
The Role of Oxidative Stress in Sarcoidosis



Abstract
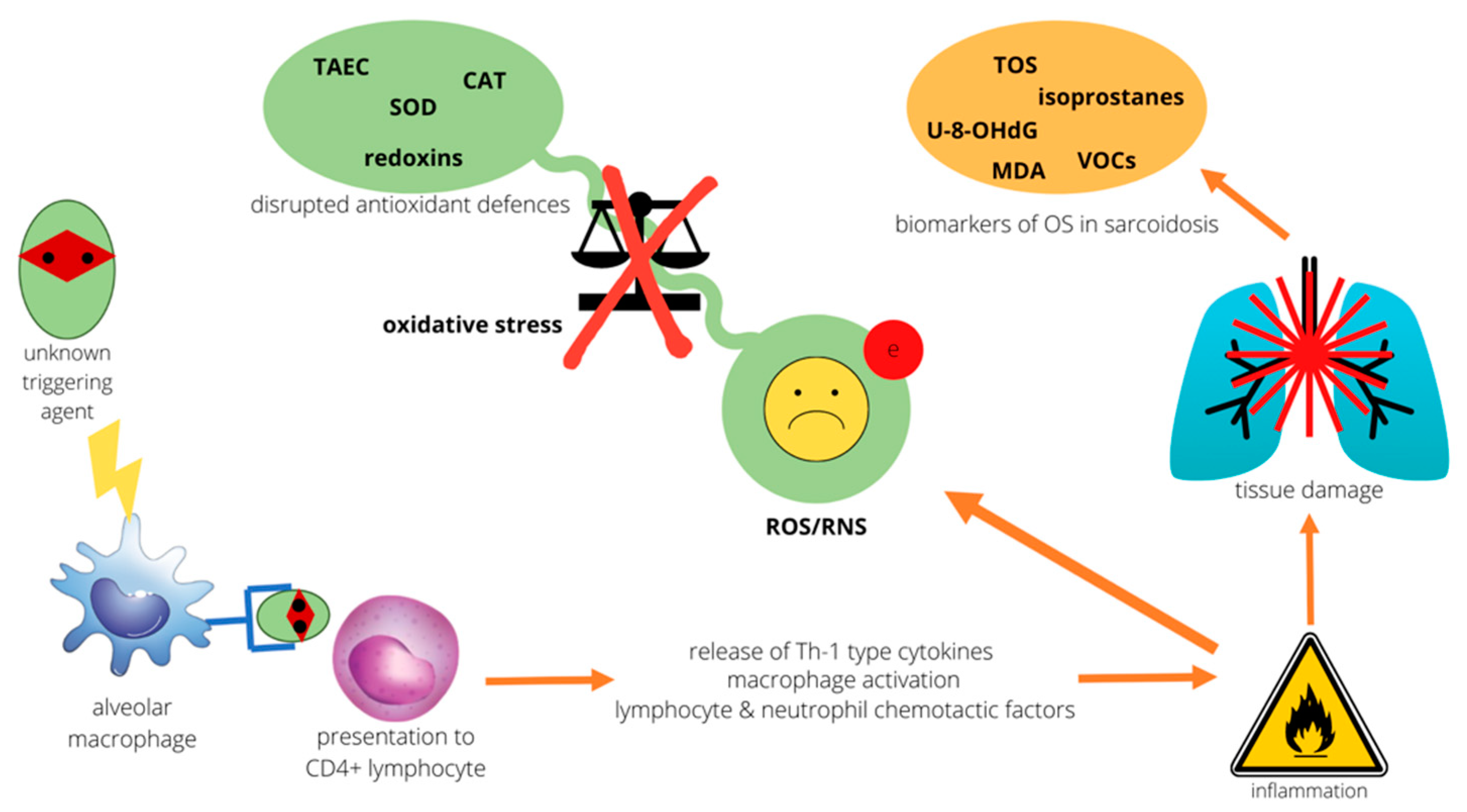
1. Introduction
2. Oxidative Stress and the Lung
3. Evidence of Redox Imbalance in Sarcoidosis
3.1. Biomarkers of Lipid Peroxidation: MDA, Isoprostanes, and VOCs
3.2. Oxidatively Damaged DNA
3.3. Disrupted Mitochondrial Homeostasis

{kind=link}
| Biomarker | Matrice | Advantages | Disadvantages | Reference |
|---|---|---|---|---|
| total oxidative status | blood | can be determined with various methods easy to reproduce correlates with disease activity | high oxidative status in blood may not be illustrative for that of matrices that are relevant to disease | [32,33,34] |
| MDA | blood | easy to determine correlates with disease activity | sampling can affect results | [38] |
| 8-isoprostane | blood BALF EBC | can be determined in various matrices correlates with disease activity, advanced stages, and pulmonary function tests | no correlation with BALF lymphocyte count in one study | [51,52,53] |
| exhaled ethane | EBC | determined with noninvasive method correlates with disease activity | single study on sarcoidosis | [58] |
| U-8-OHdG | urine | correlates with disease activity | single study on sarcoidosis | [60] |
4. Diminished Antioxidant Defenses
4.1. Redoxins
4.2. CAT and SOD
4.3. GSH and PON
| Antioxidant | Matrice | Characteristics | Comments | Reference |
|---|---|---|---|---|
| TAC | blood | easy to reproduce correlates with disease activity | low antioxidant status in blood may not reflect status of matrices that are relevant to disease | [68] |
| redoxins | lung biopsy specimens | Trx-rich antioxidant profile in sarcoidosis Grx-poor antioxidant profile in sarcoidosis in single study, as observed in other OS-related diseases | Trx may serve as marker of cell regeneration/inflammation | [72,74,75] |
| CAT | lung biopsy specimens | abundant in normal lung, no altered expression seen in sarcoidosis | [80] | |
| SOD | EBC | low in healthy subjects, MnSOD upregulated in sarcoidosis | downregulated in patients treated with steroids/immunosuppressants | [34,80] |
5. Antioxidant Interventions in Sarcoidosis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gerke, A.K.; Judson, M.A.; Cozier, Y.C.; Culver, D.A.; Koth, L.L. Disease Burden and Variability in Sarcoidosis. Ann. ATS 2017, 14 (Suppl. 6), S421–S428. [Google Scholar] [CrossRef]
- Zissel, G.; Müller-Quernheim, J. Cellular Players in the Immunopathogenesis of Sarcoidosis. Clin. Chest Med. 2015, 36, 549–560. [Google Scholar] [CrossRef]
- Schupp, J.C.; Vukmirovic, M.; Kaminski, N.; Prasse, A. Transcriptome Profiles in Sarcoidosis and Their Potential Role in Disease Prediction. Curr. Opin. Pulm. Med. 2017, 23, 487–492. [Google Scholar] [CrossRef]
- Maier, L.A.; Crouser, E.D.; Martin, W.J.; Eu, J. Executive Summary of the NHLBI Workshop Report: Leveraging Current Scientific Advancements to Understand Sarcoidosis Variability and Improve Outcomes. Ann. ATS 2017, 14 (Suppl. 6), S415–S420. [Google Scholar] [CrossRef]
- Sotgia, S.; Paliogiannis, P.; Sotgiu, E.; Mellino, S.; Zinellu, E.; Fois, A.G.; Pirina, P.; Carru, C.; Mangoni, A.A.; Zinellu, A. Systematic Review and Meta-Analysis of the Blood Glutathione Redox State in Chronic Obstructive Pulmonary Disease. Antioxidants 2020, 9, 1146. [Google Scholar] [CrossRef]
- Paliogiannis, P.; Fois, A.G.; Collu, C.; Bandinu, A.; Zinellu, E.; Carru, C.; Pirina, P.; Mangoni, A.A.; Zinellu, A. Oxidative Stress-Linked Biomarkers in Idiopathic Pulmonary Fibrosis: A Systematic Review and Meta-Analysis. Biomark. Med. 2018, 12, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Young, I.S. Antioxidants in Health and Disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef]
- Van der Vliet, A. NADPH Oxidases in Lung Biology and Pathology: Host Defense Enzymes, and More. Free Radic. Biol. Med. 2008, 44, 938–955. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, C.-H.; Ryu, J.-H.; Kim, M.-J.; Park, C.Y.; Lee, J.M.; Holtzman, M.J.; Yoon, J.-H. Reactive Oxygen Species Induce Antiviral Innate Immune Response through IFN-λ Regulation in Human Nasal Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2013, 49, 855–865. [Google Scholar] [CrossRef]
- Black, S.M. New Insights into Acute Lung Injury. Vasc. Pharmacol. 2010, 52, 171–174. [Google Scholar] [CrossRef]
- Saleh, D.; Barnes, P.J.; Giaid, A. Increased Production of the Potent Oxidant Peroxynitrite in the Lungs of Patients with Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 1997, 155, 1763–1769. [Google Scholar] [CrossRef]
- Auten, R.L.; Davis, J.M. Oxygen Toxicity and Reactive Oxygen Species: The Devil Is in the Details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef]
- Shankar, K.; Mehendale, H.M. Oxidative Stress. In Encyclopedia of Toxicology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 735–737. [Google Scholar] [CrossRef]
- Cabello-Verrugio, C.; Vilos, C.; Rodrigues-Diez, R.; Estrada, L. Oxidative Stress in Disease and Aging: Mechanisms and Therapies 2018. Oxidative Med. Cell. Longev. 2018, 2018, 2835189. [Google Scholar] [CrossRef]
- Huang, D.; Fang, F.; Xu, F. Hyperoxia Induces Inflammation and Regulates Cytokine Production in Alveolar Epithelium through TLR2/4-NF-ΚB-Dependent Mechanism. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1399–1410. [Google Scholar]
- Habib, M.P.; Clements, N.C.; Garewal, H.S. Cigarette Smoking and Ethane Exhalation in Humans. Am. J. Respir. Crit. Care Med. 1995, 151, 1368–1372. [Google Scholar] [CrossRef] [PubMed]
- Wiegman, C.H.; Li, F.; Clarke, C.J.; Jazrawi, E.; Kirkham, P.; Barnes, P.J.; Adcock, I.M.; Chung, K.F. A Comprehensive Analysis of Oxidative Stress in the Ozone-Induced Lung Inflammation Mouse Model. Clin. Sci. 2014, 126, 425–440. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, J.; Jiang, R.; Song, W. Rat Lung Response to Ozone and Fine Particulate Matter (PM 2.5) Exposures: Rat Lung Response to Ozone and Fine Pm 2.5 Exposures. Environ. Toxicol. 2015, 30, 343–356. [Google Scholar] [CrossRef]
- Rada, B.; Lekstrom, K.; Damian, S.; Dupuy, C.; Leto, T.L. The Pseudomonas Toxin Pyocyanin Inhibits the Dual Oxidase-Based Antimicrobial System as It Imposes Oxidative Stress on Airway Epithelial Cells. J. Immunol. 2008, 181, 4883–4893. [Google Scholar] [CrossRef] [PubMed]
- Holguin, F. Oxidative Stress in Airway Diseases. Ann. ATS 2013, 10, S150–S157. [Google Scholar] [CrossRef]
- Marseglia, L.; D’Angelo, G.; Granese, R.; Falsaperla, R.; Reiter, R.J.; Corsello, G.; Gitto, E. Role of Oxidative Stress in Neonatal Respiratory Distress Syndrome. Free Radic. Biol. Med. 2019, 142, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M. The Dual Role of Oxidative Stress in Lung Cancer. In Oxidative Stress in Lung Diseases; Chakraborti, S., Chakraborti, T., Das, S.K., Chattopadhyay, D., Eds.; Springer: Singapore, 2019; pp. 99–113. [Google Scholar] [CrossRef]
- Fois, A.G.; Paliogiannis, P.; Sotgia, S.; Mangoni, A.A.; Zinellu, E.; Pirina, P.; Carru, C.; Zinellu, A. Evaluation of Oxidative Stress Biomarkers in Idiopathic Pulmonary Fibrosis and Therapeutic Applications: A Systematic Review. Respir. Res. 2018, 19, 51. [Google Scholar] [CrossRef]
- Girgis, R.E.; Basha, M.A.; Maliarik, M.; Popovich, J.; Iannuzzi, M.C. Cytokines in the Bronchoalveolar Lavage Fluid of Patients with Active Pulmonary Sarcoidosis. Am. J. Respir. Crit Care. Med. 1995, 152, 71–75. [Google Scholar] [CrossRef]
- Chen, E.S.; Moller, D.R. Sarcoidosis—Scientific Progress and Clinical Challenges. Nat. Rev. Rheumatol. 2011, 7, 457–467. [Google Scholar] [CrossRef]
- Richmond, B.W.; Ploetze, K.; Isom, J.; Chambers-Harris, I.; Braun, N.A.; Taylor, T.; Abraham, S.; Mageto, Y.; Culver, D.A.; Oswald-Richter, K.A.; et al. Sarcoidosis Th17 Cells Are ESAT-6 Antigen Specific but Demonstrate Reduced IFN-γ Expression. J. Clin. Immunol. 2013, 33, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Cassatella, M.; Berton, G.; Agostini, C.; Zambello, R. Generation of Superoxide Anion by Alveolar Macrophages in Sarcoidosis: Evidence for the Activation of the Oxygen Metabolism in Patients with High-Intensity Alveolitis. Immunology 1989, 66, 451–458. [Google Scholar]
- Nathan, C.F.; Murray, H.W.; Wiebe, M.E.; Rubin, B.Y. Identification of Interferon-Gamma as the Lymphokine That Activates Human Macrophage Oxidative Metabolism and Antimicrobial Activity. J. Exp. Med. 1983, 158, 670–689. [Google Scholar] [CrossRef] [PubMed]
- Fels, A.O.; Nathan, C.F.; Cohn, Z.A. Hydrogen Peroxide Release by Alveolar Macrophages from Sarcoid Patients and by Alveolar Macrophages from Normals after Exposure to Recombinant Interferons Alpha A, Beta, and Gamma and 1,25-Dihydroxyvitamin D3. J. Clin. Investig. 1987, 80, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Lenz, A.-G.; Costabel, U.; Maier, K.L. Oxidized BAL Fluid Proteins in Patients with Interstitial Lung Diseases. Eur. Respir. J. 1996, 9, 307–312. [Google Scholar] [CrossRef]
- Zinellu, E.; Zinellu, A.; Fois, A.G.; Fois, S.S.; Piras, B.; Carru, C.; Pirina, P. Reliability and Usefulness of Different Biomarkers of Oxidative Stress in Chronic Obstructive Pulmonary Disease. Oxidative Med. Cell. Longev. 2020, 2020, 4982324. [Google Scholar] [CrossRef]
- Koutsokera, A.; Papaioannou, A.I.; Malli, F.; Kiropoulos, T.S.; Katsabeki, A.; Kerenidi, T.; Gourgoulianis, K.I.; Daniil, Z.D. Systemic Oxidative Stress in Patients with Pulmonary Sarcoidosis. Pulm. Pharmacol. Ther. 2009, 22, 603–607. [Google Scholar] [CrossRef]
- Ivanisevic, J.; Vekic, J.; Zeljkovic, A.; Stefanovic, A.; Kotur-Stevuljevic, J.; Spasojevic-Kalimanovska, V.; Spasic, S.; Vucinic-Mihailovic, V.; Videnovic-Ivanov, J.; Jelic-Ivanovic, Z. Associations of Lipoprotein Subclasses and Oxidative Stress Status in Pulmonary and Pulmonary plus Extrapulmonary Sarcoidosis. Sarcoidosis Vasc. Diffus. Lung Dis. 2018, 35, 198–205. [Google Scholar] [CrossRef]
- Ivanišević, J.; Kotur-Stevuljević, J.; Stefanović, A.; Jelić-Ivanović, Z.; Spasić, S.; Videnović-Ivanov, J.; Vučinić-Mihailović, V.; Ilić, J. Dyslipidemia and Oxidative Stress in Sarcoidosis Patients. Clin. Biochem. 2012, 45, 677–682. [Google Scholar] [CrossRef]
- Niki, E. Lipid Peroxidation: Physiological Levels and Dual Biological Effects. Free Radic. Biol. Med. 2009, 47, 469–484. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A Review of Recent Studies on Malondialdehyde as Toxic Molecule and Biological Marker of Oxidative Stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Janero, D.R.; Burghardt, B. Analysis of Cardiac Membrane Phospholipid Peroxidation Kinetics as Malondialdehyde: Nonspecificity of Thiobarbituric Acid-Reactivity. Lipids 1988, 23, 452–458. [Google Scholar] [CrossRef]
- Uzun, H.; Yanardag, H.; Gelisgen, R.; Genc, H.; Uygun, S.; Vehid, S.; Karter, Y.; Demirci, S. Levels of Paraoxonase, an Index of Antioxidant Defense, in Patients with Active Sarcoidosis. Curr. Med. Res. Opin. 2008, 24, 1651–1657. [Google Scholar] [CrossRef] [PubMed]
- Viigimaa, M.; Abina, J.; Zemtsovskaya, G.; Tikhaze, A.; Konovalova, G.; Kumskova, E.; Lankin, V. Malondialdehyde-Modified Low-Density Lipoproteins as Biomarker for Atherosclerosis. Blood Press. 2010, 19, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Ansarin, K.; Khoubnasabjafari, M.; Jouyban, A. Reliability of Malondialdehyde as a Biomarker of Oxidative Stress in Psychological Disorders. Bioimpacts 2017, 5, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Montuschi, P.; Barnes, P.; Jackson Roberts, L. Insights into Oxidative Stress: The Isoprostanes. CMC 2007, 14, 703–717. [Google Scholar] [CrossRef]
- Halliwell, B. Lipid Peroxidation, Antioxidants and Cardiovascular Disease: How Should We Move Forward? Cardiovasc. Res. 2000, 47, 410–418. [Google Scholar] [CrossRef]
- Morrow, J.D.; Hill, K.E.; Burk, R.F.; Nammour, T.M.; Badr, K.F.; Roberts, L.J. A Series of Prostaglandin F2-like Compounds Are Produced in Vivo in Humans by a Non-Cyclooxygenase, Free Radical-Catalyzed Mechanism. Proc. Natl. Acad. Sci. USA 1990, 87, 9383–9387. [Google Scholar] [CrossRef] [PubMed]
- Morrow, J.D.; Frei, B.; Longmire, A.W.; Gaziano, J.M.; Lynch, S.M.; Shyr, Y.; Strauss, W.E.; Oates, J.A.; Roberts, L.J. Increase in Circulating Products of Lipid Peroxidation (F 2 -Isoprostanes) in Smokers—Smoking as a Cause of Oxidative Damage. N. Engl. J. Med. 1995, 332, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Montuschi, P.; Toni, G.C.; Paredi, P.; Pantelidis, P.; du Bois, R.M.; Kharitonov, S.A.; Barnes, P.J. 8-Isoprostane as a Biomarker of Oxidative Stress in Interstitial Lung Diseases. Am. J. Respir. Crit. Care Med. 1998, 158, 1524–1527. [Google Scholar] [CrossRef]
- Montuschi, P.; Collins, J.V.; Ciabattoni, G.; Lazzeri, N.; Corradi, M.; Kharitonov, S.A.; Barnes, P.J. Exhaled 8-Isoprostane as an In Vivo Biomarker of Lung Oxidative Stress in Patients with COPD and Healthy Smokers. Am. J. Respir. Crit. Care Med. 2000, 162, 1175–1177. [Google Scholar] [CrossRef]
- Montuschi, P.; Corradi, M.; Ciabattoni, G.; Nightingale, J.; Kharitonov, S.A.; Barnes, P.J. Increased 8-Isoprostane, a Marker of Oxidative Stress, in Exhaled Condensate of Asthma Patients. Am. J. Respir. Crit. Care Med. 1999, 160, 216–220. [Google Scholar] [CrossRef]
- Peel, A.M.; Crossman-Barnes, C.-J.; Tang, J.; Fowler, S.J.; Davies, G.A.; Wilson, A.M.; Loke, Y.K. Biomarkers in Adult Asthma: A Systematic Review of 8-Isoprostane in Exhaled Breath Condensate. J. Breath Res. 2017, 11, 016011. [Google Scholar] [CrossRef][Green Version]
- Carpenter, C.T.; Price, P.V.; Christman, B.W. Exhaled Breath Condensate Isoprostanes Are Elevated in Patients with Acute Lung Injury or ARDS. Chest 1998, 114, 1653–1659. [Google Scholar] [CrossRef]
- Psathakis, K.; Mermigkis, D.; Papatheodorou, G.; Loukides, S.; Panagou, P.; Polychronopoulos, V.; Siafakas, N.M.; Bouros, D. Exhaled Markers of Oxidative Stress in Idiopathic Pulmonary Fibrosis. Eur. J. Clin. Investig. 2006, 36, 362–367. [Google Scholar] [CrossRef]
- Malli, F.; Bardaka, F.; Tsilioni, I.; Karetsi, E.; Gourgoulianis, K.I.; Daniil, Z. 8-Isoprostane Levels in Serum and Bronchoalveolar Lavage in Idiopathic Pulmonary Fibrosis and Sarcoidosis. Food Chem. Toxicol. 2013, 61, 160–163. [Google Scholar] [CrossRef]
- Psathakis, K.; Papatheodorou, G.; Plataki, M.; Panagou, P.; Loukides, S.; Siafakas, N.M.; Bouros, D. 8-Isoprostane, a Marker of Oxidative Stress, Is Increased in the Expired Breath Condensate of Patients with Pulmonary Sarcoidosis. Chest 2004, 125, 1005–1011. [Google Scholar] [CrossRef]
- Piotrowski, W.J.; Antczak, A.; Marczak, J.; Nawrocka, A.; Kurmanowska, Z.; Górski, P. Eicosanoids in Exhaled Breath Condensate and BAL Fluid of Patients with Sarcoidosis. Chest 2007, 132, 589–596. [Google Scholar] [CrossRef]
- Pauling, L.; Robinson, A.B.; Teranishi, R.; Cary, P. Quantitative Analysis of Urine Vapor and Breath by Gas-Liquid Partition Chromatography. Proc. Natl. Acad. Sci. USA 1971, 68, 2374–2376. [Google Scholar] [CrossRef]
- Hakim, M.; Broza, Y.Y.; Barash, O.; Peled, N.; Phillips, M.; Amann, A.; Haick, H. Volatile Organic Compounds of Lung Cancer and Possible Biochemical Pathways. Chem. Rev. 2012, 112, 5949–5966. [Google Scholar] [CrossRef]
- Amann, A.; de Costello, B.L.; Miekisch, W.; Schubert, J.; Buszewski, B.; Pleil, J.; Ratcliffe, N.; Risby, T. The Human Volatilome: Volatile Organic Compounds (VOCs) in Exhaled Breath, Skin Emanations, Urine, Feces and Saliva. J. Breath Res. 2014, 8, 034001. [Google Scholar] [CrossRef]
- Mazzone, P.J. Analysis of Volatile Organic Compounds in the Exhaled Breath for the Diagnosis of Lung Cancer. J. Thorac. Oncol. 2008, 3, 774–780. [Google Scholar] [CrossRef]
- Kanoh, S.; Kobayashi, H.; Motoyoshi, K. Exhaled Ethane. Chest 2005, 128, 2387–2392. [Google Scholar] [CrossRef]
- Hayakawa, H.; Hofer, A.; Thelander, L.; Kitajima, S.; Cai, Y.; Oshiro, S.; Yakushiji, H.; Nakabeppu, Y.; Kuwano, M.; Sekiguchi, M. Metabolic Fate of Oxidized Guanine Ribonucleotides in Mammalian Cells. Biochemistry 1999, 38, 3610–3614. [Google Scholar] [CrossRef]
- Kobayashi, S.; Myoren, T.; Oda, S.; Inari, M.; Ishiguchi, H.; Murakami, W.; Fukuda, M.; Tanaka, T.; Okuda, S.; Nao, T.; et al. Urinary 8-Hydroxy-2′-Deoxyguanosine as a Novel Biomarker of Inflammatory Activity in Patients with Cardiac Sarcoidosis. Int. J. Cardiol. 2015, 190, 319–328. [Google Scholar] [CrossRef]
- Myoren, T.; Kobayashi, S.; Oda, S.; Nanno, T.; Ishiguchi, H.; Murakami, W.; Okuda, S.; Okada, M.; Takemura, G.; Suga, K.; et al. An Oxidative Stress Biomarker, Urinary 8-Hydroxy-2′-Deoxyguanosine, Predicts Cardiovascular-Related Death after Steroid Therapy for Patients with Active Cardiac Sarcoidosis. Int. J. Cardiol. 2016, 212, 206–213. [Google Scholar] [CrossRef]
- Piantadosi, C.A.; Suliman, H.B. Mitochondrial Dysfunction in Lung Pathogenesis. Annu. Rev. Physiol. 2017, 79, 495–515. [Google Scholar] [CrossRef]
- Van der Vliet, A.; Janssen-Heininger, Y.M.W.; Anathy, V. Oxidative Stress in Chronic Lung Disease: From Mitochondrial Dysfunction to Dysregulated Redox Signaling. Mol. Asp. Med. 2018, 63, 59–69. [Google Scholar] [CrossRef]
- Mochizuki, I.; Kubo, K.; Honda, T. Relationship between Mitochondria and the Development of Specific Lipid Droplets in Capillary Endothelial Cells of the Respiratory Tract in Patients with Sarcoidosis. Mitochondrion 2011, 11, 601–606. [Google Scholar] [CrossRef]
- Daniil, Z.; Kotsiou, O.S.; Grammatikopoulos, A.; Peletidou, S.; Gkika, H.; Malli, F.; Antoniou, K.; Vasarmidi, E.; Mamuris, Z.; Gourgoulianis, K.; et al. Detection of Mitochondrial Transfer RNA (Mt-TRNA) Gene Mutations in Patients with Idiopathic Pulmonary Fibrosis and Sarcoidosis. Mitochondrion 2018, 43, 43–52. [Google Scholar] [CrossRef]
- Bhargava, M.; Viken, K.J.; Barkes, B.; Griffin, T.J.; Gillespie, M.; Jagtap, P.D.; Sajulga, R.; Peterson, E.J.; Dincer, H.E.; Li, L.; et al. Novel Protein Pathways in Development and Progression of Pulmonary Sarcoidosis. Sci. Rep. 2020, 10, 13282. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef]
- Boots, A.W.; Drent, M.; Swennen, E.L.R.; Moonen, H.J.J.; Bast, A.; Haenen, G.R.M.M. Antioxidant Status Associated with Inflammation in Sarcoidosis: A Potential Role for Antioxidants. Respir. Med. 2009, 103, 364–372. [Google Scholar] [CrossRef]
- Hopkins, B.L.; Neumann, C.A. Redoxins as Gatekeepers of the Transcriptional Oxidative Stress Response. Redox Biol. 2019, 21, 101104. [Google Scholar] [CrossRef]
- Hanschmann, E.-M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, Glutaredoxins, and Peroxiredoxins—Molecular Mechanisms and Health Significance: From Cofactors to Antioxidants to Redox Signaling. Antioxid. Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef]
- Ogata, F.T.; Branco, V.; Vale, F.F.; Coppo, L. Glutaredoxin: Discovery, Redox Defense and Much More. Redox Biol. 2021, 43, 101975. [Google Scholar] [CrossRef]
- Koura, T. Expression of Thioredoxin in Granulomas of Sarcoidosis: Possible Role in the Development of T Lymphocyte Activation. Thorax 2000, 55, 755–761. [Google Scholar] [CrossRef]
- Iwata, S.; Hori, T.; Sato, N.; Ueda-Taniguchi, Y.; Yamabe, T.; Nakamura, H.; Masutani, H.; Yodoi, J. Thiol-Mediated Redox Regulation of Lymphocyte Proliferation. Possible Involvement of Adult T Cell Leukemia-Derived Factor and Glutathione in Transferrin Receptor Expression. J. Immunol. 1994, 152, 5633–5642. [Google Scholar]
- Tiitto, L.; Kaarteenaho-Wiik, R.; Sormunen, R.; Holmgren, A.; Pääkkö, P.; Soini, Y.; Kinnula, V.L. Expression of the Thioredoxin System in Interstitial Lung Disease: Expression of the Thioredoxin System in Interstitial Lung Disease. J. Pathol. 2003, 201, 363–370. [Google Scholar] [CrossRef]
- Peltoniemi, M.; Kaarteenaho-Wiik, R.; Säily, M.; Sormunen, R.; Pääkkö, P.; Holmgren, A.; Soini, Y.; Kinnula, V.L. Expression of Glutaredoxin Is Highly Cell Specific in Human Lung and Is Decreased by Transforming Growth Factor-β in Vitro and in Interstitial Lung Diseases in Vivo. Hum. Pathol. 2004, 35, 1000–1007. [Google Scholar] [CrossRef]
- Celada, L.J.; Kropski, J.A.; Herazo-Maya, J.D.; Luo, W.; Creecy, A.; Abad, A.T.; Chioma, O.S.; Lee, G.; Hassell, N.E.; Shaginurova, G.I.; et al. PD-1 up-Regulation on CD4 + T Cells Promotes Pulmonary Fibrosis through STAT3-Mediated IL-17A and TGF-Β1 Production. Sci. Transl. Med. 2018, 10, eaar8356. [Google Scholar] [CrossRef]
- Arsalane, K.; Dubois, C.M.; Muanza, T.; Bégin, R.; Boudreau, F.; Asselin, C.; Cantin, A.M. Transforming Growth Factor- β 1 Is a Potent Inhibitor of Glutathione Synthesis in the Lung Epithelial Cell Line A549: Transcriptional Effect on the GSH Rate-Limiting Enzyme γ-Glutamylcysteine Synthetase. Am. J. Respir. Cell Mol. Biol. 1997, 17, 599–607. [Google Scholar] [CrossRef]
- Winternitz, M.C.; Meloy, C.R. On the occurrence of catalase in human tissues and its variations in diseases. J. Exp. Med. 1908, 10, 759–781. [Google Scholar] [CrossRef][Green Version]
- Glorieux, C.; Calderon, P.B. Catalase, a Remarkable Enzyme: Targeting the Oldest Antioxidant Enzyme to Find a New Cancer Treatment Approach. Biol. Chem. 2017, 398, 1095–1108. [Google Scholar] [CrossRef]
- Lakari, E.; Pääkkö, P.; Kinnula, V.L. Manganese Superoxide Dismutase, but Not CuZn Superoxide Dismutase, Is Highly Expressed in the Granulomas of Pulmonary Sarcoidosis and Extrinsic Allergic Alveolitis. Am. J. Respir. Crit. Care Med. 1998, 158, 589–596. [Google Scholar] [CrossRef]
- Lewis-Molock, Y.; Suzuki, K.; Taniguchi, N.; Nguyen, D.H.; Mason, R.J.; White, C.W. Lung Manganese Superoxide Dismutase Increases during Cytokine-Mediated Protection against Pulmonary Oxygen Toxicity in Rats. Am. J. Respir. Cell Mol. Biol. 1994, 10, 133–141. [Google Scholar] [CrossRef]
- Wong, G.; Goeddel, D. Induction of Manganous Superoxide Dismutase by Tumor Necrosis Factor: Possible Protective Mechanism. Science 1988, 242, 941–944. [Google Scholar] [CrossRef]
- Tsan, M.-F. Superoxide Dismutase and Pulmonary Oxygen Toxicity. Exp. Biol. Med. 1997, 214, 107–113. [Google Scholar] [CrossRef]
- Park, E.-M.; Park, Y.-M.; Gwak, Y.-S. Oxidative Damage in Tissues of Rats Exposed to Cigarette Smoke. Free Radic. Biol. Med. 1998, 25, 79–86. [Google Scholar] [CrossRef]
- Ghezzi, P. Role of Glutathione in Immunity and Inflammation in the Lung. Int. J. Gen. Med. 2011, 4, 105. [Google Scholar] [CrossRef]
- Behr, J.; Degenkolb, B.; Beinert, T.; Krombach, F.; Vogelmeier, C. Pulmonary Glutathione Levels in Acute Episodes of Farmer’s Lung. Am. J. Respir. Crit. Care Med. 2000, 161, 1968–1971. [Google Scholar] [CrossRef][Green Version]
- Cantin, A.M.; Hubbard, R.C.; Crystal, R.G. Glutathione Deficiency in the Epithelial Lining Fluid of the Lower Respiratory Tract in Idiopathic Pulmonary Fibrosis. Am. Rev. Respir. Dis. 1989, 139, 370–372. [Google Scholar] [CrossRef]
- Meyer, A.; Buhl, R.; Magnussen, H. The Effect of Oral N-Acetylcysteine on Lung Glutathione Levels in Idiopathic Pulmonary Fibrosis. Eur. Respir. J. 1994, 7, 431–436. [Google Scholar] [CrossRef]
- Pacht, E.R.; Timerman, A.P.; Lykens, M.G.; Merola, J. Deficiency of Alveolar Fluid Glutathione in Patients with Sepsis and the Adult Respiratory Distress Syndrome. Chest 1991, 100, 1397–1403. [Google Scholar] [CrossRef]
- Sadegh Soltan-Sharifi, M.; Mojtahedzadeh, M.; Najafi, A.; Reza Khajavi, M.; Reza Rouini, M.; Moradi, M.; Mohammadirad, A.; Abdollahi, M. Improvement by N-Acetylcysteine of Acute Respiratory Distress Syndrome through Increasing Intracellular Glutathione, and Extracellular Thiol Molecules and Anti-Oxidant Power: Evidence for Underlying Toxicological Mechanisms. Hum. Exp. Toxicol. 2007, 26, 697–703. [Google Scholar] [CrossRef]
- Rothkrantz-Kos, S.; Drent, M.; Vuil, H.; De Boer, M.; Bast, A.; Wouters, E.F.M.; Roos, D.; van Dieijen-Visser, M.P. Decreased Redox State in Red Blood Cells from Patients with Sarcoidosis. Sarcoidosis Vasc. Diffus. Lung Dis. 2002, 19, 114–120. [Google Scholar]
- García-Heredia, A.; Marsillach, J.; Rull, A.; Triguero, I.; Fort, I.; Mackness, B.; Mackness, M.; Shih, D.M.; Joven, J.; Camps, J. Paraoxonase-1 Inhibits Oxidized Low-Density Lipoprotein-Induced Metabolic Alterations and Apoptosis in Endothelial Cells: A Nondirected Metabolomic Study. Mediat. Inflamm. 2013, 2013, 156053. [Google Scholar] [CrossRef]
- Getz, G.S.; Reardon, C.A. Paraoxonase, a Cardioprotective Enzyme: Continuing Issues. Curr. Opin. Lipidol. 2004, 15, 261–267. [Google Scholar] [CrossRef]
- Mackness, B.; Durrington, P.N.; Mackness, M.I. Human Serum Paraoxonase. Gen. Pharmacol. Vasc. Syst. 1998, 31, 329–336. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Melnichenko, A.A.; Orekhov, A.N.; Bobryshev, Y.V. Paraoxonase and Atherosclerosis-Related Cardiovascular Diseases. Biochimie 2017, 132, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Rosenblat, M.; Bisgaier, C.L.; Newton, R.S.; Primo-Parmo, S.L.; La Du, B.N. Paraoxonase Inhibits High-Density Lipoprotein Oxidation and Preserves Its Functions. A Possible Peroxidative Role for Paraoxonase. J. Clin. Investig. 1998, 101, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, L.; Hernández, A.F.; López-Caballero, J.J.; Gil, F.; Pla, A. Immunohistochemical Evidence for the Expression and Induction of Paraoxonase in Rat Liver, Kidney, Lung and Brain Tissue. Implications for Its Physiological Role. Chem.-Biol. Interact. 2001, 137, 123–137. [Google Scholar] [CrossRef]
- Sarioglu, N.; Bilen, C.; Cevik, C.; Gencer, N. Paraoxonase Activity and Phenotype Distribution in Patients with Chronic Obstructive Pulmonary Disease. Eurasian J. Med. 2020, 52, 161–165. [Google Scholar] [CrossRef]
- Grdić Rajković, M.; Popović-Grle, S.; Vukić Dugac, A.; Rogić, D.; Rako, I.; Radić Antolić, M.; Beriša, M.; Rumora, L. PON1 Gene Polymorphisms in Patients with Chronic Obstructive Pulmonary Disease. J. Clin. Pathol. 2018, 71, 963–970. [Google Scholar] [CrossRef]
- Chen, W.; Xie, Z.; Wang, X.; Zhao, J.; Hu, Q.; Chen, Y.; Gao, W.; Liu, Y. Influences of PON1 on Airway Inflammation and Remodeling in Bronchial Asthma. J. Cell. Biochem. 2018, 119, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Aldonza, M.B.D.; Son, Y.S.; Sung, H.-J.; Ahn, J.M.; Choi, Y.-J.; Kim, Y.-I.; Cho, S.; Cho, J.-Y. Paraoxonase-1 (PON1) Induces Metastatic Potential and Apoptosis Escape via Its Antioxidative Function in Lung Cancer Cells. Oncotarget 2017, 8, 42817–42835. [Google Scholar] [CrossRef]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an Antioxidant and Disulphide Breaking Agent: The Reasons Why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef]
- Cu, A.; Ye, Q.; Sarria, R.; Nakamura, S.; Guzman, J.; Costabel, U. N-Acetylcysteine Inhibits TNF-Alpha, STNFR, and TGF-Beta1 Release by Alveolar Macrophages in Idiopathic Pulmonary Fibrosis in Vitro. Sarcoidosis Vasc. Diffus. Lung Dis. 2009, 26, 147–154. [Google Scholar]
- Martinez, F.J.; de Andrade, J.A. The Idiopathic Pulmonary Fibrosis Clinical Research Network. Randomized Trial of Acetylcysteine in Idiopathic Pulmonary Fibrosis. N. Engl. J. Med. 2014, 370, 2093–2101. [Google Scholar] [CrossRef] [PubMed]
- Niewoehner, D.; Johnson, K.; McEvoy, C.; Naqvi, S.; Wendt, C.; Reilkoff, R.; Kunisaki, K.; Wetherbee, E.; Nelson, D.; Tirouvanziam, R. High-Dose Oral N-Acetylcysteine Fails to Improve Respiratory Health Status in Patients with Chronic Obstructive Pulmonary Disease and Chronic Bronchitis: A Randomized, Placebo-Controlled Trial. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 799. [Google Scholar] [CrossRef][Green Version]
- Hamzeh, N.; Li, L.; Barkes, B.; Huang, J.; Canono, B.; Gillespie, M.; Maier, L.; Day, B. The Effect of an Oral Anti-Oxidant, N-Acetyl-Cysteine, on Inflammatory and Oxidative Markers in Pulmonary Sarcoidosis. Respir. Med. 2016, 112, 106–111. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef] [PubMed]
- Boots, A.W.; Haenen, G.R.M.M.; Bast, A. Health Effects of Quercetin: From Antioxidant to Nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Nair, M.P.; Mahajan, S.; Reynolds, J.L.; Aalinkeel, R.; Nair, H.; Schwartz, S.A.; Kandaswami, C. The Flavonoid Quercetin Inhibits Proinflammatory Cytokine (Tumor Necrosis Factor Alpha) Gene Expression in Normal Peripheral Blood Mononuclear Cells via Modulation of the NF-Κβ System. Clin. Vaccine Immunol. 2006, 13, 319–328. [Google Scholar] [CrossRef]
- Boots, A.W.; Drent, M.; de Boer, V.C.J.; Bast, A.; Haenen, G.R.M.M. Quercetin Reduces Markers of Oxidative Stress and Inflammation in Sarcoidosis. Clin. Nutr. 2011, 30, 506–512. [Google Scholar] [CrossRef]
- Rossman, M.D.; Newman, L.S.; Baughman, R.P.; Teirstein, A.; Weinberger, S.E.; Miller, W.; Sands, B.E. A Double-Blinded, Randomized, Placebo-Controlled Trial of Infliximab in Subjects with Active Pulmonary Sarcoidosis. Sarcoidosis Vasc. Diffus. Lung Dis. 2006, 23, 201–208. [Google Scholar]
- Hostettler, K.E.; Studler, U.; Tamm, M.; Brutsche, M.H. Long-Term Treatment with Infliximab in Patients with Sarcoidosis. Respiration 2012, 83, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Montes, A.; Ruiz-Corro, L.; López-Reyes, A.; Castrejón-Gómez, E.; Armendáriz-Borunda, J. Potent Antioxidant Role of Pirfenidone in Experimental Cirrhosis. Eur. J. Pharmacol. 2008, 595, 69–77. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fois, S.S.; Canu, S.; Fois, A.G. The Role of Oxidative Stress in Sarcoidosis. Int. J. Mol. Sci. 2021, 22, 11712. https://doi.org/10.3390/ijms222111712
Fois SS, Canu S, Fois AG. The Role of Oxidative Stress in Sarcoidosis. International Journal of Molecular Sciences. 2021; 22(21):11712. https://doi.org/10.3390/ijms222111712
Chicago/Turabian StyleFois, Sara Solveig, Sara Canu, and Alessandro Giuseppe Fois. 2021. "The Role of Oxidative Stress in Sarcoidosis" International Journal of Molecular Sciences 22, no. 21: 11712. https://doi.org/10.3390/ijms222111712
APA StyleFois, S. S., Canu, S., & Fois, A. G. (2021). The Role of Oxidative Stress in Sarcoidosis. International Journal of Molecular Sciences, 22(21), 11712. https://doi.org/10.3390/ijms222111712