis-Regulatory Sequences in Arabidopsis. Plant Physiol. 2012, 160, 178–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Puche, L.; Molina-Hidalgo, F.J.; Boersma, M.; Schuurink, R.C.; López-Vidriero, I.; Solano, R.; Franco-Zorrilla, J.-M.; Caballero, J.L.; Blanco-Portales, R.; García-Limones, C. An R2R3-MYB Transcription Factor Regulates Eugenol Production in Ripe Strawberry Fruit Receptacles. Plant Physiol. 2015, 168, 598–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koeduka, T.; Suzuki, S.; Iijima, Y.; Ohnishi, T.; Suzuki, H.; Watanabe, B.; Shibata, D.; Umezawa, T.; Pichersky, E.; Hiratake, J. Enhancement of production of eugenol and its glycosides in transgenic aspen plants via genetic engineering. Biochem. Biophys. Res. Commun. 2013, 436, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cna’Ani, A.; Shavit, R.; Ravid, J.; Aravena-Calvo, J.; Skaliter, O.; Masci, T.; Vainstein, A. Phenylpropanoid Scent Compounds in Petunia x hybrida Are Glycosylated and Accumulate in Vacuoles. Front. Plant Sci. 2017, 8, 1898. [Google Scholar] [CrossRef] [Green Version]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites1 1Edited by F. E. Cohen. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef]
- Huang, C.-H.; Su, M.-G.; Kao, H.-J.; Jhong, J.-H.; Weng, S.-L.; Lee, T.-Y. UbiSite: Incorporating two-layered machine learning method with substrate motifs to predict ubiquitin-conjugation site on lysines. BMC Syst. Biol. 2016, 10, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Chaieb, K.; Hajlaoui, H.; Zmantar, T.; Ben Kahla-Nakbi, A.; Rouabhia, M.; Mahdouani, K.; Bakhrouf, A. The chemical composition and biological activity of clove essential oil, Eugenia caryophyllata (Syzigium aromaticum L. Myrtaceae): A short review. Phytother. Res. 2007, 21, 501–506. [Google Scholar] [CrossRef]
- DeLory, B.M.; Delaplace, P.; Fauconnier, M.; Du Jardin, P. Root-emitted volatile organic compounds: Can they mediate belowground plant-plant interactions? Plant Soil 2016, 402, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Muchlinski, A.; Chen, X.; Lovell, J.T.; Köllner, T.G.; Pelot, K.A.; Zerbe, P.; Ruggiero, M.; Callaway, L.I.; Laliberte, S.; Chen, F.; et al. Biosynthesis and Emission of Stress-Induced Volatile Terpenes in Roots and Leaves of Switchgrass (Panicum virgatum L.). Front. Plant Sci. 2019, 10, 1144. [Google Scholar] [CrossRef] [Green Version]
- Pozo, M.J.; Van Loon, L.C.; Pieterse, C.M.J. Jasmonates—Signals in plant-microbe interactions. J. Plant Growth Regul. 2004, 23, 211–222. [Google Scholar]
- Ncube, B.; Finnie, J.; Amoo, S.O. Quality from the field: The impact of environmental factors as quality determinants in medicinal plants. S. Afr. J. Bot. 2012, 82, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 2016, 6, 29265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashiguchi, A.; Komatsu, S. Chapter Six—Posttranslational Modifications and Plant–Environment Interaction. In Methods in Enzymology; Shukla, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 97–113. [Google Scholar]
- Xie, Z.; Kapteyn, J.; Gang, D.R. A systems biology investigation of the MEP/terpenoid and shikimate/phenylpropanoid pathways points to multiple levels of metabolic control in sweet basil glandular trichomes. Plant J. 2008, 54, 349–361. [Google Scholar] [CrossRef]
- Wang, J.P.; Chuang, L.; Loziuk, P.L.; Chen, H.; Lin, Y.-C.; Shi, R.; Qu, G.-Z.; Muddiman, D.C.; Sederoff, R.R.; Chiang, V.L. Phosphorylation is an on/off switch for 5-hydroxyconiferaldehyde O-methyltransferase activity in poplar monolignol biosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 8481–8486. [Google Scholar] [CrossRef] [Green Version]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef]
- Jin, J.; Panicker, D.; Wang, Q.; Kim, M.J.; Liu, J.; Yin, J.-L.; Wong, L.; Jang, I.-C.; Chua, N.; Sarojam, R. Next generation sequencing unravels the biosynthetic ability of Spearmint (Mentha spicata) peltate glandular trichomes through comparative transcriptomics. BMC Plant Biol. 2014, 14, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nicot, N.; Hausman, J.-F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, X.; Garreton, V.; Chua, N. The AIP2 E3 ligase acts as a novel negative regulator of ABA signaling by promoting ABI3 degradation. Genes Dev. 2005, 19, 1532–1543. [Google Scholar] [CrossRef] [Green Version]
- Lou, Y.-G.; Du, M.-H.; Turlings, T.C.J.; Cheng, J.-A.; Shan, W.-F. Exogenous Application of Jasmonic Acid Induces Volatile Emissions in Rice and Enhances Parasitism of Nilaparvata lugens Eggs by theParasitoid Anagrus nilaparvatae. J. Chem. Ecol. 2005, 31, 1985–2002. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Cahill, D.M.; Conlan, X.A.; Adholeya, A. A Novel in Vitro Whole Plant System for Analysis of Polyphenolics and Their Antioxidant Potential in Cultivars of Ocimum basilicum. J. Agric. Food Chem. 2014, 62, 10064–10075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.-C.; Mahmood, T. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddy, V.A.; Li, C.; Nadimuthu, K.; Tjhang, J.G.; Jang, I.-C.; Rajani, S. Sweet Basil Has Distinct Synthases for Eugenol Biosynthesis in Glandular Trichomes and Roots with Different Regulatory Mechanisms. Int. J. Mol. Sci. 2021, 22, 681. https://doi.org/10.3390/ijms22020681
Reddy VA, Li C, Nadimuthu K, Tjhang JG, Jang I-C, Rajani S. Sweet Basil Has Distinct Synthases for Eugenol Biosynthesis in Glandular Trichomes and Roots with Different Regulatory Mechanisms. International Journal of Molecular Sciences. 2021; 22(2):681. https://doi.org/10.3390/ijms22020681
Chicago/Turabian StyleReddy, Vaishnavi Amarr, Chunhong Li, Kumar Nadimuthu, Jessica Gambino Tjhang, In-Cheol Jang, and Sarojam Rajani. 2021. "Sweet Basil Has Distinct Synthases for Eugenol Biosynthesis in Glandular Trichomes and Roots with Different Regulatory Mechanisms" International Journal of Molecular Sciences 22, no. 2: 681. https://doi.org/10.3390/ijms22020681