1. Introduction
Glucocorticoids (GCs) are steroid hormones which regulate the various metabolic and homeostatic processes essential for life: inflammatory and immune reactions, metabolic control, fertility and reproduction, cognitive function and development. Their role is vital for the physiological function of many organs.
Although endogenous hypercortisolism is a relatively rare condition (the incidence of newly diagnosed cases varies between 0.7–3.0/1 million inhabitants/year [
1,
2]), synthetic glucocorticoids are widely used for their anti-inflammatory and immunomodulatory effects. GCs are essential in the treatment of chronic inflammatory diseases, after organ transplantations and in the field of oncohematology, but the harmful effects accompanying hypercortisolic periods throughout prolonged GC administration represent a serious risk for many pathological conditions.
GC signaling is mediated by the glucocorticoid receptor whose activation ultimately leads to the stimulation of the target gene’s expression [
3,
4]. Diverse signaling mechanisms admit glucocorticoids to control the physiological processes at the level of different organs and tissues, but it is difficult to define the “main” signaling pathways which are responsible for these processes. The most studied pathways related to GC signaling are: the GH/IGF-1 axis, TGFβ-SMAD signaling, PI3/Akt signaling, MAPK signaling, Wnt-signaling, BMP signaling and NF-κB signaling [
5,
6,
7,
8,
9,
10,
11].
Wnt ligands are small glycoproteins which bind to the Frizzled (Fz) receptor. In the absence of a Wnt ligand, a destruction complex composed of Axin and adenomatous polyposis coli is responsible for the constant phosphorylation of β-catenin by glycogen synthase kinase 3β (GSK-3β) and casein kinase type 1. The binding of Wnt to the Fz receptor requires the presence of low-density lipoprotein receptor-related-protein 5 or 6 (LRP 5/6) as coreceptor. Wnt-Fz-LRP 5/6 complex formation leads to the membrane recruitment of the Axin complex and to the consequential suspension of the degrading complex’s operation. Escaping from phosphorylation, the free β-catenin level increases and forms complexes with the DNA-bounded T cell factor/lymphoid enhancer factor (TCF/LEF) to activate Wnt target gene expression. Wnt-pathway agonists are a large family from which the role of the Dickkopf family (Dkks), sclerostin (Sost) and secreted Frizzled-related proteins (sFRPs) were identified as important inhibitors in glucocorticoid excess [
12,
13,
14,
15].
MiRNAs, evolutionary conserved non-coding RNAs regulate various biological processes through post-transcriptional modifications of mRNAs. Their role was demonstrated in numerous physiological and pathological processes in different tissues, including the development, homeostasis and aging processes [
16].
The mediating role of the miRNAs in glucocorticoid action has been extensively studied. miRNAs regulate glucocorticoid production and glucocorticoid availability, the latter being accomplished through the modulation of glucocorticoid receptors’ expression levels [
17]. They also have an important role in cell response to the presence of glucocorticoids [
17]. MiRNAs are implicated in the regulation of immune and inflammatory processes, through mediating glucocorticoids’ tumor-suppressive effects in hematological malignancies (and also, in the regulation of resistance to GC-induced apoptotic processes) and in fine-tuning the responses of GCs during developmental processes and through activities which require continuous remodeling (brain, bones) [
17].
Beyond the physiological processes, miRNAs have been identified as the intermediaries of modified functioning during GC excess.
Hypercortisolism represents a risk factor for cardiovascular events which can be explained, at least partly, by endothelial dysfunctions. In patients with Cushing’s syndrome (CS), miRNA analysis identified the dysregulation of endothelium-related miRNAs: miRNA-17-5p, miRNA-126-3p, and miRNA-126-5p were downregulated, while miRNA-150-5p and miRNA-223-3p were upregulated compared to controls [
18]. Thrombospondin-1, an adhesive glycoprotein associated with platelet aggregation, angiogenesis and tumorigenesis, was downregulated in the pituitary tissue of patients with Cushing’s disease, and this correlated inversely with miR-449c expression [
19].
Using miRNAs, it seems possible to distinguish Cushing’s disease from ectopic ACTH overproduction: miR-16-5p was overexpressed in CD, but downregulated in hypercortisolism from an ectopic source, compared to healthy controls [
20]. Interestingly, in a recent article, miR-122-5p was identified as a possible link between nonphysiological (externally administered) GCs and the effects caused by GC administration. Additionally, the measurement of miR-122-5p expression can be a potential marker to monitor appropriate glucocorticoid activity and to avoid overtreatment [
21].
The aim of our work was to study the involvement of different miRNAs in mediating the effects of hypercortisolism and to reveal the most affected, miRNA-regulated signaling pathways during this process in the GC producing and target tissue/cell types. Our experiment consisted of three models for miRNA profiling. We investigated the changes of the miRNA expression pattern in adrenal tissues obtained from patients with hormonally active Cushing’s syndrome compared with normal adrenal tissues. Then, we examined the changes of miRNA expression in steroid-producing adrenocortical H295R cells and in hormonally inactive (hormone nonproducing) HeLa cells before and after dexamethasone treatment. The HeLa cell line was included as a general model for studying glucocorticoid effects on epithelial cells. It is the most widely used cell line model for investigating human cellular and molecular biology [
22,
23] and it was also reported to be a relevant model for studying glucocorticoid-related effects due to its epithelioid origin and as it expresses glucocorticoid receptor [
24,
25,
26,
27,
28,
29]. Following miRNA-profiling in these experiments, pathway analysis and literature data mining were performed to investigate the relevant miRNA-pathway interactions related to glucocorticoid excess. Furthermore, we analyzed the transcriptome data of GC producing and target organs/cell types (bone tissue originated from Cushing patients and primary bone, synovial fibroblast, subcutaneous and omental adipose tissue, skin and brain cells) upon GC effect. We performed tissue-specific target prediction and gene set enrichment analysis in order to investigate the changes in Wnt signaling.
3. Discussion
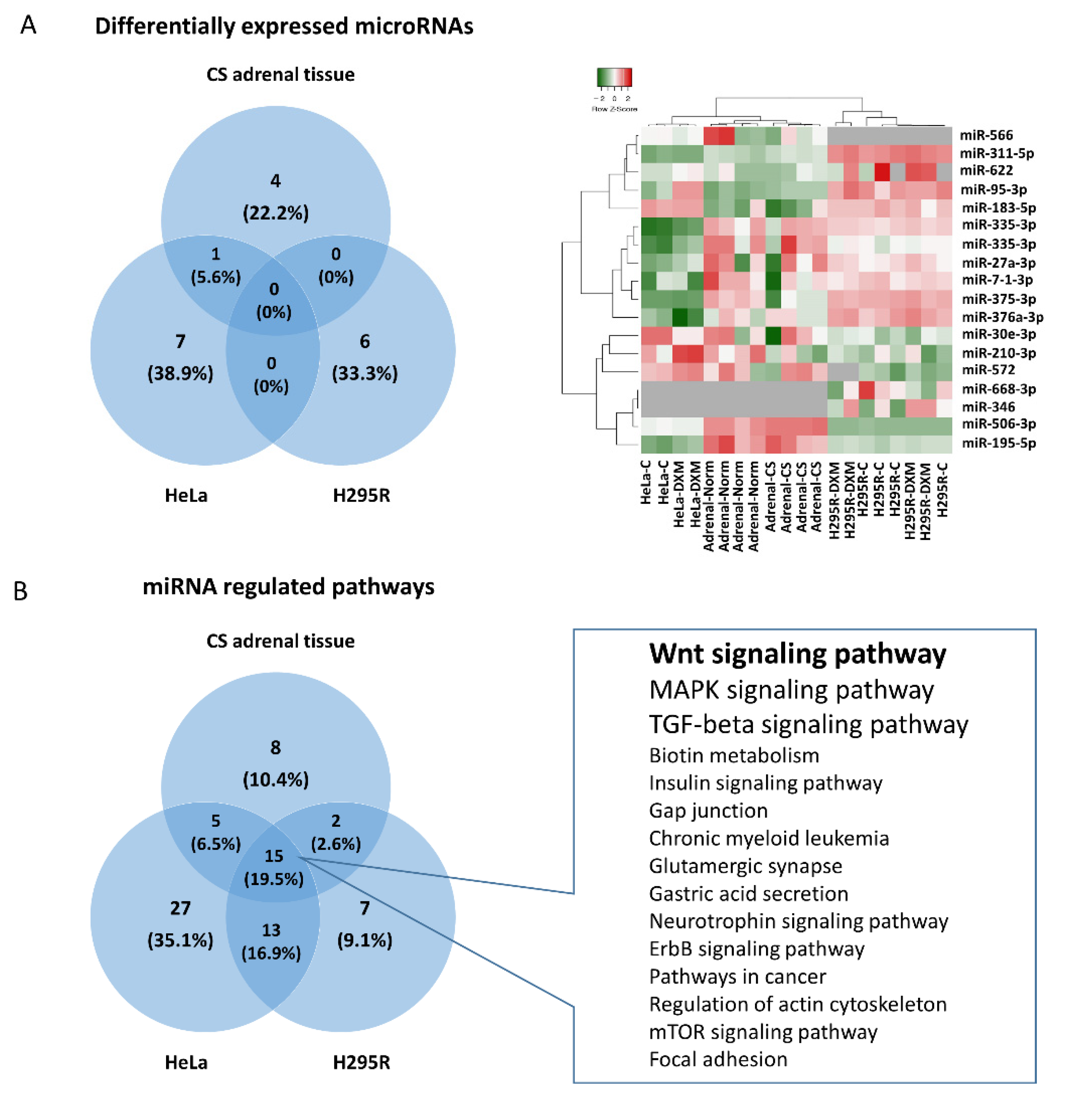
The miRNA expression pattern is characteristic to each cell type; therefore, some miRNAs can be used as potential biomarkers for different conditions as well. GCs have diverse effects on different tissues involving various molecules including miRNAs. Our in vitro data showed that GC excess had significant effects on miRNA expression profiles on all tested cell types. In this context, commonly regulated miRNAs in HeLa and GC-producing/target tissues can be considered important mediators of GC effect, as HeLa cells represent a valid epithelioid-type target model and several GC-related processes occur in these cell types. Studying the biological role of significantly altered miRNAs upon GC treatment in three models, different miRNAs had a convergent effect on the same signaling pathways. Of these pathways, Wnt signaling was identified as the most relevant.
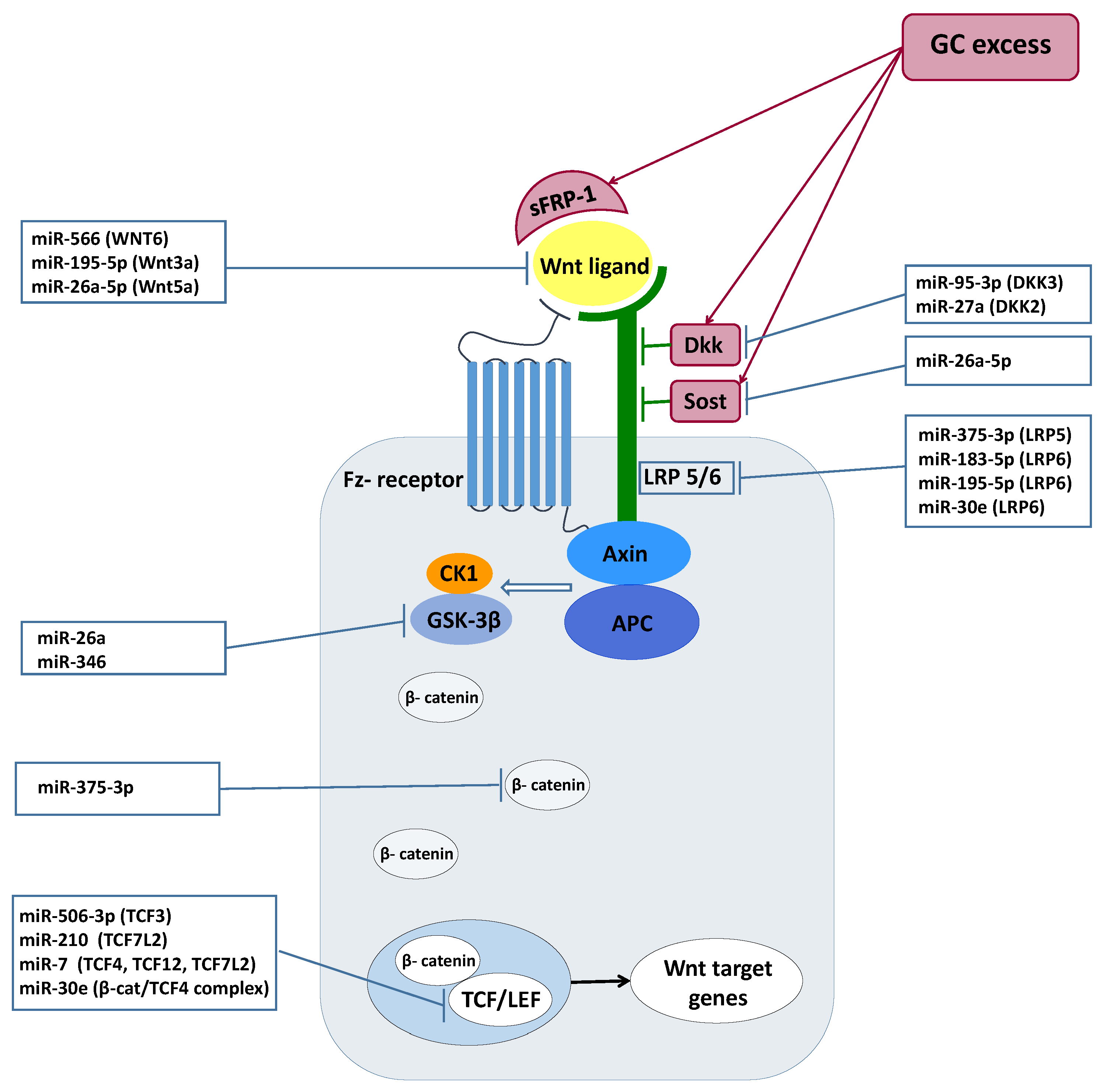
The regulation of Wnt signaling through miRNAs has been studied and confirmed in several different tissues and cell types. Relying on literature data, we summarized the effect of the differentially expressed miRNAs identified in our experiments on the operation of the Wnt-signaling pathway. (
Figure 2)
GC effect on theGC-producing adrenal gland. Interestingly, the tissue-specific target prediction indicated that even the cortisol-producing adrenal GCs have a feedback effect on miRNA expression that, as a consequence, influences Wnt signaling, among others. As expected, GC had a lower impact on the whole transcriptome in GC-producing cells and tissues compared to GC target tissues (4–8% of all genes changed vs. ~10–20%, respectively). It was described that miRNAs could play a role in regulating the operation of glucocorticoid receptors (GR) and consequently circadian rhythm [
61]. The adrenal gland has its own peripheral circadian clock which is tightly linked to steroidogenesis by the steroidogenic acute regulatory protein [
62]. These may give a potential explanation to the GC feedback effect in the adrenal gland.
Additionally, global transcriptome data indicated that in the adrenal gland the GCs regulated the Wnt pathway. Indeed, using both miRNA and gene expression data we performed tissue specific target prediction that indicated that in adrenal tissue miR-375 and miR-506 regulated SFRP4, APC, NFAT5 and CTBP2 expression among Wnt signaling members upon GC effect.
In the adrenal tissues obtained from hypercortisolic patients, the Wnt-signaling pathway was ranked backward compared to in vitro cell line experiments. Here, Wnt-signaling is preceded by
Biotin metabolism,
Long-term depression,
Insulin signaling pathway and
Adipocytokine signaling pathway. Not surprisingly, these pathways define some of the characteristic symptoms of Cushing’s disease including obesity, insulin resistance, depression, fatigue, irritability, insomnia, hair loss, dry skin [
63,
64]. This draws the attention to evaluate the results obtained on cell lines carefully as they miss the complex environment which is available in tissue experiments and in in vivo environments.
GC effect on the GC-target cells/tissues. In order to investigate the potential involvement of Wnt signaling in the GC effect on GC target tissues, we re-analyzed the data available in data repositories collected from bone tissue originating from Cushing patients and primary bone, synovial fibroblast, subcutaneous and omental adipose tissue, skin and brain cells upon GC effect. We found that the ratio of transcriptome regulated by GC depended on tissue type. Our results were in line with previous findings that in skin a relatively smaller gene set (6.3%) while in adipose tissue, brain and peripheral blood mononuclear cells a larger gene set (10–20%) were regulated by GC, [
21,
58,
65,
66]. Interestingly, in the liver GC induced vast transcriptional responses, with more than 30% of genes being regulated [
59]. As we focused on Wnt signaling we investigated its involvement in whole transcriptome changes by three approaches (i) gene set enrichment analysis of global gene expression profile, (ii) gene set enrichment analysis of commonly changed gene profile at least in six different studies and (iii) analysis of individual Wnt pathways. In all approaches Wnt signaling was significantly altered upon GC effect and additionally we identified several miRNAs implicated in this pathway.
miRNAs influencing Wnt signaling. The identified GC-mediated miRNAs were demonstrated to influence Wnt signaling at different levels. Regarding the
Wnt ligands, miR-195 and miR-26a-5p, suppressed the Wnt/β-catenin pathway among others through repressing
WNT3a, WNT5a, and
WNT7a in osteogenic differentiation of mesenchymal stem cells and periodontal ligament cells [
38,
39,
41].
WNT6 was also proved to be a target of miR-566 in human breast cancer [
31].
Wnt co-receptors were also described as being controlled by GC-regulated miRNAs. LDL receptor-related proteins 5/6 (
LRP5 and 6) as co-receptors are indispensable for the activation of the canonical pathway. It was shown that miR-375-3p decreased the levels of
LRP5 and β-catenin by directly binding to their 3′UTR, therefore negatively regulating Wnt signaling [
30]. Similarly, miR-183, by targeting
LRP6, inhibited Wnt/β-catenin signaling pathway and consequently promoted adipogenesis [
36]. Among
Wnt agonists,
DKK3 was demonstrated to be regulated by miR-183 in prostate cancer [
37]. miR-95-3p promoted cell proliferation, migration and invasion in PCa by targeting also
DKK3 and activating the Wnt/β-catenin pathway [
32]. Additionally,
DKK2, which is another direct inhibitor of Wnt binding to
LRP5/6, was negatively regulated by miR-27a in in vitro miRNA overexpression and inhibition experiments [
48].
Wnt pathway mediators are also regulated by GC-affected miRNAs. While the Wnt pathway effector
TCF7L2 was regulated by miR-7 leading to inhibition of Wnt/β-catenin signaling in neuronal cells [
47],
TCF3, another Wnt mediator, was repressed by miR-506-3p, resulting in neural stem cell proliferation and differentiation [
33].
The role of GC mediated miRNAs on biological processes. Regarding the potential function of miRNAs altered upon GC effect, current literature data suggested that Wnt-signaling can be associated with cell proliferation and differentiation, tumorigenesis and bone homeostasis.
Cell proliferation and differentiation. MiR-506-3p upregulated in Cushing adenomas compared to normal tissues. In neural stem cells, the overexpression of this miRNA increased the cells’ differentiation and reduced their proliferation through influencing Wnt/β-catenin pathway by targeting
TCF3 [
33]. In dermal papilla cells, miR-195-5p, by targeting LRP6 protein expression, influenced hair follicle inductivity [
38]. The overexpression of miR-195-5p significantly inhibited osteogenic differentiation of periodontal ligament cells under mechanical loading together with regulating directly the WNT family member 3A (
WNT3A) [
39]. Upregulated miR-195-5p following GC treatment could suggest that this miRNA contributed to the bone damage and hair loss frequently observed in Cushing’s syndrome.
Tumorigenesis. In Cushing adenomas, as in a high glucocorticoid environment, we detected decreased miR-566 level. In breast cancer patients, the low expression of this miRNA predisposed to larger tumor size, advanced tumor grade and higher incidence of lymphatic metastasis [
31]. MiR-566 represents an important link in this process by targeting
WNT6 and preventing malignant progression in breast cancer patients [
31]. miR-95-3p, that was described upregulated in prostate carcinoma tissues, promoted cell proliferation, migration and invasion of cancerous cells through repressing
DKK3 prostate [
32]. miR-183-5p expression was reported significantly upregulated in colorectal cancer tissues compared to normal tissues [
67,
68]. Its role in Wnt regulation was demonstrated by pathway analysis and functional miR-inhibition experiments that also led to the downregulation of the Wnt pathway downstream genes [
67,
68].
Bone homeostasis. Bones and bone-forming mechanisms are severely damaged by GC excess and a number of miRNAs have been identified that play important roles during these processes. Indeed, we presented the hypothesis that the expression profile of both human bone tissue biopsies from CS patients and patient-derived primary osteoblast cell lines treated by dexamethasone indicated that the Wnt pathway was significantly influenced by miRNAs. We also suggested that several GC-regulated miRNAs were demonstrated to be targeting Wnt signaling, including miR-375-3p, miR-210-3p and miR-26a-5p. MiR-375-3p was described to negatively modulate osteogenesis by inducing apoptosis in the mouse osteoblastic MC3T3-E1 cell line through targeting
LRP5 and β-catenin [
30]. MiR-26a-5p also played a negative role by hindering the osteogenic differentiation of adipose-derived mesenchymal stem cells. This effect is accomplished directly, through the inhibition of
WNT5a expression [
41]. Overexpression of miR-210-3p promoted osteogenic differentiation and inhibited adipogenic differentiation in bone marrow-derived mesenchymal stem cells (BMSCs). In cells overexpressing miR-210 regulatory factors of the Wnt signaling pathway, such as
LRP5,
GSK-3β, β-catenin and
TCF4 were significantly increased [
34]. The osteogenic effect of this miRNA was also demonstrated by downregulating the Wnt-pathway inhibitor sclerostin and by enhancing the migration capability of BMSCs [
35]. In line with the in vitro results mentioned above, miR-210-3p was found to be downregulated in bone marrow samples of women suffering from postmenopausal osteoporosis [
69]. In sphenoid bone samples of patients with active Cushing’s disease, the expression of miR-26a-5p was upregulated compared to patients with nonfunctioning pituitary adenoma [
70].
Our results corroborate previous general knowledge about the function of miRNAs. Namely, that (i) miRNAs, similarly to mRNAs, have a tissue-specific expressional pattern [
71], (ii) miRNAs exert a fine-tuning effect [
72], and (iii) they work in network [
73]. These three properties of miRNAs are also manifest in the mediation of the glucocorticoid effect. In different tissues different miRNA expressional patterns can be observed of the glucocorticoid effect, however, these differentially expressed miRNAs regulate similar biological processes in the different tissues, in particular, Wnt signaling.
4. Materials and Methods
4.1. Patients
Adrenal adenoma specimens from four patients with active Cushing’s syndrome were obtained during surgical interventions at Semmelweis University Department of Transplantation and Surgery. Normal adrenal samples from another four patients adjacent to hormonally inactive adrenal adenomas served as controls. The study was conducted in accordance with the Declaration of Helsinki and has been approved by the Scientific and Research Committee of the Medical Research Council of Hungary (ETT-TUKEB 4457/2012/EKU, approval date: 2 February 2012). All subjects gave written informed consent in accordance with the Declaration of Helsinki.
4.2. In Vitro Cell Culture Experiments
HeLa human epithelioid cervix cells were cultured in MEM (Gibco, 31095029, Thermofisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (Gibco 10270106, Thermofisher Scientific, Waltham, MA, USA), 1% sodium-pyruvate (Gibco, 11360070, Thermofisher Scientific, Waltham, MA, USA), and 1% antibiotic-antimycotic solution (Sigma-Aldrich, Merck, Kenilworth, NJ, USA). H295R cells were grown in Dulbecco’s Modified Eagle Medium Nutrient Mixture F-12 (Gibco: 10565018, Thermofisher Scientific, Waltham, MA, USA) supplemented with 2.5% Nuserum (Corning 355100, Corning, NY, USA), 1% Insulin-Transferrin-Selenium solution (41400-045 Gibco, Thermofisher Scientific, Waltham, MA, USA) and 1% antibiotic-antimycotic solution (Sigma-Aldrich, Merck, Kenilworth, NJ, USA). Cells were cultivated in a humidified incubator infused with 5% CO2 at 37 °C.
Before treatment, cells were grown in their complete media using hormone-free fetal bovine serum for 48 h. Hormone-free FBS were prepared by incubating and mixing 0.1 g dextran-coated active charcoal (C6241, Sigma-Aldrich, Merck, Kenilworth, NJ, USA) per 6 mL FBS for 24 h at 4 °C. After 24 h, mixtures were centrifuged 300× g for 10 min until charcoal settled and supernatant was filtered through 0.22 μm filter.
A measure of 100 nM dexamethasone (D4902, Sigma-Aldrich, Merck, Kenilworth, NJ, USA) or dimethyl sulfoxide (DMSO, 276855, Sigma-Aldrich, Merck, Kenilworth, NJ, USA) as vehicle treatment was applied on both cell lines for 24 h.
4.3. Gene Expression Profiling and Gene Expression Re-Analysis
Investigating transcriptome changes in GC-producing (adrenal tissue and cell line) and target organs (bone, synovial fibroblast, subcutaneous and omental adipose tissue, skin and brain) high-throughput mRNA profiling data (gene expression microarray, RNA sequencing and RT-PCR array [
51,
52,
53,
54,
55,
56,
57,
58,
74]) were downloaded from NCBI Gene Expression Omnibus (
Table 6). Altogether, data of 11 studies (82 control samples and 57 GC effected samples) were included. In order to reduce bias originating from usage of different platforms, different analysis type and cut-offs, we re-analyzed data using the same bioinformatical process, GEO2R algorithm with default settings and Benjamini−Hochberg (False discovery rate) adjustment [
75].
4.4. RNA Extraction
Tumor tissue specimens were immediately frozen in liquid nitrogen after the surgery and stored at −80 °C until further use. From both tissues and cell cultures total RNA was extracted with miRNeasy Mini Kit (Qiagen Inc., Chatsworth, Los Angeles, CA, USA). RNA integrity and concentration were measured using Agilent Bioanalyzer 2100 System (AgilentTech Inc., Santa Clara, CA, USA).
4.5. miRNA Profiling in Adrenocortical Tissues Using TLDA Cards
All procedures were performed following the manufacturer’s instructions and as previously described [
76]. Briefly, 900 ng total RNA per sample was reverse transcribed using Megaplex RT primer Pool A and B using TaqMan MicroRNA Reverse Transcription Kit (P/N: 4366596). MiRNA expression profiles were investigated using TaqMan Low Density Array (TLDA) Human Micro RNA Panel v.1 (Applied Biosystems, Foster City, CA, USA) on 7900HT Fast Real-Time PCR System (Applied Biosystems).
RT–qPCR data analysis was performed using RQ Manager 1.2 (Applied Biosystems). Real-Time StatMiner software (Integromics, Granada, Spain) were used to assess best endogenous control using Normfinder algorithm. (Normfinder calculates the overall stability for all candidate housekeeping genes tested on a sample set indifferent from its composition. It generates an overall stability rank from distinct intragroup and intergroup measures of variability). RNU48 for adrenal tissues and HeLa cells, the geometric mean of MammU6-RNU44-RNU6B for H295R were used as endogenous controls. Expression level was calculated by the ddCt method, and fold changes were obtained using the formula 2−ddCt.
4.6. Gene Set Enrichment and Pathway Analysis
Differentially expressed miRNAs were uploaded into the DIANA-mirPath v.2.0 tool. In silico target predictions were performed by microT-CDS algorithm and followed by enrichment analysis of multiple miRNA target genes comparing each set of miRNA targets to all known KEGG pathways [
77].
Differentially expressed gene functions were analyzed using ToppGene Suite. Gene set enrichment analyses were performed for Gene Ontology categories (Biological Processes, Molecular Function and Cell Component) and KEGG Pathways gene sets. Results were considered to be significant with p < 0.05.
4.7. Statistical Analysis
Differences between Cushing vs. normal and dexamethasone treatment vs. DMSO treated cells were evaluated using unpaired T-test or Mann–Whitney U test depending on data distribution determined by Shapiro–Wilks normality test. Statistical analysis was performed using Integromics RealTime StatMiner and Statistica 13.4.0.14 (TIBCO Software Inc., Palo Alto, CA, USA).
Hierarchical cluster analysis was performed by UPGMA clustering method using Euclidean distance for similarity measuring. A value of p < 0.05 was considered to be significant.






{kind=link}
{kind=link}