
Plant Gravitropism and Signal Conversion under a Stress Environment of Altered Gravity

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. History of Plant Cultivation Systems in Outer Space
3. How Plants Sense Gravity and Change under Stress Environments of Altered Gravity
3.1. How Plant Sensing Gravity
3.2. Transduction of Gravity Signal and Auxin as a Signal
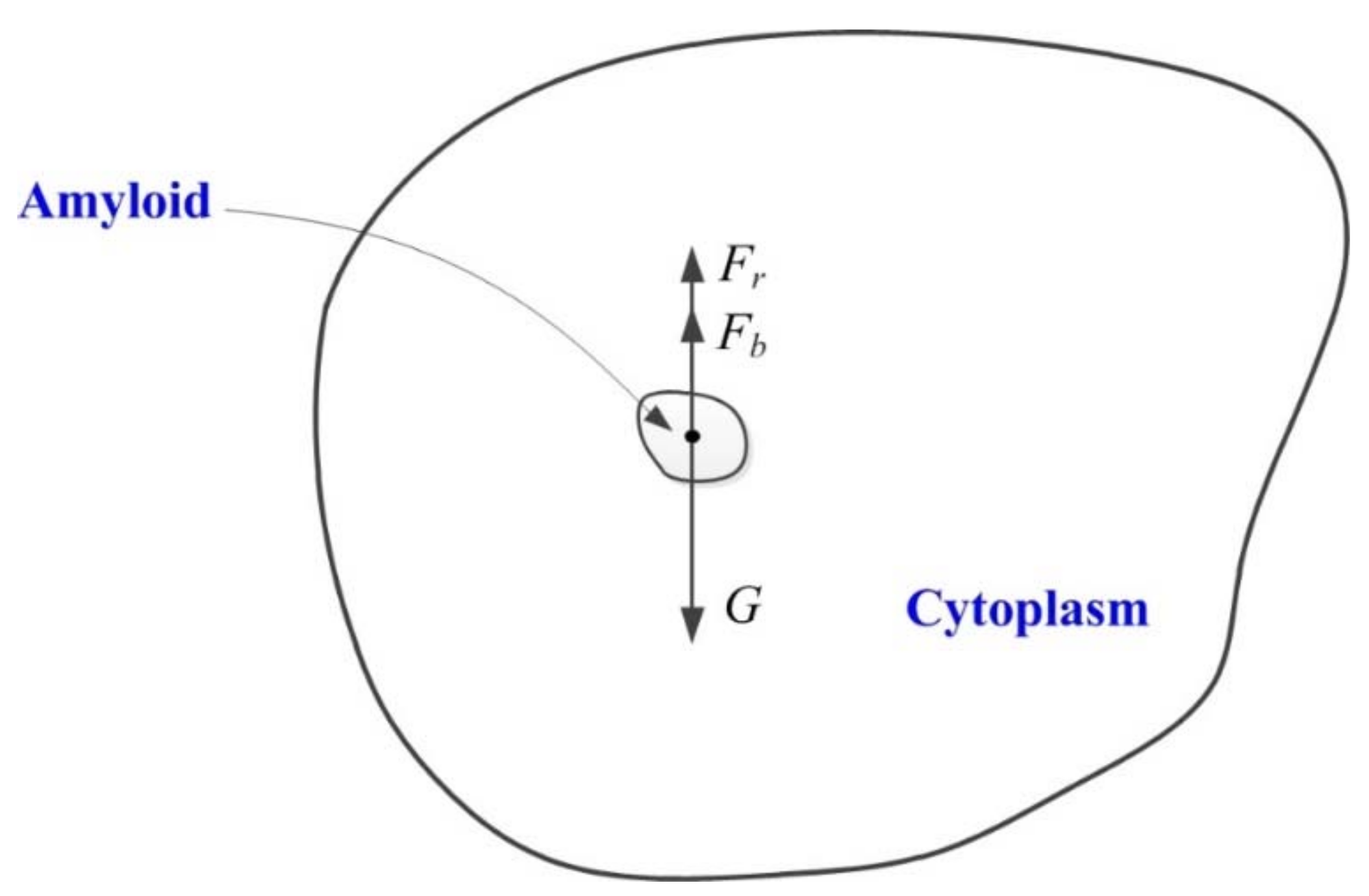
3.3. The Dynamic Model of Amyloplast Sedimentation
3.4. The Lateral Root Sense Altered Gravity
3.5. The Plant Gravitropism Related to Phototropism
4. The Plant Experiments under Simulated and Real Altered Gravity Environment
4.1. The Plant Experiments under Simulated Microgravity (0 g), Moon Gravity (0.17 g) and Mars Gravity (0.38 g) Environment
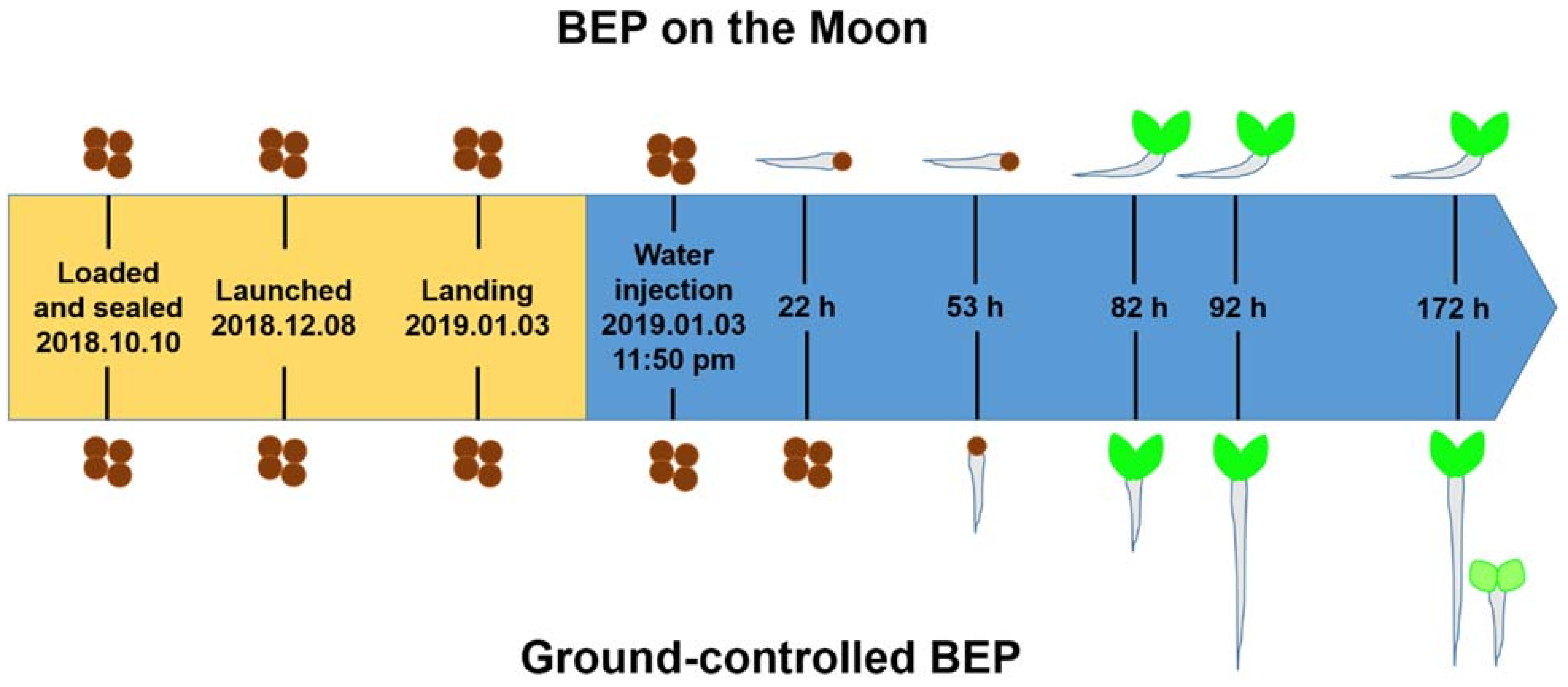
4.2. The Plant Experiment under a Real Moon Gravity Environment (0.17 g)
5. Concluding Remarks and Future Perspectives: Mars and Lunar Base Construction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature 2001, 409, 1092–1101. [Google Scholar] [CrossRef]
- Tazi, L.; Breakwell, D.P.; Harker, A.R.; Crandall, K.A. Life in extreme environments: Microbial diversity in Great Salt Lake, Utah. Extrem. Life Under Extrem. Cond. 2014, 18, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Tych, K.M.; Hoffmann, T.; Batchelor, M.; Hughes, M.L.; Kendrick, K.E.; Walsh, D.L.; Wilson, M.; Brockwell, D.J.; Dougan, L. Life in extreme environments: Single molecule force spectroscopy as a tool to explore proteins from extremophilic organisms. Biochem. Soc. Trans. 2015, 43, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Ruyters, G.; Braun, M. Plant biology in space: Recent accomplishments and recommendations for future research. Plant Biol. 2014, 16, 4–11. [Google Scholar] [CrossRef]
- Bamsey, M.; Graham, T.; Thompson, C.; Berinstain, A.; Scott, A.; Dixon, M. Ion-specific nutrient management in closed systems: The necessity for ion-selective sensors in terrestrial and space-based agriculture and water management systems. Sensors 2012, 12, 13349–13392. [Google Scholar] [CrossRef] [PubMed]
- Vandenbrink, J.P.; Kiss, J.Z.; Herranz, R.; Medina, F.J. Light and gravity signals synergize in modulating plant development. Front. Plant Sci. 2014, 5, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbrink, J.P.; Kiss, J.Z. Space, the final frontier: A critical review of recent experiments performed in microgravity. Plant Sci. Int. J. Exp. Plant Biol. 2016, 243, 115–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamal, K.Y.; Herranz, R.; van Loon, J.; Medina, F.J. Simulated microgravity, Mars gravity, and 2g hypergravity affect cell cycle regulation, ribosome biogenesis, and epigenetics in Arabidopsis cell cultures. Sci. Rep. 2018, 8, 6424. [Google Scholar] [CrossRef] [Green Version]
- Kiss, J.Z. Plant biology in reduced gravity on the Moon and Mars. Plant Biol. 2014, 16 (Suppl. 1), 12–17. [Google Scholar] [CrossRef]
- Wamelink, G.W.; Frissel, J.Y.; Krijnen, W.H.; Verwoert, M.R.; Goedhart, P.W. Can plants grow on Mars and the moon: A growth experiment on Mars and moon soil simulants. PLoS ONE 2014, 9, e103138. [Google Scholar] [CrossRef]
- Gibney, E. How To Build a Moon Base Researchers are ramping up plans for livimg on the Moon. Nature 2018, 562, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Wolff, S.A.; Coelho, L.H.; Karoliussen, I.; Jost, A.I. Effects of the Extraterrestrial Environment on Plants: Recommendations for Future Space Experiments for the MELiSSA Higher Plant Compartment. Life 2014, 4, 189–204. [Google Scholar] [CrossRef]
- Vernikos, J.; Walter, N.; Worms, J.C.; Blanc, S. THESEUS: The European research priorities for human exploration of space. NPJ Microgravity 2016, 2, 16034. [Google Scholar] [CrossRef] [Green Version]
- Zabel, P.; Bamsey, M.; Schubert, D.; Tajmar, M. Review and analysis of over 40 years of space plant growth systems. Life Sci. Space Res. 2016, 10, 1–16. [Google Scholar] [CrossRef]
- Paul, A.L.; Amalfitano, C.E.; Ferl, R.J. Plant growth strategies are remodeled by spaceflight. BMC Plant Biol. 2012, 12, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, R.C.; Crabb, T.M. Biomass production system (BPS) plant growth unit. Adv. Space Res. 2000, 26, 289–298. [Google Scholar] [CrossRef]
- Paul, A.L.; Bamsey, M.; Berinstain, A.; Braham, S.; Neron, P.; Murdoch, T.; Graham, T.; Ferl, R.J. Deployment of a Prototype Plant GFP Imager at the Arthur Clarke Mars Greenhouse of the Haughton Mars Project. Sensors 2008, 8, 2762–2773. [Google Scholar] [CrossRef]
- Bingham, G.E.; Jones, S.B.; Or, D.; Podolski, I.G.; Levinskikh, M.A.; Sytchov, V.N.; Ivanova, T.; Kostov, P.; Sapunova, S.; Dandolov, I.; et al. Microgravity effects on water supply and substrate properties in porous matrix root support systems. Acta Astronaut. 2000, 47, 839–848. [Google Scholar] [CrossRef]
- Stutte, G.W.; Monje, O.; Goins, G.D.; Tripathy, B.C. Microgravity effects on thylakoid, single leaf, and whole canopy photosynthesis of dwarf wheat. Planta 2005, 223, 46–56. [Google Scholar] [CrossRef]
- Hamatani, S.; Uetake, Y.; Karahara, I.; Masuda, K.; Kamisaka, S.; Hoson, T.; Wakabayashi, K.; Soga, K.; Nishitani, K.; Goto, N.; et al. Examination of growth environment for a long-term growth experiment of Arabidopsis thaliana L. on International Space Station. Uchu Seibutsu Kagaku 2001, 15, 262–263. [Google Scholar]
- Link, B.M.; Durst, S.J.; Zhou, W.; Stankovic, B. Seed-to-seed growth of Arabidopsis thaliana on the International Space Station. Adv. Space Res. 2003, 31, 2237–2243. [Google Scholar] [CrossRef]
- Driss-Ecole, D.; Legue, V.; Carnero-Diaz, E.; Perbal, G. Gravisensitivity and automorphogenesis of lentil seedling roots grown on board the International Space Station. Physiol. Plant. 2008, 134, 191–201. [Google Scholar] [CrossRef]
- Porterfield, D.M.; Neichitailo, G.S.; Mashinski, A.L.; Musgrave, M.E. Spaceflight hardware for conducting plant growth experiments in space: The early years 1960–2000. Adv. Space Res.-Ser. 2003, 31, 183–193. [Google Scholar] [CrossRef]
- Jost, A.I.; Hoson, T.; Iversen, T.H. The Utilization of Plant Facilities on the International Space Station-The Composition, Growth, and Development of Plant Cell Walls under Microgravity Conditions. Plants 2015, 4, 44–62. [Google Scholar] [CrossRef]
- Robinson, S.; Kuhlemeier, C. Global Compression Reorients Cortical Microtubules in Arabidopsis Hypocotyl Epidermis and Promotes Growth. Curr. Biol. CB 2018, 28, 1794–1802.e2. [Google Scholar] [CrossRef] [Green Version]
- Kume, A.; Kamachi, H.; Onoda, Y.; Hanba, Y.T.; Hiwatashi, Y.; Karahara, I.; Fujita, T. How plants grow under gravity conditions besides 1 g: Perspectives from hypergravity and space experiments that employ bryophytes as a model organism. Plant Mol. Biol. 2021. [Google Scholar] [CrossRef]
- Paul, A.L.; Wheeler, R.M.; Levine, H.G.; Ferl, R.J. Fundamental Plant Biology Enabled by the Space Shuttle. Am. J. Bot. 2013, 100, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Benavides Damm, T.; Walther, I.; Wuest, S.L.; Sekler, J.; Egli, M. Cell cultivation under different gravitational loads using a novel random positioning incubator. Biotechnol. Bioeng. 2014, 111, 1180–1190. [Google Scholar] [CrossRef] [Green Version]
- Correll, M.J.; Pyle, T.P.; Millar, K.D.L.; Sun, Y.J.; Yao, J.; Edelmann, R.E.; Kiss, J.Z. Transcriptome analyses of Arabidopsis thaliana seedlings grown in space: Implications for gravity-responsive genes. Planta 2013, 238, 519–533. [Google Scholar] [CrossRef]
- Kuang, A.; Popova, A.; Xiao, Y.; Musgrave, M.E. Pollination and embryo development in Brassica rapa L. in microgravity. Int. J. Plant Sci. 2000, 161, 203–211. [Google Scholar] [CrossRef]
- Sychev, V.N.; Levinskikh, M.A.; Gostimsky, S.A.; Bingham, G.E.; Podolsky, I.G. Spaceflight effects on consecutive generations of peas grown onboard the Russian segment of the International Space Station. Acta Astronaut. 2007, 60, 426–432. [Google Scholar] [CrossRef]
- Foster, J.S.; Wheeler, R.M.; Pamphile, R. Host-microbe interactions in microgravity: Assessment and implications. Life 2014, 4, 250–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, A.; Lee, S.H.; Cho, H.T. Functional identification of the phosphorylation sites of Arabidopsis PIN-FORMED3 for its subcellular localization and biological role. Plant J. Cell Mol. Biol. 2012, 71, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.R.; Masson, P.H. ARL2, ARG1 and PIN3 define a gravity signal transduction pathway in root statocytes. Plant J. Cell Mol. Biol. 2008, 53, 380–392. [Google Scholar] [CrossRef]
- Su, S.H.; Gibbs, N.M.; Jancewicz, A.L.; Masson, P.H. Molecular Mechanisms of Root Gravitropism. Curr. Biol. CB 2017, 27, R964–R972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Liu, B.; Chen, X.; Zhu, H.; Zou, C.; Men, S. AUX1 acts upstream of PIN2 in regulating root gravitropism. Biochem. Biophys. Res. Commun. 2018, 507, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Scherer, G.F.; Pietrzyk, P. Gravity-dependent differentiation and root coils in Arabidopsis thaliana wild type and phospholipase-A-I knockdown mutant grown on the International Space Station. Plant Biol. 2014, 16 (Suppl. 1), 97–106. [Google Scholar] [CrossRef] [PubMed]
- Harmer, S.L.; Brooks, C.J. Growth-mediated plant movements: Hidden in plain sight. Curr. Opin. Plant Biol. 2018, 41, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Soga, K.; Wakabayashi, K.; Kamisaka, S.; Hoson, T. Stimulation of elongation growth and xyloglucan breakdown in Arabidopsis hypocotyls under microgravity conditions in space. Planta 2002, 215, 1040–1046. [Google Scholar] [CrossRef]
- Manzano, A.I.; van Loon, J.J.; Christianen, P.C.; Gonzalez-Rubio, J.M.; Medina, F.J.; Herranz, R. Gravitational and magnetic field variations synergize to cause subtle variations in the global transcriptional state of Arabidopsis in vitro callus cultures. BMC Genom. 2012, 13, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abas, L.; Benjamins, R.; Malenica, N.; Paciorek, T.; Wisniewska, J.; Moulinier-Anzola, J.C.; Sieberer, T.; Friml, J.; Luschnig, C. Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root gravitropism. Nat. Cell Biol. 2006, 8, 249–256. [Google Scholar] [CrossRef]
- Kordyum, E.L. Plant cell gravisensitivity and adaptation to microgravity. Plant Biol. 2014, 16 (Suppl. 1), 79–90. [Google Scholar] [CrossRef] [PubMed]
- Fasano, J.M.; Swanson, S.J.; Blancaflor, E.B.; Dowd, P.E.; Kao, T.H.; Gilroy, S. Changes in root cap pH are required for the gravity response of the Arabidopsis root. Plant Cell 2001, 13, 907–921. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Z.; Yang, K.Z.; Zou, J.J.; Zhu, L.L.; Xie, Z.D.; Morita, M.T.; Tasaka, M.; Friml, J.; Grotewold, E.; Beeckman, T.; et al. Transcriptional regulation of PIN genes by FOUR LIPS and MYB88 during Arabidopsis root gravitropism. Nat. Commun. 2015, 6, 8822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleine-Vehn, J.; Ding, Z.; Jones, A.R.; Tasaka, M.; Morita, M.T.; Friml, J. Gravity-induced PIN transcytosis for polarization of auxin fluxes in gravity-sensing root cells. Proc. Natl. Acad. Sci. USA 2010, 107, 22344–22349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, K.L.; Strohm, A.K.; Masson, P.H. Gravity sensing and signal transduction in vascular plant primary roots. Am. J. Bot 2013, 100, 126–142. [Google Scholar] [CrossRef]
- Hader, D.P.; Lebert, M. Graviperception and gravitaxis in algae. Adv. Space Res. 2001, 27, 861–870. [Google Scholar] [CrossRef]
- Valbuena, M.A.; Manzano, A.; Vandenbrink, J.P.; Pereda-Loth, V.; Carnero-Diaz, E.; Edelmann, R.E.; Kiss, J.Z.; Herranz, R.; Medina, F.J. The combined effects of real or simulated microgravity and red-light photoactivation on plant root meristematic cells. Planta 2018, 248, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.H.; Liu, M.J.; Xiong, Y.; Sheen, J.; Wu, S.H. TOR and RPS6 transmit light signals to enhance protein translation in deetiolating Arabidopsis seedlings. Proc. Natl. Acad. Sci. USA 2018, 115, 12823–12828. [Google Scholar] [CrossRef] [Green Version]
- Vandenbrink, J.P.; Herranz, R.; Medina, F.J.; Edelmann, R.E.; Kiss, J.Z. A novel blue-light phototropic response is revealed in roots of Arabidopsis thaliana in microgravity. Planta 2016, 244, 1201–1215. [Google Scholar] [CrossRef]
- Kiss, J.Z.; Millar, K.D.; Edelmann, R.E. Phototropism of Arabidopsis thaliana in microgravity and fractional gravity on the International Space Station. Planta 2012, 236, 635–645. [Google Scholar] [CrossRef]
- Manzano, A.; Herranz, R.; den Toom, L.A.; Te Slaa, S.; Borst, G.; Visser, M.; Medina, F.J.; van Loon, J. Novel, Moon and Mars, partial gravity simulation paradigms and their effects on the balance between cell growth and cell proliferation during early plant development. NPJ Microgravity 2018, 4, 9. [Google Scholar] [CrossRef]
- Hoson, T. Plant Growth and Morphogenesis under Different Gravity Conditions: Relevance to Plant Life in Space. Life 2014, 4, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.Q.; Han, F.; Le, J. Higher Plants in Space: Microgravity Perception, Response, and Adaptation. Microgravity Sci. Technol. 2015, 27, 377–386. [Google Scholar] [CrossRef]
- Ferl, R.J.; Paul, A.L. Lunar plant biology—A review of the Apollo era. Astrobiology 2010, 10, 261–274. [Google Scholar] [CrossRef]
- Fisahn, J.; Klingele, E.; Barlow, P. Lunar gravity affects leaf movement of Arabidopsis thaliana in the International Space Station. Planta 2015, 241, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Baque, M.; Verseux, C.; Bottger, U.; Rabbow, E.; de Vera, J.P.; Billi, D. Preservation of Biomarkers from Cyanobacteria Mixed with Mars-Like Regolith Under Simulated Martian Atmosphere and UV Flux. Orig. Life Evol. Biosph. J. Int. Soc. Study Orig. Life 2016, 46, 289–310. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.A.; Palma, C.F.; Marcelis, L.; Kittang Jost, A.I.; van Delden, S.H. Testing New Concepts for Crop Cultivation in Space: Effects of Rooting Volume and Nitrogen Availability. Life 2018, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, L.; Xie, J.; Zheng, H. Differential protein expression profiling of Arabidopsis thaliana callus under microgravity on board the Chinese SZ-8 spacecraft. Planta 2015, 241, 475–488. [Google Scholar] [CrossRef]
- Aerts, J.W.; Roling, W.F.; Elsaesser, A.; Ehrenfreund, P. Biota and biomolecules in extreme environments on Earth: Implications for life detection on Mars. Life 2014, 4, 535–565. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Sumner, D.Y.; Kah, L.C.; Stack, K.; Gupta, S.; Edgar, L.; Rubin, D.; Lewis, K.; Schieber, J.; Mangold, N.; et al. A Habitable Fluvio-Lacustrine Environment at Yellowknife Bay, Gale Crater, Mars. Science 2014, 343. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, A.R.; Hurford, T.A.; Barge, L.M.; Bland, M.T.; Bowman, J.S.; Brinckerhoff, W.; Buratti, B.J.; Cable, M.L.; Castillo-Rogez, J.; Collins, G.C.; et al. The NASA Roadmap to Ocean Worlds. Astrobiology 2019, 19, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, D.; Jian, Y.; Zhang, Y.; Xie, G. Plant Gravitropism and Signal Conversion under a Stress Environment of Altered Gravity. Int. J. Mol. Sci. 2021, 22, 11723. https://doi.org/10.3390/ijms222111723
Qiu D, Jian Y, Zhang Y, Xie G. Plant Gravitropism and Signal Conversion under a Stress Environment of Altered Gravity. International Journal of Molecular Sciences. 2021; 22(21):11723. https://doi.org/10.3390/ijms222111723
Chicago/Turabian StyleQiu, Dan, Yongfei Jian, Yuanxun Zhang, and Gengxin Xie. 2021. "Plant Gravitropism and Signal Conversion under a Stress Environment of Altered Gravity" International Journal of Molecular Sciences 22, no. 21: 11723. https://doi.org/10.3390/ijms222111723
APA StyleQiu, D., Jian, Y., Zhang, Y., & Xie, G. (2021). Plant Gravitropism and Signal Conversion under a Stress Environment of Altered Gravity. International Journal of Molecular Sciences, 22(21), 11723. https://doi.org/10.3390/ijms222111723