Modulation of Intestinal Phosphate Transport in Young Goats Fed a Low Phosphorus Diet

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Intake, Body Weight and Daily Body Weight Gain of Young Goats Fed a Low P Diet
2.2. Concentrations of Pi and Ca in Body Fluids of Young Goats Fed a Low P Diet
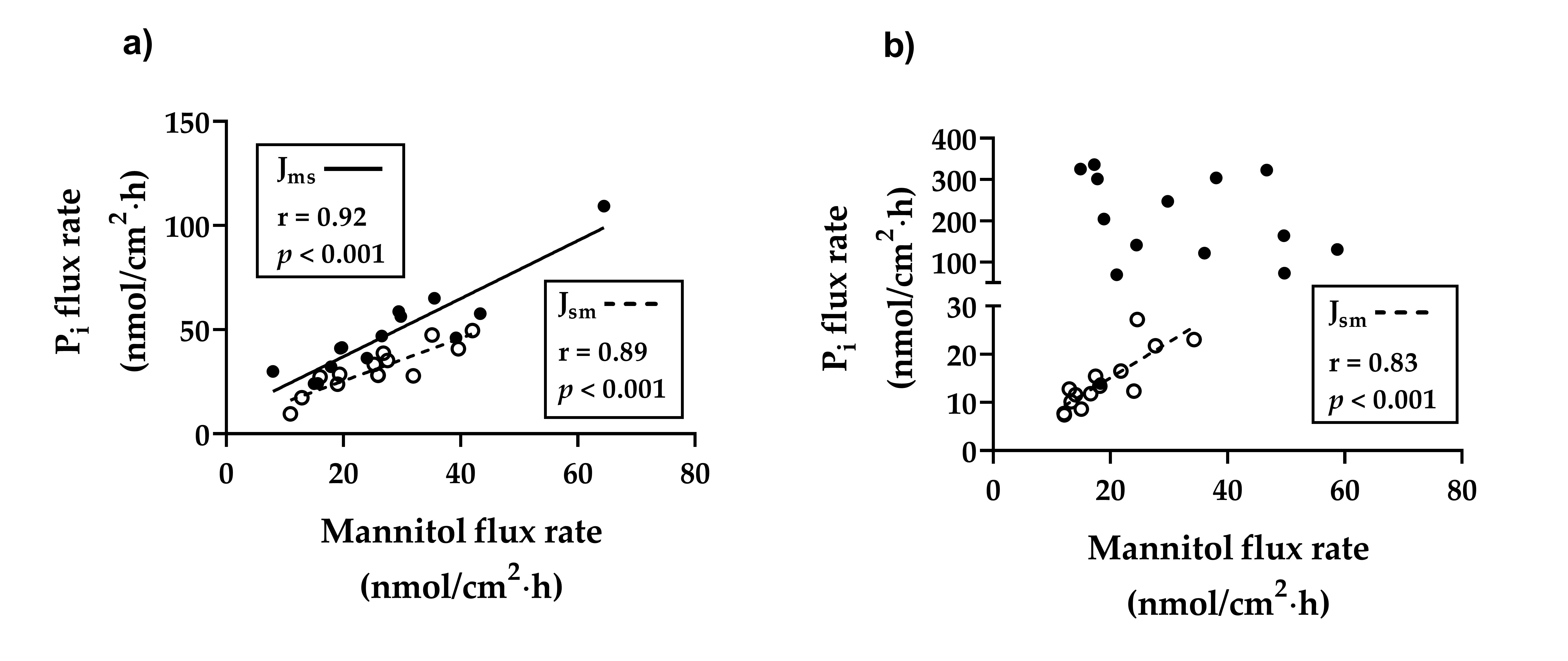
2.3. Flux Rates of Pi Across the Duodenal and Ileal Epithelia of Young Goats Fed a Low P Diet
2.4. Electrophysiological Parameters of Duodenal and Ileal Epithelia of Young Goats Fed a Low P Diet
2.5. Intestinal mRNA Expression of Na+/K+-ATPase, NaPiIIb, PiT1, PiT2, VDR, XPR1, Cadherin-17, Claudin-1, Claudin-2, Claudin-12, Claudin-15, Occludin and ZO-1 in Young Goats Fed a Low P Diet
2.6. Intestinal Protein Expressions of Na+/K+-ATPase, NaPiIIb and PiT1 in Ileal Epithelia of Young Goats Fed a Low P Diet
3. Discussion
4. Materials and Methods
4.1. Animals and Feeding Regimes
4.2. Diets
4.3. Intestinal Tissue and Body Fluid Sampling
4.4. Ussing Chamber Experiments
4.4.1. Flux Rate Studies
4.4.2. Electrophysiological Studies
4.5. Biochemical Determinations
4.6. RNA Isolation and Reverse Transcription
4.7. Intestinal mRNA Expression of Na+/K+-ATPase, NaPiIIb, PiT1, PiT2, VDR, XPR1, Cadherin-17, Claudin-1, Claudin-2, Claudin-12, Claudin-15, Occludin and ZO-1
4.8. Intestinal Protein Expression of Na+/K+-ATPase, NaPiIIb and PiT1
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1,25-(OH)2D3 | 1,25-dihydroxy-vitamin D3 |
| 25-(OH)D3 | 25-hydroxy-vitamin D3 |
| AJ | Adherens-junctions |
| BBM | Brush border membranes |
| Ca | Calcium |
| CP | Crude protein |
| DM | Dry matter |
| K+ | Potassium |
| ME | Metabolisable energy |
| N | Nitrogen |
| Na+ | Sodium |
| Na+/K+-ATPase | Sodium-potassium adenosine triphosphatase |
| NaPiIIb | Type IIb sodium-dependent phosphate transporter |
| P | Phosphorus |
| Pi | Inorganic phosphate |
| RT | Room temperatur |
| PiT1/2 | Type III sodium-dependent phosphate transporter 1/2 |
| TJ | Tight-junctions |
| VDR | Vitamin D receptor |
| XPR1 | xenotropic and polytropic retrovirus receptor 1 |
| ZO-1 | Zonula occludens protein-1 |
References
- Radanovic, T.; Wagner, C.A.; Murer, H.; Biber, J. Regulation of intestinal phosphate transport. I. Segmental expression and adaptation to low-P(i) diet of the type IIb Na(+)-P(i) cotransporter in mouse small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G496–G500. [Google Scholar]
- Giral, H.; Caldas, Y.; Sutherland, E.; Wilson, P.; Breusegem, S.; Barry, N.; Blaine, J.; Jiang, T.; Wang, X.X.; Levi, M. Regulation of rat intestinal Na-dependent phosphate transporters by dietary phosphate. Am. J. Physiol. Ren. Physiol. 2009, 297, F1466–F1475. [Google Scholar]
- Elfers, K.; Wilkens, M.R.; Breves, G.; Muscher-Banse, A.S. Modulation of intestinal calcium and phosphate transport in young goats fed a nitrogen- and/or calcium-reduced diet. Br. J. Nutr. 2015, 114, 1949–1964. [Google Scholar] [PubMed] [Green Version]
- Schröder, B.; Kappner, H.; Failing, K.; Pfeffer, E.; Breves, G. Mechanisms of intestinal phosphate transport in small ruminants. Br. J. Nutr. 1995, 74, 635–648. [Google Scholar] [PubMed] [Green Version]
- Virkki, L.V.; Biber, J.; Murer, H.; Forster, I.C. Phosphate transporters: A tale of two solute carrier families. Am. J. Physiol. Ren. Physiol. 2007, 293, F643–F654. [Google Scholar]
- Feild, J.A.; Zhang, L.; Brun, K.A.; Brooks, D.P.; Edwards, R.M. Cloning and functional characterization of a sodium-dependent phosphate transporter expressed in human lung and small intestine. Biochem. Biophys. Res. Commun. 1999, 258, 578–582. [Google Scholar] [PubMed]
- Forster, I.C.; Hernando, N.; Biber, J.; Murer, H. Phosphate transporters of the SLC20 and SLC34 families. Mol. Asp. Med. 2013, 34, 386–395. [Google Scholar]
- Shirazi-Beechey, S.P.; Beechey, R.B.; Penny, J.; Vayro, S.; Buchan, W.; Scott, D. Mechanisms of phosphate-transport in sheep intestine and parotid-gland—Response to variation in dietary phosphate supply. Exp. Physiol. 1991, 76, 231–241. [Google Scholar]
- Huber, K.; Walter, C.; Schröder, B.; Breves, G. Phosphate transport in the duodenum and jejunum of goats and its adaptation by dietary phosphate and calcium. Am. J. Physiol. Regul. I 2002, 283, R296–R302. [Google Scholar]
- Schröder, B.; Breves, G. Mechanisms of phosphate uptake into brush border membrane vesicles from goat jejunum. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 1996, 166, 230–240. [Google Scholar]
- Huber, K.; Walter, C.; Schröder, B.; Biber, J.; Murer, H.; Breves, G. Epithelial phosphate transporters in small ruminants. Ann. N. Y. Acad. Sci. 2000, 915, 95–97. [Google Scholar] [PubMed]
- Candeal, E.; Caldas, Y.A.; Guillen, N.; Levi, M.; Sorribas, V. Intestinal phosphate absorption is mediated by multiple transport systems in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G355–G366. [Google Scholar]
- Katai, K.; Miyamoto, K.; Kishida, S.; Segawa, H.; Nii, T.; Tanaka, H.; Tani, Y.; Arai, H.; Tatsumi, S.; Morita, K.; et al. Regulation of intestinal Na+-dependent phosphate co-transporters by a low-phosphate diet and 1,25-dihydroxyvitamin D3. Biochem. J. 1999, 343, 705–712. [Google Scholar] [PubMed]
- Hattenhauer, O.; Traebert, M.; Murer, H.; Biber, J. Regulation of small intestinal Na-P(i) type IIb cotransporter by dietary phosphate intake. Am. J. Physiol. 1999, 277, G756–G762. [Google Scholar] [PubMed]
- Capuano, P.; Radanovic, T.; Wagner, C.A.; Bacic, D.; Kato, S.; Uchiyama, Y.; St-Arnoud, R.; Murer, H.; Biber, J. Intestinal and renal adaptation to a low-Pi diet of type II NaPi cotransporters in vitamin D receptor- and 1alphaOHase-deficient mice. Am. J. Physiol. Cell Physiol. 2005, 288, C429–C434. [Google Scholar]
- Wild, R.; Gerasimaite, R.; Jung, J.Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M.; et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [Google Scholar] [PubMed] [Green Version]
- Giovannini, D.; Touhami, J.; Charnet, P.; Sitbon, M.; Battini, J.L. Inorganic phosphate export by the retrovirus receptor XPR1 in metazoans. Cell Rep. 2013, 3, 1866–1873. [Google Scholar]
- Van Itallie, C.M.; Anderson, J.M. The molecular physiology of tight junction pores. Physiology 2004, 19, 331–338. [Google Scholar]
- Markov, A.G.; Veshnyakova, A.; Fromm, M.; Amasheh, M.; Amasheh, S. Segmental expression of claudin proteins correlates with tight junction barrier properties in rat intestine. J. Comp. Physiol. B 2010, 180, 591–598. [Google Scholar]
- Rosenthal, R.; Gunzel, D.; Piontek, J.; Krug, S.M.; Ayala-Torres, C.; Hempel, C.; Theune, D.; Fromm, M. Claudin-15 forms a water channel through the tight junction with distinct function compared to claudin-2. Acta Physiol. 2020, 228, e13334. [Google Scholar]
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishii, N.; Oshima, T.; Li, M.; Eda, H.; Nakamura, K.; Tamura, A.; Ogawa, T.; Yamasaki, T.; Kondo, T.; Kono, T.; et al. Lubiprostone induces claudin-1 and protects intestinal barrier function. Pharmacology 2020, 105, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: A lesson from claudin-1-deficient mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, B.R.; Siliciano, J.D.; Mooseker, M.S.; Goodenough, D.A. Identification of ZO-1: A high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J. Cell Biol. 1986, 103, 755–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Itallie, C.M.; Fanning, A.S.; Bridges, A.; Anderson, J.M. ZO-1 stabilizes the tight junction solute barrier through coupling to the perijunctional cytoskeleton. Mol. Biol. Cell 2009, 20, 3930–3940. [Google Scholar] [CrossRef] [Green Version]
- Al-Sadi, R.; Khatib, K.; Guo, S.; Ye, D.; Youssef, M.; Ma, T. Occludin regulates macromolecule flux across the intestinal epithelial tight junction barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G1054–G1064. [Google Scholar] [CrossRef] [Green Version]
- Angst, B.; Marcozzi, C.; Magee, A. The cadherin superfamily: Diversity in form and function. J. Cell Sci. 2001, 114, 629–641. [Google Scholar]
- Kutuzova, G.D.; Deluca, H.F. Gene expression profiles in rat intestine identify pathways for 1,25-dihydroxyvitamin D(3) stimulated calcium absorption and clarify its immunomodulatory properties. Arch. Biochem. Biophys. 2004, 432, 152–166. [Google Scholar] [CrossRef]
- Drochner, W.; Ernährungsphysiologie, G.F.; Bedarfsnormen, A.F. Recommendations for the Supply of Energy and Nutrients to Goats = Empfehlungen zur Energie- und Nährstoffversorgung der Ziegen; DLG-Verlags-GmbH: Frankfurt am Main, Germany, 2003. [Google Scholar]
- Field, A.C.; Suttle, N.F.; Nisbet, D.I. Effect of diets low in calcium and phosphorus on development of growing lambs. J. Agric. Sci. 1975, 85, 435–442. [Google Scholar] [CrossRef]
- Lee, D.B.; Brautbar, N.; Walling, M.W.; Silis, V.; Coburn, J.W.; Kleeman, C.R. Effect of phosphorus depletion on intestinal calcium and phosphorus absorption. Am. J. Physiol. 1979, 236, E451–E457. [Google Scholar] [CrossRef]
- Schröder, B.; Pfeffer, E.; Failing, K.; Breves, G. Binding properties of goat intestinal vitamin D receptors as affected by dietary calcium and/or phosphorus depletion. Zentralbl. Veterinarmed. A 1995, 42, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Pickard, D.W.; Care, A.D.; Murray, T.M. Effect of low phosphorus diets on intestinal calcium absorption and the concentration of calcium-binding protein in intact and parathyroidectomized pigs. J. Endocrinol. 1978, 78, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Baylink, D.; Wergedal, J.; Stauffer, M. Formation, mineralization, and resorption of bone in hypophosphatemic rats. J. Clin. Investig. 1971, 50, 2519–2530. [Google Scholar] [CrossRef] [PubMed]
- Grunberg, W.; Witte, S.; Cohrs, I.; Golbeck, L.; Brouwers, J.F.; Muller, A.E.; Schmicke, M. Liver phosphorus content and liver function in states of phosphorus deficiency in transition dairy cows. PLoS ONE 2019, 14, e0219546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, T.J.; Nichols, W.W.; Brenner, B.J.; Peterson, J.C. Reversible depression in myocardial performance in dogs with experimental phosphorus deficiency. J. Clin. Investig. 1978, 62, 1194–1200. [Google Scholar] [CrossRef]
- Harper, A.F.; Kornegay, E.T.; Schell, T.C. Phytase supplementation of low-phosphorus growing-finishing pig diets improves performance, phosphorus digestibility, and bone mineralization and reduces phosphorus excretion. J. Anim. Sci. 1997, 75, 3174–3186. [Google Scholar] [CrossRef] [PubMed]
- Milton, J.T.B.; Ternouth, J.H. Phosphorus-metabolism in ruminants .2. Effects of inorganic phosphorus concentration upon food-intake and digestibility. Aust. J. Agric. Res. 1985, 36, 647–654. [Google Scholar] [CrossRef]
- Kamphues, J.; Muller, E.; Drochner, W. Clinical effects of an extreme Ca overdose in feed for growing piglets. Tierarztl. Prax. 1990, 18, 251–254. [Google Scholar]
- Wimsatt, J.; Marks, S.L.; Campbell, T.W.; Johnson, J.D.; Nachreiner, R.F. Dietary vitamin D toxicity in a household of pot-bellied pigs (Sus scrofa). J. Vet. Intern. Med. 1998, 12, 42–44. [Google Scholar] [CrossRef]
- Midkiff, A.M.; Chew, D.J.; Randolph, J.F.; Center, S.A.; DiBartola, S.P. Idiopathic hypercalcemia in cats. J. Vet. Intern. Med. 2000, 14, 619–626. [Google Scholar] [CrossRef]
- Lichtman, M.A.; Miller, D.R.; Freeman, R.B. Erythrocyte adenosine triphosphate depletion during hypophosphatemia in a uremic subject. N. Engl. J. Med. 1969, 280, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Finch, N.C. Hypercalcaemia in cats: The complexities of calcium regulation and associated clinical challenges. J. Feline Med. Surg. 2016, 18, 387–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragno, A.; Pepe, J.; Badiali, D.; Minisola, S.; Romagnoli, E.; Severi, C.; D’erasmo, E. Chronic constipation in hypercalcemic patients with primary hyperparathyroidism. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 884–889. [Google Scholar] [PubMed]
- Jungbluth, H.; Binswanger, U. Unidirectional duodenal and jejunal calcium and phosphorus transport in the rat: Effects of dietary phosphorus depletion, ethane-1-hydroxy-1,1-diphosphonate and 1,25 dihydroxycholecalciferol. Res. Exp. Med. 1989, 189, 439–449. [Google Scholar] [CrossRef]
- Breves, G.; Schröder, B. Comparative aspects of gastrointestinal phosphorus metabolism. Nutr. Res. Rev. 1991, 4, 125–140. [Google Scholar] [CrossRef]
- Muschen, H.; Petri, A.; Breves, G.; Pfeffer, E. Response of lactating goats to low phosphorus intake .1. Milk-yield and fecal excretion of P and Ca. J. Agric. Sci. 1988, 111, 255–263. [Google Scholar] [CrossRef]
- Kincaid, R.; Rodehutscord, M. Phosphorus metabolism in the rumen reducing the environmental impact of cattle operations. In Nitrogen and Phosphorus Nutrition of Cattle: Reducing the Environmental Impact of Cattle Operations; CABI Publishing: Wallington, UK, 2005; pp. 187–194. [Google Scholar]
- Hughes, M.R.; Brumbaugh, P.F.; Hussler, M.R.; Wergedal, J.E.; Baylink, D.J. Regulation of serum 1alpha,25-dihydroxyvitamin D3 by calcium and phosphate in the rat. Science 1975, 190, 578–580. [Google Scholar] [CrossRef]
- Sommerville, B.A.; Maunder, E.; Ross, R.; Care, A.D.; Brown, R.C. Effect of dietary calcium and phosphorus depletion on vitamin D metabolism and calcium binding protein in the growing pig. Horm. Metab. Res. 1985, 17, 78–81. [Google Scholar] [CrossRef]
- Baxter, L.A.; DeLuca, H.F. Stimulation of 25-hydroxyvitamin D3-1alpha-hydroxylase by phosphate depletion. J. Biol. Chem. 1976, 251, 3158–3161. [Google Scholar] [CrossRef]
- Young, V.R.; Richards, W.P.; Lofgreen, G.P.; Luick, J.R. Phosphorus depletion in sheep and ratio of calcium to phosphorus in diet with reference to calcium and phosphorus absorption. Br. J. Nutr. 1966, 20, 783–794. [Google Scholar] [CrossRef]
- Abdel-Hafeez, H.M.; Manas-Almendros, M.; Ross, R.; Care, A.D.; Marshall, D.H. Effects of dietary phosphorus and calcium on the intestinal absorption of Ca in sheep. Br. J. Nutr. 1982, 47, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Bushinsky, D.A.; Riera, G.S.; Favus, M.J.; Coe, F.L. Evidence that blood ionized calcium can regulate serum 1,25(OH)2D3 independently of parathyroid hormone and phosphorus in the rat. J. Clin. Investig. 1985, 76, 1599–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, T.; Ikeda, K.; Morita, K.; Fukumoto, S.; Takahashi, H.; Ogata, E. Blood Ca2+ modulates responsiveness of renal 25(OH)D3-1 alpha-hydroxylase to PTH in rats. Am. J. Physiol. 1987, 253, E503–E507. [Google Scholar] [CrossRef] [PubMed]
- Kowarski, S.; Schachter, D. Effects of vitamin D on phosphate transport and incorporation into mucosal constituents of rat intestinal mucosa. J. Biol. Chem. 1969, 244, 211–217. [Google Scholar] [CrossRef]
- Hernando, N.; Pastor-Arroyo, E.M.; Marks, J.; Schnitzbauer, U.; Knopfel, T.; Burki, M.; Bettoni, C.; Wagner, C.A. 1,25(OH)2 vitamin D3 stimulates active phosphate transport but not paracellular phosphate absorption in mouse intestine. J. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Danisi, G.; Bonjour, J.P.; Straub, R.W. Regulation of Na-dependent phosphate influx across the mucosal border of duodenum by 1,25-dihydroxycholecalciferol. Pflug. Arch. 1980, 388, 227–232. [Google Scholar] [CrossRef]
- Cao, M.; Fang, R.; Chen, J.; He, J. Vitamin D3 increased intestinal Na/Pi-IIb and CYP27B1 mRNA level in rats fed low-phosphorus diets. Anim. Nutr. 2016, 2, 211–217. [Google Scholar] [CrossRef]
- Kido, S.; Miyamoto, K.; Mizobuchi, H.; Taketani, Y.; Ohkido, I.; Ogawa, N.; Kaneko, Y.; Harashima, S.; Takeda, E. Identification of regulatory sequences and binding proteins in the type II sodium/phosphate cotransporter NPT2 gene responsive to dietary phosphate. J. Biol. Chem. 1999, 274, 28256–28263. [Google Scholar] [CrossRef] [Green Version]
- Segawa, H.; Kaneko, I.; Yamanaka, S.; Ito, M.; Kuwahata, M.; Inoue, Y.; Kato, S.; Miyamoto, K. Intestinal Na-P(i) cotransporter adaptation to dietary P(i) content in vitamin D receptor null mice. Am. J. Physiol. Ren. Physiol. 2004, 287, F39–F47. [Google Scholar] [CrossRef] [Green Version]
- Starke, S.; Reimers, J.; Muscher-Banse, A.S.; Schröder, B.; Breves, G.; Wilkens, M.R. Gastrointestinal transport of calcium and phosphate in lactating goats. Livest. Sci. 2016, 189, 23–31. [Google Scholar] [CrossRef]
- Foote, A.P.; Lambert, B.D.; Brady, J.A.; Muir, J.P. Phosphate transporter expression in Holstein cows. J. Dairy Sci. 2011, 94, 1913–1916. [Google Scholar] [CrossRef] [PubMed]
- Geall, M.G.; Summerskill, W.H. Electric-potential difference—A neglected parameter of gut integrity and function? Gut 1969, 10, 418–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knöpfel, T.; Himmerkus, N.; Guenzel, D.; Bleich, M.; Hernando, N.; Wagner, C.A. Paracellular transport of phosphate along the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G233–G241. [Google Scholar] [CrossRef] [PubMed]
- Ikuta, K.; Segawa, H.; Sasaki, S.; Hanazaki, A.; Fujii, T.; Kushi, A.; Kawabata, Y.; Kirino, R.; Sasaki, S.; Noguchi, M.; et al. Effect of Npt2b deletion on intestinal and renal inorganic phosphate (Pi) handling. Clin. Exp. Nephrol. 2018, 22, 517–528. [Google Scholar] [CrossRef]
- Salaun, C.; Gyan, E.; Rodrigues, P.; Heard, J.M. Pit2 assemblies at the cell surface are modulated by extracellular inorganic phosphate concentration. J. Virol. 2002, 76, 4304–4311. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, P.; Heard, J.M. Modulation of phosphate uptake and amphotropic murine leukemia virus entry by posttranslational modifications of PIT-2. J. Virol. 1999, 73, 3789–3799. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.H. Blockage of gallbladder tight junction cation-selective channels by 2,4,6-triaminopyrimidinium (TAP). J. Gen. Physiol. 1975, 66, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Clarke, L.L. A guide to Ussing chamber studies of mouse intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1151–G1166. [Google Scholar] [CrossRef] [Green Version]
- Marks, J.; Debnam, E.S.; Unwin, R.J. Phosphate homeostasis and the renal-gastrointestinal axis. Am. J. Physiol. Ren. Physiol. 2010, 299, F285–F296. [Google Scholar] [CrossRef] [Green Version]
- Elfers, K.; Marr, I.; Wilkens, M.R.; Breves, G.; Langeheine, M.; Brehm, R.; Muscher-Banse, A.S. Expression of tight junction proteins and cadherin 17 in the small intestine of young goats offered a reduced N and/or Ca diet. PLoS ONE 2016, 11, e0154311. [Google Scholar] [CrossRef] [Green Version]
- Ravera, S.; Virkki, L.V.; Murer, H.; Forster, I.C. Deciphering PiT transport kinetics and substrate specificity using electrophysiology and flux measurements. Am. J. Physiol. Cell Physiol. 2007, 293, C606–C620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haussler, M.R.; Whitfield, G.K.; Haussler, C.A.; Hsieh, J.C.; Thompson, P.D.; Selznick, S.H.; Dominguez, C.E.; Jurutka, P.W. The nuclear vitamin D receptor: Biological and molecular regulatory properties revealed. J. Bone Miner. Res. 1998, 13, 325–349. [Google Scholar] [CrossRef] [PubMed]
- Demay, M.B. Mechanism of vitamin D receptor action. Ann. N. Y. Acad. Sci. 2006, 1068, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Sriussadaporn, S.; Wong, M.S.; Pike, J.W.; Favus, M.J. Tissue specificity and mechanism of vitamin D receptor up-regulation during dietary phosphorus restriction in the rat. J. Bone Miner. Res. 1995, 10, 271–280. [Google Scholar] [CrossRef]
- Schröder, B.; Breves, G.; Pfeffer, E. Binding properties of duodenal 1,25-dihydroxyvitamin D3 receptors as affected by phosphorus depletion in lactating goats. Comp. Biochem. Physiol. Comp. Physiol. 1990, 96, 495–498. [Google Scholar] [CrossRef]
- Frizzell, R.A.; Schultz, S.G. Ionic conductances of extracellular shunt pathway in rabbit ileum. Influence of shunt on transmural sodium transport and electrical potential differences. J. Gen. Physiol. 1972, 59, 318–346. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ghedalia, D.; Tagari, H.; Zamwel, S.; Bondi, A. Solubility and net exchange of calcium, magnesium and phosphorus in digesta flowing along the gut of the sheep. Br. J. Nutr. 1975, 33, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Naumann, C.; Bassler, R. Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten. Methodenbuch Band III. Die chemische Untersuchung von Futtermitteln. Mit Ergänzungslieferungen 1983, 1988, 1993, 1997 (Association of German Agriculture Testing and Research Institutes. Methods Book Volume III. The Chemical Analysis of Feed. With Additional Deliveries in 1983, 1988, 1993, 1997); VDLUFA-Verlag: Darmstadt, Germany, 1976. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Busch, A.; Wagner, C.; Schuster, A.; Waldegger, S.; Biber, J.; Murer, H.; Lang, F. Properties of electrogenic Pi transport by a human renal brush border Na+/Pi transporter. J. Am. Soc. Nephrol. JASN 1996, 6, 1547–1551. [Google Scholar]
- Simmons, N.L.; Naftalin, R.J. Bidirectional sodium ion movements via the paracellular and transcellular routes across short-circuited rabbit ileum. Biochim. Biophys. Acta 1976, 448, 426–450. [Google Scholar] [CrossRef]
- Auchere, D.; Tardivel, S.; Gounelle, J.C.; Drueke, T.; Lacour, B. Role of transcellular pathway in ileal Ca2+ absorption: Stimulation by low-Ca2+ diet. Am. J. Physiol. 1998, 275, G951–G956. [Google Scholar] [CrossRef] [PubMed]
- Schultz, S.G.; Zalusky, R. Ion transport in isolated rabbit ileum. I. Short-circuit current and Na fluxes. J. Gen. Physiol. 1964, 47, 567–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, B.C.; Chauhan, U.P. A new method for determining micro quantities of calcium in biological materials. Anal. Biochem. 1967, 20, 155–166. [Google Scholar] [CrossRef]
- Kruse-Jarres, J. Klinische Chemie, Spezielle Klinische Analytik (Clinical Chemistry, Special Clinical Analysis); Urban & Fischer Mchn: Stuttgart, Germany, 1979; p. 288. [Google Scholar]
- Wilkens, M.R.; Kunert-Keil, C.; Brinkmeier, H.; Schröder, B. Expression of calcium channel TRPV6 in ovine epithelial tissue. Vet. J. 2009, 182, 294–300. [Google Scholar] [CrossRef] [PubMed]
- BLAST NCBI (National Center for Biotechnology Information). Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 15 January 2021).
- Wilkens, M.R.; Elfers, K.; Schmicke, M.; Breves, G.; Muscher-Banse, A.S. Dietary nitrogen and calcium modulate CYP27B1 expression in young goats. Domest. Anim. Endocrinol. 2018, 64, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Herm, G.; Muscher-Banse, A.S.; Breves, G.; Schröder, B.; Wilkens, M.R. Renal mechanisms of calcium homeostasis in sheep and goats. J. Anim. Sci. 2015, 93, 1608–1621. [Google Scholar] [CrossRef]
- Wilkens, M.R.; Mrochen, N.; Breves, G.; Schröder, B. Gastrointestinal calcium absorption in sheep is mostly insensitive to an alimentary induced challenge of calcium homeostasis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Muscher, A.; Hattendorf, J.; Pfeffer, E.; Breves, G.; Huber, K. Hormonal regulation of phosphate homeostasis in goats during transition to rumination. J. Comp. Physiol. B 2008, 178, 585–596. [Google Scholar] [CrossRef]
- GraphPad. Available online: https://www.graphpad.com/ (accessed on 15 January 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| DM intake (g/day) | 614 ± 10 | 528 ± 25 | 0.008 |
| Concentrate intake (g/day) | 555 ± 12 | 469 ± 29 | 0.017 |
| P intake (g/day) | 2.40 ± 0.05 | 0.53 ± 0.03 | <0.001 |
| Ca intake (g/day) | 6.50 ± 0.13 | 5.42 ± 0.31 | 0.008 |
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| Initial body weight (kg) * | 19.21 ± 0.75 | 18.93 ± 1.09 | 0.83 |
| Final body weight (kg) † | 25.29 ± 0.74 | 20.21 ± 0.93 | 0.001 |
| Body weight gain (kg/d) | 0.14 ± 0.01 | 0.03 ± 0.01 | <0.001 |
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| Plasma Pi (mmol/L) | 2.09 ± 0.12 | 0.70 ± 0.04 | <0.001 |
| Plasma Ca (mmol/L) | 3.16 ± 0.09 | 4.34 ± 0.17 | <0.001 |
| Saliva Pi (mmol/L) | 31.77 ± 3.96 | 10.41 ± 0.83 | <0.001 |
| Saliva Ca (mmol/L) | 0.42 ± 0.03 | 0.53 ± 0.02 | 0.01 |
| Ruminal fluid Pi (mmol/L) | 46.69 ± 6.11 | 6.07 ± 0.76 | <0.001 |
| Ruminal fluid Ca (mmol/L) | 1.68 ± 0.11 | 5.75 ± 1.39 | 0.01 |
| Abomasal fluid Pi (mmol/L) | 39.02 ± 3.91 | 7.77 ± 0.78 | <0.001 |
| Abomasal fluid Ca (mmol/L) | 17.83 ± 1.79 | 23.80 ± 3.48 | 0.15 |
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| Duodenum | |||
| Pi Jms | 46.72 ± 5.69 | 48.85 ± 10.80 | 0.87 |
| Pi Jsm | 32.92 ± 5.11 | 29.30 ± 3.09 | 0.56 |
| Pi Jnet | 13.80 ± 4.91 | 19.55 ± 10.59 | 0.63 |
| Man Jms | 26.31 ± 4.16 | 29.14 ± 6.87 | 0.73 |
| Man Jsm | 26.13 ± 3.99 | 25.08 ± 3.38 | 0.84 |
| Man Jnet | 0.18 ± 3.52 | 4.06 ± 4.90 | 0.53 |
| Ileum | |||
| Pi Jms | 215.49 ± 39.03 | 177.91 ± 45.56 | 0.54 |
| Pi Jsm | 13.12 ± 2.49 | 15.44 ± 2.14 | 0.49 |
| Pi Jnet | 202.37 ± 40.09 | 162.47 ± 46.85 | 0.53 |
| Man Jms | 29.30 ± 5.51 | 33.77 ± 5.99 | 0.59 |
| Man Jsm | 17.08 ± 1.95 | 20.70 ± 3. 01 | 0.33 |
| Man Jnet | 12.22 ± 4.48 | 13.07 ± 3.41 | 0.88 |
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| Duodenum | |||
| Isc | 0.61 ± 0.06 | 0.71 ± 0.04 | 0.20 |
| Gt | 18.92 ± 1.83 | 18.05 ± 0.97 | 0.68 |
| Ileum | |||
| Isc | 1.73 ± 0.29 | 1.12 ± 0.24 | 0.13 |
| Gt | 17.18 ± 1.70 | 16.78 ± 1.25 | 0.85 |
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| Duodenum | |||
| ∆Isc Na+-arsenate | 0.31 ± 0.06 | 0.39 ± 0.03 | 0.25 |
| ∆Isc TAP | −0.15 ± 0.03 | −0.13 ± 0.03 | 0.59 |
| ∆Gt Na+-arsenate | 4.91 ± 0.87 | 8.65 ± 2.13 | 0.13 |
| ∆Gt TAP | −3.96 ± 0.98 | −3.03 ± 1.19 | 0.56 |
| Ileum | |||
| ∆Isc Na+-arsenate | −0.79 ± 0.21 | −0.44 ± 0.18 | 0.23 |
| ∆Isc TAP | −0.41 ± 0.17 | −0.33 ± 0.10 | 0.69 |
| ∆Gt Na+-arsenate | 8.42 ± 1.61 | 6.10 ± 1.93 | 0.37 |
| ∆Gt TAP | −1.69 ± 0.44 | −2.17 ± 0.50 | 0.49 |
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| Duodenum | |||
| Na+/K+-ATPase | 1.657 ± 0.318 | 1.615 ± 0.252 | 0.921 |
| NaPiIIb | 1.361 × 10−6 ± 5.762 × 10−7 | 6.360 × 10−6 ± 2.581 × 10−6 | 0.083 |
| PiT1 | 0.010 ± 0.001 | 0.012 ± 0.003 | 0.606 |
| PiT2 | 0.008 ± 0.004 | 0.010 ± 0.002 | 0.621 |
| VDR | 0.019 ± 0.002 | 0.018 ± 0.003 | 0.966 |
| XPR1 | 0.089 ± 0.016 | 0.088 ± 0.012 | 0.959 |
| Cadherin-17 | 1.494 ± 0.384 | 1.508 ± 0.162 | 0.974 |
| Claudin-1 | 0.017 ± 0.003 | 0.015 ± 0.003 | 0.657 |
| Claudin-2 | 0.228 ± 0.028 | 0.215 ± 0.048 | 0.824 |
| Claudin-12 | 0.004 ± 0.001 | 0.005 ± 0.001 | 0.603 |
| Claudin-15 | 0.104 ± 0.011 | 0.112 ± 0.019 | 0.712 |
| Occludin | 0.041 ± 0.006 | 0.042 ± 0.005 | 0.948 |
| ZO-1 | 0.011 ± 0.002 | 0.010 ± 0.001 | 0.660 |
| Mid jejunum | |||
| Na+/K+-ATPase | 2.925 ± 0.399 | 2.589 ± 0.208 | 0.468 |
| NaPiIIb | 0.158 ± 0.050 | 0.0831 ± 0.018 | 0.189 |
| PiT1 | 0.011 ± 0.002 | 0.013 ± 0.002 | 0.537 |
| PiT2 | 0.009 ± 0.001 | 0.008 ± 0.001 | 0.618 |
| VDR | 0.060 ± 0.009 | 0.050 ± 0.003 | 0.317 |
| XPR1 | 0.020 ± 0.002 | 0.020 ± 0.002 | 0.948 |
| Cadherin-17 | 2.197 ± 0.369 | 1.718 ± 0.119 | 0.240 |
| Claudin-1 | 0.071 ± 0.008 | 0.073 ± 0.012 | 0.889 |
| Claudin-2 | 0.759 ± 0.117 | 0.391 ± 0.088 | 0.027 |
| Claudin-12 | 0.010 ± 0.002 | 0.009 ± 0.002 | 0.743 |
| Claudin-15 | 0.788 ± 0.134 | 0.617 ± 0.076 | 0.287 |
| Occludin | 0.146 ± 0.030 | 0.111 ± 0.010 | 0.289 |
| ZO-1 | 0.014 ± 0.002 | 0.011 ± 0.0005 | 0.215 |
| Ileum | |||
| Na+/K+-ATPase | 0.911 ± 0.195 | 1.175 ± 0.123 | 0.274 |
| NaPiIIb | 0.006 ± 0.001 | 0.005 ± 0.002 | 0.858 |
| PiT1 | 0.017 ± 0.003 | 0.025 ± 0.002 | 0.038 |
| PiT2 | 0.014 ± 0.002 | 0.020 ± 0.002 | 0.046 |
| VDR | 0.007 ± 0.001 | 0.007 ± 0.001 | 0.890 |
| XPR1 | 0.014 ± 0.002 | 0.017 ± 0.002 | 0.357 |
| Cadherin-17 | 2.582 ± 0.425 | 3.478 ± 0.450 | 0.174 |
| Claudin-1 | 0.013 ± 0.002 | 0.017 ± 0.003 | 0.277 |
| Claudin-2 | 0.461 ± 0.088 | 0.510 ± 0.085 | 0.699 |
| Claudin-12 | 0.009 ± 0.002 | 0.009 ± 0.001 | 0.990 |
| Claudin-15 | 0.045 ± 0.012 | 0.040 ± 0.005 | 0.718 |
| Occludin | 0.136 ± 0.029 | 0.187 ± 0.032 | 0.254 |
| ZO-1 | 0.020 ± 0.004 | 0.027 ± 0.003 | 0.164 |
| Items | Control | P Reduction | p-Value |
|---|---|---|---|
| Na+/K+-ATPase | 0.119 ± 0.004 | 0.115 ± 0.005 | 0.61 |
| NaPiIIb | 0.040 ± 0.006 | 0.027 ± 0.005 | 0.11 |
| PiT1 | 0.012 ± 0.002 | 0.007 ± 0.001 | 0.03 |
| Items | Wheat Straw | Control | P Reduction |
|---|---|---|---|
| Components (as fed) (g/kg) | |||
| Soybean meal | 68.0 | 68.0 | |
| Urea | 24.5 | 24.5 | |
| Wheat starch | 378 | 379 | |
| Beet pulp | 399 | 399 | |
| Mineral-vitamin premix † | 10.0 | 10.0 | |
| MgHPO₄ · 3 H₂O | 9.3 | - | |
| MgO | - | 2.2 | |
| NaH2PO4 · 2 H2O | 9.7 | 0.4 | |
| NaCl | 1.4 | 1.4 | |
| NaHCO3 | - | 5.0 | |
| CaCO3 | 14.3 | 14.3 | |
| Sipernat 22S ‡ | 41.8 | 52.3 | |
| Molasses | 10.0 | 10.0 | |
| Soybean oil | 34.0 | 34.0 | |
| Composition | |||
| DM (g/kg) | 904 | 880 | 884 |
| Nutrients (g/kg DM) | |||
| Crude ash | 39.8 | 100 | 101 |
| CP | 25.4 | 165 | 169 |
| ADFom | 579 | 87.5 | 87.1 |
| aNDFom | 855 | 157 | 179 |
| Crude fat | 24.3 | 43.2 | 47.5 |
| Urea | BDL | 27.8 | 31.1 |
| Ca | 2.8 | 12.6 | 12.3 |
| P | 0.6 | 4.8 | 1.1 |
| Vitamin D3 (IU/kg DM) | BDL | 1000 | 1075 |
| ME (MJ/kg DM) | 8.3 | 12.8 | 12.9 |
| Genes | Primers and Probes (5′ → 3′) | Accession Number | References |
|---|---|---|---|
| 18S | Forward: AAAAATAACAATACAGGACTCTTTCG Reverse: GCTATTGGAGCTGGAATTACCG 6FAM-TggAATgAgTCCACTTTAAATCCTTCCgC--BBQ | KY129860 | [90] |
| Claudin-2 | Forward: CCAAAGACAGAGTGGCGGT Antisense: TCAAATTTCATGCTGTCAGGCAC FAM-TCCTGGGCTTCATCCCYGTTGC-BBQ | XM_005700206.3 | [72] |
| Claudin-12 | Sense: GCTGCTCTGCCTCATCGG Antisense: GCAGCCYGCACTATTGACCA FAM-TGTGTAACACGGCCTTCAGGTCCTC-BBQ | XM_005678898.3 | [72] |
| Genes | Primers (5′ → 3′) | Accession Number | References |
|---|---|---|---|
| Na+/K+-ATPase | Sense: TGGAACTCGGAGAAGAAGGA Antisense: GCCACTCGGTCCTGATATGT | XM_005690616.3 | [3] |
| NaPiIIb | Sense: CGGTCCAAAACAAAAGTATGATCAAG Reverse: AGCCAAAGGGGTAAGGGAA | XM_005681484.3 to XM_013964569.2 | [3] |
| PiT1 | Sense: ATTCATCCTCCGTAAGGCAGATC Antisense: CAGCAATGGTGCTCCAGTATACA | XM_018055327.1 | [3] |
| PiT2 | Forward: CCAATCTCGGGGACTCACTG Reverse: GGAACGGGGTCCTCCTTTTT | XM_018041866.1 to XM_018041872.1 | This study |
| VDR | Forward: GCACTTCCTTACCTGACCCC Reverse: CCGCTTGAGGATCATCTCCC | XM_004007435.1 | [91] |
| XPR1 | Forward: AATGCCGATGATCAGACGCT Reverse: AGCCTTGGATTGAGAAGCGA | XM_018060672.1 | This study |
| Cadherin-17 | Sense: CACCCTTTTGGTCATCGGTAT Antisense: CATCAGTTTCTCAGAGGCTTGACT | XM_018058228.1 | [72] |
| Claudin-1 | Forward: TTCATCCTGGCGTTTCTGGG Reverse: GTTGCTTGCAGAGTGCTGTT | XM_005675123.3 | This study |
| Claudin-15 | Forward: AGGGACTTCTTCGACCCCTT Reverse: CGTTATCACGGGGGCTTTGT | XM_013974836.2 to XM_018040880.1 | This study |
| Occludin | Sense: CTCGTCTGGATAAAGAACTGGATGA Antisense: CTCGTCTGGATAAAGAACTGGATGA | XM_018065677.1 to XM_018065681.1 | [72] |
| ZO-1 | Sense: CTCAGTACAGCCAGGGTGCT Reverse: TCCGGTTTGGACACTAATGAGTT | XM_018066114.1 to XM_018066118.1 | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behrens, J.L.; Schnepel, N.; Hansen, K.; Hustedt, K.; Burmester, M.; Klinger, S.; Breves, G.; Muscher-Banse, A.S. Modulation of Intestinal Phosphate Transport in Young Goats Fed a Low Phosphorus Diet. Int. J. Mol. Sci. 2021, 22, 866. https://doi.org/10.3390/ijms22020866
Behrens JL, Schnepel N, Hansen K, Hustedt K, Burmester M, Klinger S, Breves G, Muscher-Banse AS. Modulation of Intestinal Phosphate Transport in Young Goats Fed a Low Phosphorus Diet. International Journal of Molecular Sciences. 2021; 22(2):866. https://doi.org/10.3390/ijms22020866
Chicago/Turabian StyleBehrens, Joie L., Nadine Schnepel, Kathrin Hansen, Karin Hustedt, Marion Burmester, Stefanie Klinger, Gerhard Breves, and Alexandra S. Muscher-Banse. 2021. "Modulation of Intestinal Phosphate Transport in Young Goats Fed a Low Phosphorus Diet" International Journal of Molecular Sciences 22, no. 2: 866. https://doi.org/10.3390/ijms22020866
APA StyleBehrens, J. L., Schnepel, N., Hansen, K., Hustedt, K., Burmester, M., Klinger, S., Breves, G., & Muscher-Banse, A. S. (2021). Modulation of Intestinal Phosphate Transport in Young Goats Fed a Low Phosphorus Diet. International Journal of Molecular Sciences, 22(2), 866. https://doi.org/10.3390/ijms22020866