
Distinct Exosomal miRNA Profiles from BALF and Lung Tissue of COPD and IPF Patients

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
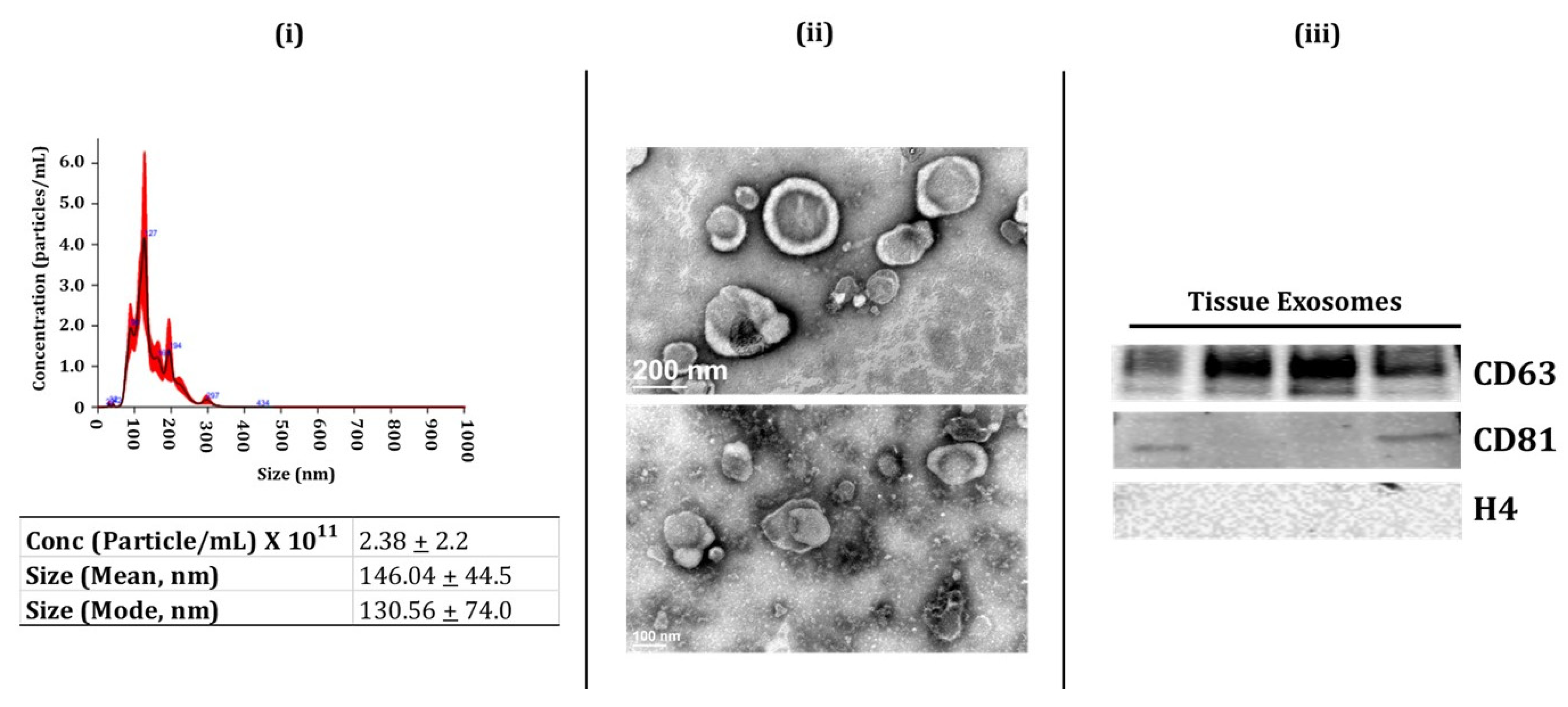
2.1. Characterization of BALF and Lung-Tissue-Derived Exosomes
2.2. Batch Variations in the Exosome-Derived miRNA Expression Profiles amongst the Various Study Groups
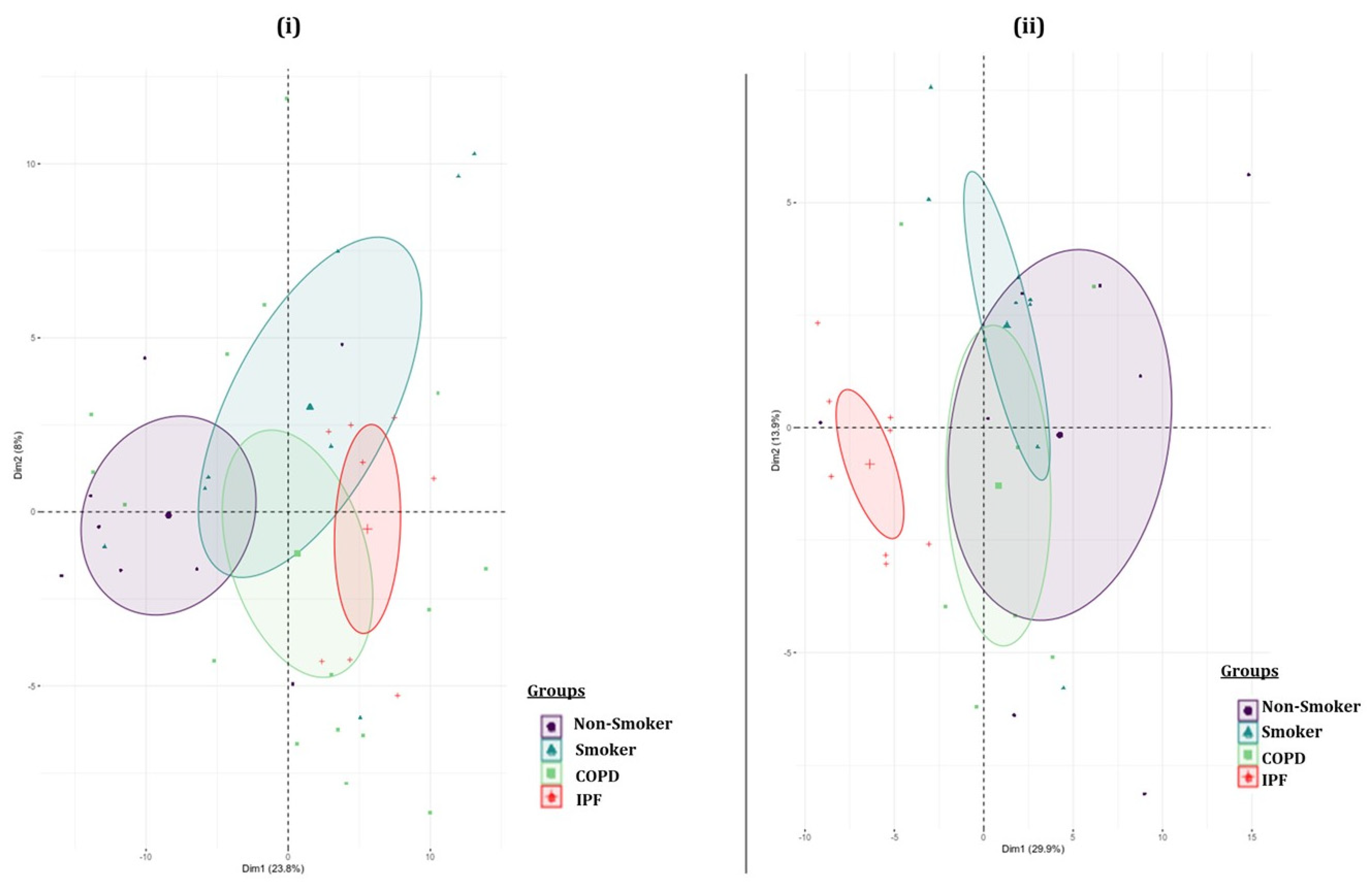
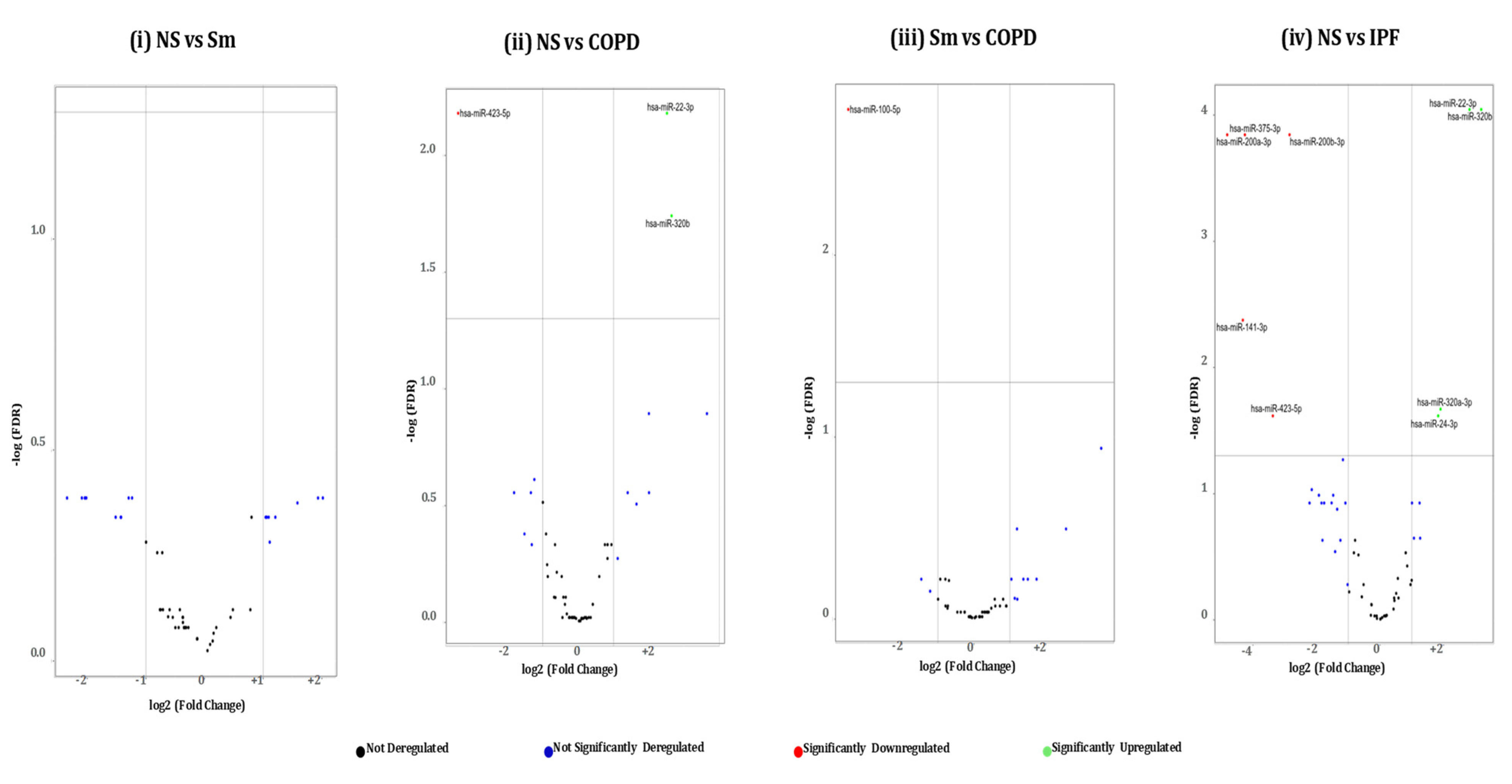
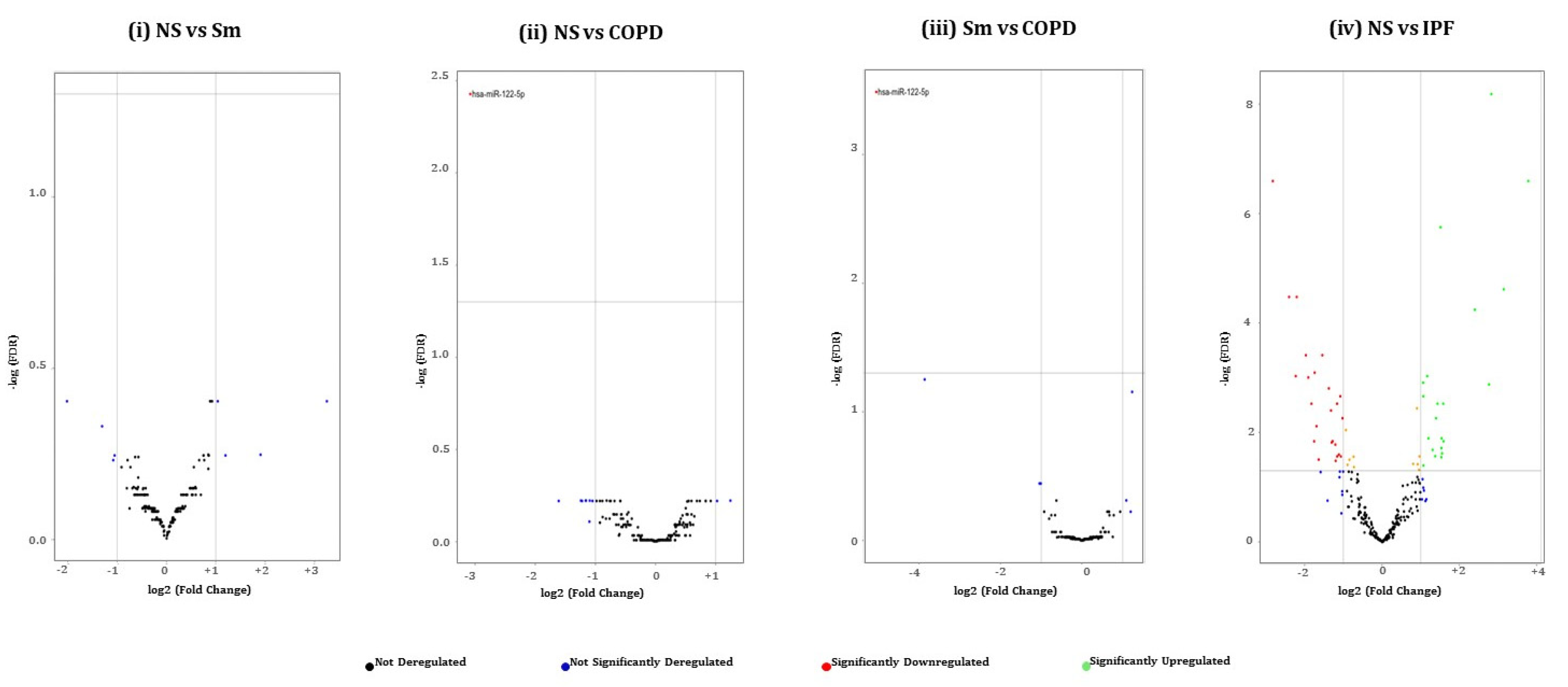
2.3. Pairwise Comparison of BALF- and Lung-Tissue-Derived Exosomal miRNA Expression Profiles
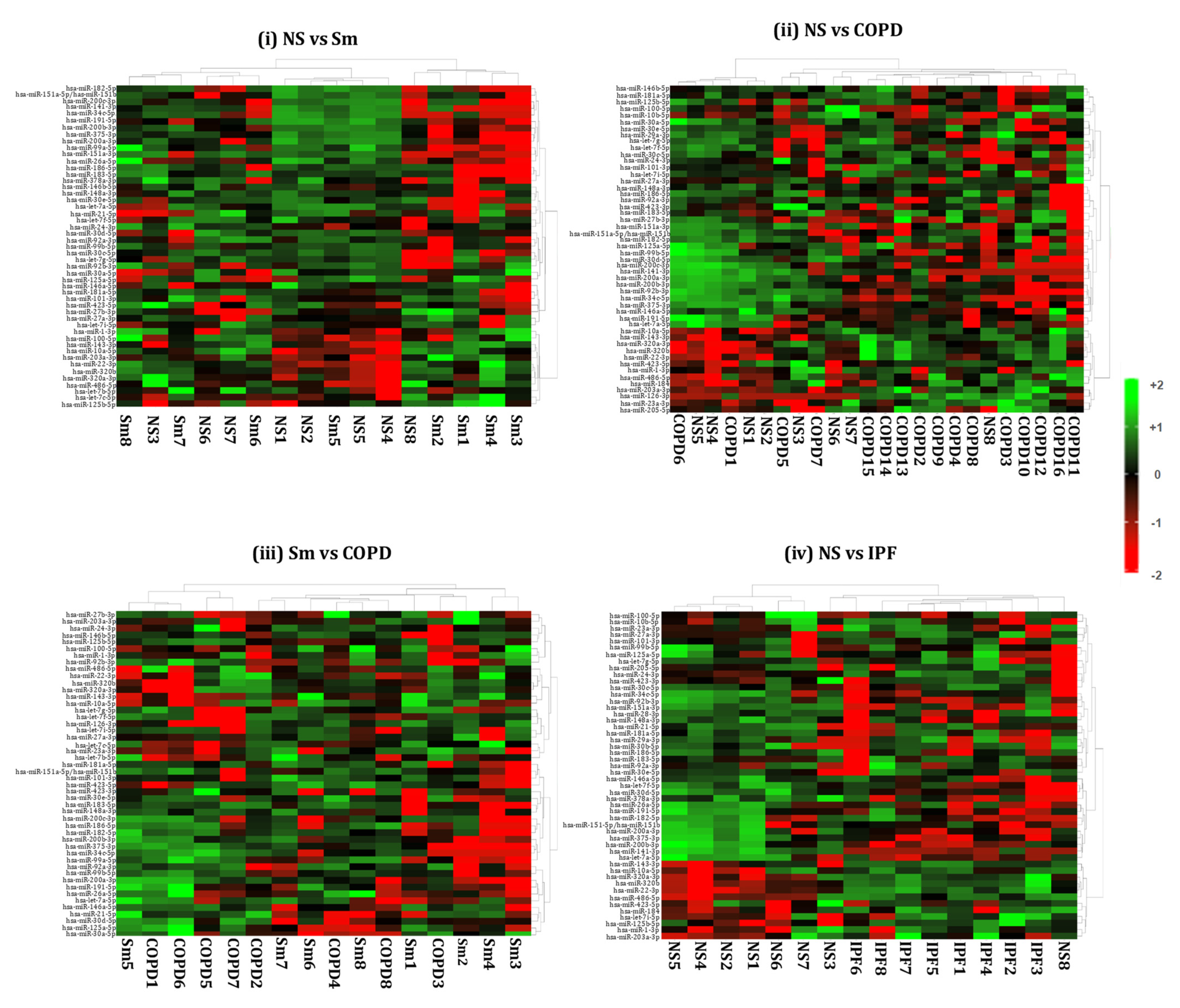
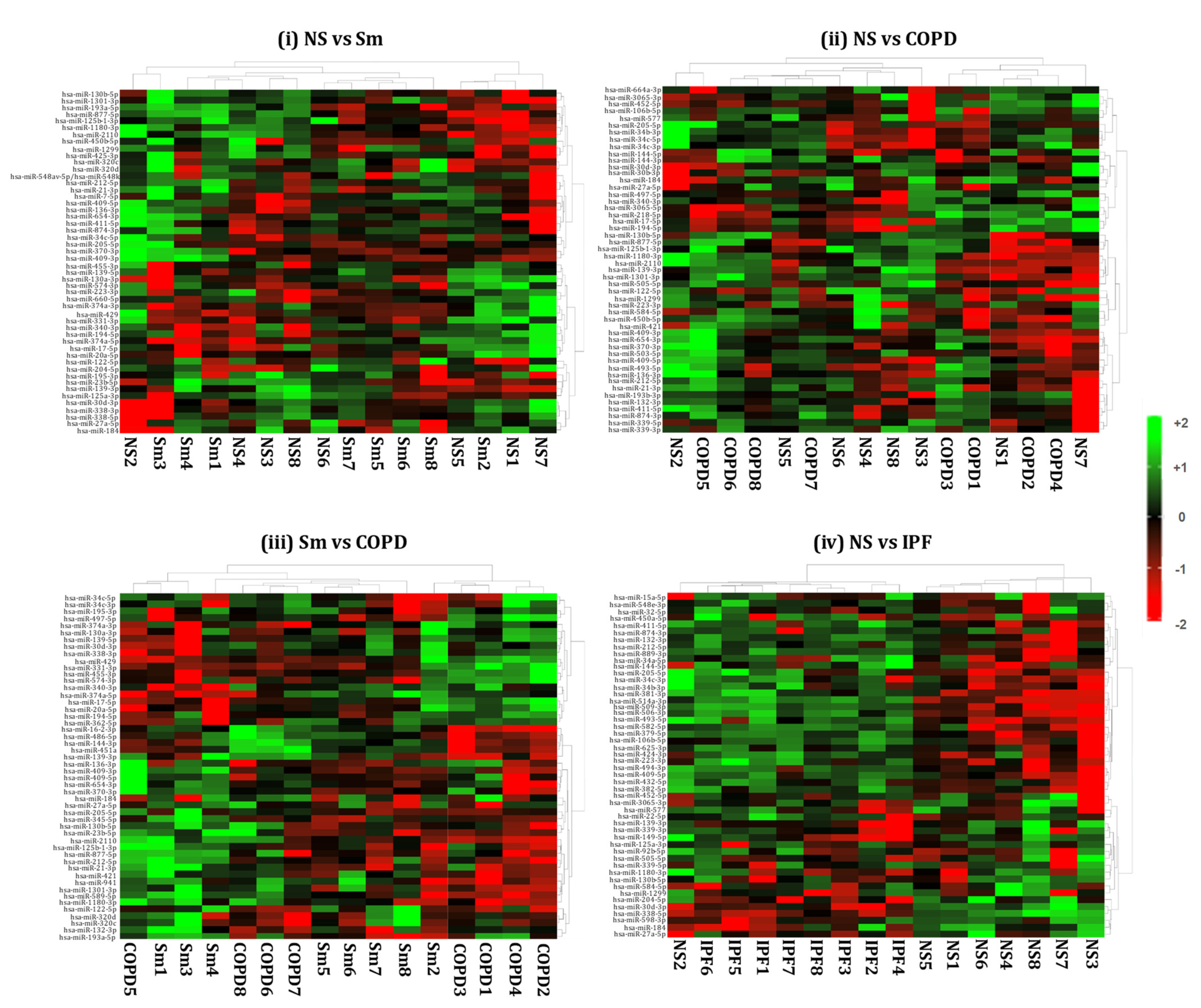
2.4. Hierarchical Clustering Identified Differentially Expressed miRNAs in the BALF or Lung-Tissue-Derived Exosomes from Non-Smokers, Smokers, and Patients of COPD and IPF
2.5. GO Enrichment and KEGG Analyses of Differentially Expressed miRNAs from BALF- and Lung-Derived Exosomes in COPD and IPF Patients
3. Discussion
4. Materials and Methods
4.1. Ethics/Approval
4.2. Study Population and Sample Collection
4.3. BALF Exosome Isolation
4.4. Lung Tissue Exosome Isolation
4.5. Exosome Characterization
4.6. Exosomal RNA Extraction
4.7. Library Preparation
4.8. Next-Generation Sequencing and Data Analysis
4.9. Gene Ontology and KEGG Analyses
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COPD | Chronic obstructive pulmonary disease |
| IPF | Idiopathic pulmonary disease |
| TEM | Transmission electron microscopy |
| NGS | Next-generation sequencing |
| miRNA | Micro RNA |
| BALF | Bronchoalveolar lavage fluid |
| EMT | Epithelial–mesenchymal transition |
| ISEV | International Society for Extracellular Vesicles |
References
- Sousa, C.; Rodrigues, M.; Carvalho, A.; Viamonte, B.; Cunha, R.; Guimarães, S.; de Moura, C.S.; Morais, A.; Pereira, J.M. Diffuse smoking-related lung diseases: Insights from a radiologic-pathologic correlation. Insights Imaging 2019, 10, 73. [Google Scholar] [CrossRef]
- Ryu, J.; Colby, T.; Hartman, T.; Vassallo, R. Smoking-related interstitial lung diseases: A concise review. Eur. Respir. J. 2001, 17, 122–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilosi, M.; Poletti, V.; Rossi, A. The pathogenesis of COPD and IPF: Distinct horns of the same devil? Respir. Res. 2012, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Devine, J.F. Chronic Obstructive Pulmonary Disease: An Overview. Am. Health Drug Benefits 2008, 1, 34–42. [Google Scholar]
- Glassberg, M.K. Overview of idiopathic pulmonary fibrosis, evidence-based guidelines, and recent developments in the treatment landscape. Am. J. Manag. Care 2019, 25, S195–S203. [Google Scholar]
- O’Farrell, H.E.; Yang, I.A. Extracellular vesicles in chronic obstructive pulmonary disease (COPD). J. Thorac. Dis. 2019, 11, S2141–S2154. [Google Scholar] [CrossRef]
- Wang, N.; Wang, Q.; Du, T.; Gabriel, A.N.A.; Wang, X.; Sun, L.; Li, X.; Xu, K.; Jiang, X.; Zhang, Y. The Potential Roles of Exosomes in Chronic Obstructive Pulmonary Disease. Front. Med. 2021, 7, 618506. [Google Scholar] [CrossRef]
- Minnis, P.; Kane, R.; Anglin, R.; Walsh, S.; Worrel, J.; Khan, F.; Lumsden, R.V.; Whitty, S.; Keane, M.P. Serum exosomes from IPF patients display a fibrotic miRNA profile that correlates to clinical measures of disease severity. Eur. Respir. J. 2015, 46, PA3845. [Google Scholar] [CrossRef]
- Njock, M.-S.; Guiot, J.; A Henket, M.; Nivelles, O.; Thiry, M.; Dequiedt, F.; Corhay, J.-L.; E Louis, R.; Struman, I. Sputum exosomes: Promising biomarkers for idiopathic pulmonary fibrosis. Thorax 2018, 74, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Mohan, A.; Agarwal, S.; Clauss, M.; Britt, N.S.; Dhillon, N.K. Extracellular vesicles: Novel communicators in lung diseases. Respir. Res. 2020, 21, 175. [Google Scholar] [CrossRef]
- Nederveen, J.P.; Warnier, G.; Di Carlo, A.; Nilsson, M.I.; Tarnopolsky, M.A. Extracellular Vesicles and Exosomes: Insights From Exercise Science. Front. Physiol. 2021, 11, 1757. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundar, I.K.; Li, D.; Rahman, I. Small RNA-sequence analysis of plasma-derived extracellular vesicle miRNAs in smokers and patients with chronic obstructive pulmonary disease as circulating biomarkers. J. Extracell. Vesicles 2019, 8, 1684816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundar, I.K.; Li, D.; Rahman, I. Proteomic Analysis of Plasma-Derived Extracellular Vesicles in Smokers and Patients with Chronic Obstructive Pulmonary Disease. ACS Omega 2019, 4, 10649–10661. [Google Scholar] [CrossRef]
- Sundar, I.K.; Yao, H.; Rahman, I. Oxidative Stress and Chromatin Remodeling in Chronic Obstructive Pulmonary Disease and Smoking-Related Diseases. Antioxid. Redox Signal. 2013, 18, 1956–1971. [Google Scholar] [CrossRef] [PubMed]
- Lerner, C.A.; Sundar, I.K.; Rahman, I. Mitochondrial redox system, dynamics, and dysfunction in lung inflammaging and COPD. Int. J. Biochem. Cell Biol. 2016, 81, 294–306. [Google Scholar] [CrossRef] [Green Version]
- Salimian, J.; Mirzaei, H.; Moridikia, A.; Harchegani, A.B.; Sahebkar, A.; Salehi, H. Chronic obstructive pulmonary disease: MicroRNAs and exosomes as new diagnostic and therapeutic biomarkers. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2018, 23, 27. [Google Scholar] [CrossRef]
- Francis, S.M.S.; Davidson, M.R.; E Tan, M.; Wright, C.M.; Clarke, B.E.; E Duhig, E.; Bowman, R.V.; Hayward, N.; Fong, K.M.; A Yang, I. MicroRNA-34c is associated with emphysema severity and modulates SERPINE1 expression. BMC Genom. 2014, 15, 88. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Kobayashi, S.; Fujino, N.; Suzuki, T.; Ota, C.; He, M.; Yamada, M.; Suzuki, S.; Yanai, M.; Kurosawa, S.; et al. Increased circulating endothelial microparticles in COPD patients: A potential biomarker for COPD exacerbation susceptibility. Thorax 2012, 67, 1067–1074. [Google Scholar] [CrossRef] [Green Version]
- Porro, C.; Lacedonia, D.; Carpagnano, G.E.; Trotta, T.; Palladino, G.P.; Panaro, M.A.; Zoppo, L.D.; Barbaro, M.P.F. Microparticles in sputum of COPD patients: A potential biomarker of the disease? Int. J. Chronic Obstr. Pulm. Dis. 2016, 11, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.B.; Armitage, J.; Teo, T.-H.; Ong, N.E.; Shin, H.; Moodley, Y.P. Elevated levels of circulating exosome in COPD patients are associated with systemic inflammation. Respir. Med. 2017, 132, 261–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltom, S.; Dale, N.; Raemdonck, K.; Stevenson, C.S.; Snelgrove, R.J.; Sacitharan, P.K.; Recchi, C.; Wavre-Shapton, S.; McAuley, D.; O’Kane, C.; et al. Respiratory Infections Cause the Release of Extracellular Vesicles: Implications in Exacerbation of Asthma/COPD. PLoS ONE 2014, 9, e101087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, C.; Gudi, K.; Krause, A.; Sackrowitz, R.; Harvey, B.-G.; Strulovici-Barel, Y.; Mezey, J.G.; Crystal, R.G. Circulating Endothelial Microparticles as a Measure of Early Lung Destruction in Cigarette Smokers. Am. J. Respir. Crit. Care Med. 2011, 184, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Wang, L.; Wu, Y.; Ou, Y.; Lu, H.; Yao, X. A novel diagnostic signature based on three circulating exosomal mircoRNAs for chronic obstructive pulmonary disease. Exp. Ther. Med. 2021, 22, 1–12. [Google Scholar] [CrossRef]
- Guiot, J.; Njock, M.-S.; Henket, M.; Nivelles, O.; Louis, R.; Struman, I. Sputum exosomal microRNAs in IPF. Eur. Respir. J. 2018, 52, OA2119. [Google Scholar] [CrossRef]
- Liu, B.; Li, R.; Zhang, J.; Meng, C.; Zhang, J.; Song, X.; Lv, C. MicroRNA-708-3p as a potential therapeutic target via the ADAM17-GATA/STAT3 axis in idiopathic pulmonary fibrosis. Exp. Mol. Med. 2018, 50, e465. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-W.R.; Leung, J.M.; Sin, D.D. A Systematic Review of Diagnostic Biomarkers of COPD Exacerbation. PLoS ONE 2016, 11, e0158843. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.G.; Vaughan, A.; Dent, A.G.; O’Hare, P.; Goh, F.; Bowman, R.V.; Fong, K.; Yang, I.A. Biomarkers of progression of chronic obstructive pulmonary disease (COPD). J. Thorac. Dis. 2014, 6, 1532–1547. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.-Y.; Papoutsakis, E.T. Extracellular vesicles: Exosomes, microparticles, their parts, and their targets to enable their biomanufacturing and clinical applications. Curr. Opin. Biotechnol. 2019, 60, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Holtzman, J.; Lee, H. Emerging role of extracellular vesicles in the respiratory system. Exp. Mol. Med. 2020, 52, 887–895. [Google Scholar] [CrossRef]
- Santos, P.; Almeida, F. Role of Exosomal miRNAs and the Tumor Microenvironment in Drug Resistance. Cells 2020, 9, 1450. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Shi, K.; Yang, S.; Liu, J.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Yuan, W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol. Cancer 2018, 17, 1–19. [Google Scholar] [CrossRef]
- Ingenito, F.; Roscigno, G.; Affinito, A.; Nuzzo, S.; Scognamiglio, I.; Quintavalle, C.; Condorelli, G. The Role of Exo-miRNAs in Cancer: A Focus on Therapeutic and Diagnostic Applications. Int. J. Mol. Sci. 2019, 20, 4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Araya, J.; Ito, S.; Kobayashi, K.; Kosaka, N.; Yoshioka, Y.; Kadota, T.; Hara, H.; Kuwano, K.; Ochiya, T. Suppression of autophagy by extracellular vesicles promotes myofibroblast differentiation in COPD pathogenesis. J. Extracell. Vesicles 2015, 4, 28388. [Google Scholar] [CrossRef]
- Bartel, S.; Deshane, J.; Wilkinson, T.; Gabrielsson, S. Extracellular Vesicles as Mediators of Cellular Cross Talk in the Lung Microenvironment. Front. Med. 2020, 7, 326. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Q.; Yu, Q.; Xiao, J.; Zhao, H. Cigarette smoke extract-treated airway epithelial cells-derived exosomes promote M1 macrophage polarization in chronic obstructive pulmonary disease. Int. Immunopharmacol. 2021, 96, 107700. [Google Scholar] [CrossRef]
- Genschmer, K.R.; Russell, D.W.; Lal, C.; Szul, T.; Bratcher, P.E.; Noerager, B.D.; Roda, M.A.; Xu, X.; Rezonzew, G.; Viera, L.; et al. Activated PMN Exosomes: Pathogenic Entities Causing Matrix Destruction and Disease in the Lung. Cell 2019, 176, 113–126.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Kosaka, N.; Araya, J.; Kuwano, K.; Ochiya, T. Extracellular vesicles in lung microenvironment and pathogenesis. Trends Mol. Med. 2015, 21, 533–542. [Google Scholar] [CrossRef]
- Xu, H.; Ji, L.; Yu, C.; Chen, Q.; Ge, Q.; Lu, Y. MiR-423-5P Regulates Cells Apoptosis and Extracellular Matrix Degradation via Nucleotide-Binding, Leucine-Rich Repeat Containing X1 (NLRX1) in Interleukin 1 beta (IL-1β)-Induced Human Nucleus Pulposus Cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e922497. [Google Scholar] [CrossRef] [PubMed]
- Molina-Pinelo, S.; Pastor, M.D.; Suarez, R.; Romero-Romero, B.; De La Pena, M.G.; Salinas, A.; Garcia-Carbonero, R.; De Miguel, M.J.; Rodriguez-Panadero, F.; Carnero, A.; et al. MicroRNA clusters: Dysregulation in lung adenocarcinoma and COPD. Eur. Respir. J. 2014, 43, 1740–1749. [Google Scholar] [CrossRef]
- Dang, X.; Qu, X.; Wang, W.; Liao, C.; Li, Y.; Zhang, X.; Xu, D.; Baglole, C.J.; Shang, D.; Chang, Y. Bioinformatic analysis of microRNA and mRNA Regulation in peripheral blood mononuclear cells of patients with chronic obstructive pulmonary disease. Respir. Res. 2017, 18, 4. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tang, X.; He, Q.; Yang, X.; Ren, X.; Wen, X.; Zhang, J.; Wang, Y.; Liu, N.; Ma, J. Overexpression of Mitochondria Mediator Gene TRIAP1 by miR-320b Loss Is Associated with Progression in Nasopharyngeal Carcinoma. PLoS Genet. 2016, 12, e1006183. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; You, R.; Yuan, X.; Yang, T.; Samuel, E.; Marcano, D.C.; Sikkema, W.; Tour, J.; Rodriguez, A.; Kheradmand, F.; et al. The microRNA miR-22 inhibits the histone deacetylase HDAC4 to promote TH17 cell–dependent emphysema. Nat. Immunol. 2015, 16, 1185–1194. [Google Scholar] [CrossRef] [Green Version]
- Velasco-Torres, Y.; Ruiz, V.; Montaño, M.; Pérez-Padilla, R.; Falfán-Valencia, R.; Pérez-Ramos, J.; Pérez-Bautista, O.; Ramos, C. Participation of the miR-22-HDAC4-DLCO Axis in Patients with COPD by Tobacco and Biomass. Biomolecules 2019, 9, 837. [Google Scholar] [CrossRef] [Green Version]
- Zhu, K.; Zhou, S.; Xu, A.; Sun, L.; Li, M.; Jiang, H.; Zhang, B.; Zeng, D.; Fei, G.; Wang, R. Microbiota Imbalance Contributes to COPD Deterioration by Enhancing IL-17a Production via miR-122 and miR-30a. Mol. Ther.-Nucleic Acids 2020, 22, 520–529. [Google Scholar] [CrossRef]
- Osei, E.; Florez-Sampedro, L.; Timens, W.; Postma, D.S.; Heijink, I.H.; Brandsma, C.-A. Unravelling the complexity of COPD by microRNAs: It’s a small world after all. Eur. Respir. J. 2015, 46, 807–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Y.; Mao, Z.-D.; Shi, Y.-J.; Liu, Z.-G.; Cao, Q.; Zhang, Q. Comprehensive Analysis of miRNA-mRNA-lncRNA Networks in Non-Smoking and Smoking Patients with Chronic Obstructive Pulmonary Disease. Cell. Physiol. Biochem. 2018, 50, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Rich, H.E.; Alcorn, J.F. IL-17 Strikes a Chord in Chronic Obstructive Pulmonary Disease Exacerbation. Am. J. Respir. Cell Mol. Biol. 2018, 58, 669–670. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Bagam, P.; Pinkston, R.; Singh, D.P.; Batra, S. Cigarette smoke-induced inflammation: NLRP10-mediated mechanisms. Toxicology 2018, 398–399, 52–67. [Google Scholar] [CrossRef]
- Liang, Y.; Zhao, G.; Tang, L.; Zhang, J.; Li, T.; Liu, Z. MiR-100-3p and miR-877-3p regulate overproduction of IL-8 and IL-1β in mesangial cells activated by secretory IgA from IgA nephropathy patients. Exp. Cell Res. 2016, 347, 312–321. [Google Scholar] [CrossRef]
- Han, W.; Ren, X.; Yang, Y.; Li, H.; Zhao, L.; Lin, Z. microRNA -100 functions as a tumor suppressor in non-small cell lung cancer via regulating epithelial-mesenchymal transition and Wnt/β-catenin by targeting HOXA1. Thorac. Cancer 2020, 11, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Akbas, F.; Coskunpinar, E.; Aynacı, E.; Oltulu, Y.M.; Yildiz, P. Analysis of Serum Micro-Rnas as Potential Biomarker in Chronic Obstructive Pulmonary Disease. Exp. Lung Res. 2012, 38, 286–294. [Google Scholar] [CrossRef]
- Yang, S.; Banerjee, S.; Freitas, A.; Sanders, Y.Y.; Ding, Q.; Matalon, S.; Thannickal, V.J.; Abraham, E.; Liu, G. Participation of miR-200 in Pulmonary Fibrosis. Am. J. Pathol. 2012, 180, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Moimas, S.; Salton, F.; Kosmider, B.; Ring, N.; Volpe, M.C.; Bahmed, K.; Braga, L.; Rehman, M.; Vodret, S.; Graziani, M.L.; et al. miR-200 family members reduce senescence and restore idiopathic pulmonary fibrosis type II alveolar epithelial cell transdifferentiation. ERJ Open Res. 2019, 5, 00138–02019. [Google Scholar] [CrossRef]
- Zhu, M.; An, Y.; Zhang, X.; Wang, Z.; Duan, H. Experimental pulmonary fibrosis was suppressed by microRNA-506 through NF-kappa-mediated apoptosis and inflammation. Cell Tissue Res. 2019, 378, 255–265. [Google Scholar] [CrossRef]
- Makiguchi, T.; Yamada, M.; Yoshioka, Y.; Sugiura, H.; Koarai, A.; Chiba, S.; Fujino, N.; Tojo, Y.; Ota, C.; Kubo, H.; et al. Serum extracellular vesicular miR-21-5p is a predictor of the prognosis in idiopathic pulmonary fibrosis. Respir. Res. 2016, 17, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Kumar, M.; Choudhury, S.N.; Buscaglia, L.E.B.; Barker, J.R.; Kanakamedala, K.; Liu, M.-F.; Li, Y. Loss of the miR-21 allele elevates the expression of its target genes and reduces tumorigenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 10144–10149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-F.; Jing, Y.; Hao, J.; Frankfort, N.C.; Zhou, X.; Shen, B.; Liu, X.; Wang, L.; Li, R. MicroRNA-21 in the pathogenesis of acute kidney injury. Protein Cell 2013, 4, 813–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis-Dusenbery, B.; Hilyard, A.C.; Lagna, G.; Hata, A. SMAD proteins control DROSHA-mediated microRNA maturation. Nat. Cell Biol. 2008, 454, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Jin, K. Peripheral Circulating Exosomal miRNAs Potentially Contribute to the Regulation of Molecular Signaling Networks in Aging. Int. J. Mol. Sci. 2020, 21, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Anca, M.; Fenoglio, C.; Serpente, M.; Arosio, B.; Cesari, M.; Scarpini, E.A.; Galimberti, D. Exosome Determinants of Physiological Aging and Age-Related Neurodegenerative Diseases. Front. Aging Neurosci. 2019, 11, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Tahara, H. The role of exosomes and microRNAs in senescence and aging. Adv. Drug Deliv. Rev. 2013, 65, 368–375. [Google Scholar] [CrossRef]
- Roth, M.D.; Connett, J.E.; D’Armiento, J.M.; Foronjy, R.F.; Friedman, P.J.; Goldin, J.; Louis, T.A.; Mao, J.T.; Muindi, J.R.; O’Connor, G.; et al. Feasibility of Retinoids for the Treatment of Emphysema Study. Chest 2006, 130, 1334–1345. [Google Scholar] [CrossRef] [PubMed]
- Agudelo, C.W.; Kumley, B.K.; Area-Gomez, E.; Xu, Y.; Dabo, A.J.; Geraghty, P.; Campos, M.; Foronjy, R.; Garcia-Arcos, I. Decreased surfactant lipids correlate with lung function in chronic obstructive pulmonary disease (COPD). PLoS ONE 2020, 15, e0228279. [Google Scholar] [CrossRef]
- Maremanda, K.P.; Sundar, I.K.; Li, D.; Rahman, I. Age-Dependent Assessment of Genes Involved in Cellular Senescence, Telomere, and Mitochondrial Pathways in Human Lung Tissue of Smokers, COPD, and IPF: Associations With SARS-CoV-2 COVID-19 ACE2-TMPRSS2-Furin-DPP4 Axis. Front. Pharmacol. 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Sundar, I.K.; Yin, Q.; Baier, B.S.; Yan, L.; Mazur, W.; Li, D.; Susiarjo, M.; Rahman, I. DNA methylation profiling in peripheral lung tissues of smokers and patients with COPD. Clin. Epigenet. 2017, 9, 1–18. [Google Scholar] [CrossRef]
- Dooner, M.S.; Stewart, C.; Deng, Y.; Papa, E.; Pereira, M.; Del Tatto, M.; Johnson, S.; Wen, S.; Amaral, A.; Aliotta, J.; et al. Daily rhythms influence the ability of lung-derived extracellular vesicles to modulate bone marrow cell phenotype. PLoS ONE 2018, 13, e0207444. [Google Scholar] [CrossRef]
- Singh, K.P.; Maremanda, K.P.; Li, D.; Rahman, I. Exosomal microRNAs are novel circulating biomarkers in cigarette, waterpipe smokers, E-cigarette users and dual smokers. BMC Med. Genom. 2020, 13, 1–20. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Nakaya, A. The KEGG databases at GenomeNet. Nucleic Acids Res. 2002, 30, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; Smyth, G.K. Moderated statistical tests for assessing differences in tag abundance. Bioinformatics 2007, 23, 2881–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Term | Ontology | n | p-Value * |
|---|---|---|---|---|
| NS vs. COPD | ||||
| GO:0016043 | Cellular component organization | BP | 17 | 0.183535762 |
| GO:0071840 | Cellular component organization or biogenesis | BP | 17 | 0.183535762 |
| GO:0061024 | Membrane organization | BP | 3 | 0.149797571 |
| GO:0051259 | Protein complex oligomerization | BP | 3 | 0.149797571 |
| GO:0006720 | Isoprenoid metabolic process | BP | 2 | 0.101214575 |
| GO:0051186 | Co-factor metabolic process | BP | 2 | 0.101214575 |
| GO:0051188 | Co-factor biosynthetic process | BP | 2 | 0.101214575 |
| GO:0051262 | Protein tetramerization | BP | 2 | 0.101214575 |
| GO:0008299 | Isoprenoid biosynthetic process | BP | 2 | 0.101214575 |
| GO:0006732 | Co-enzyme metabolic process | BP | 1 | 0.051282051 |
| GO:0009108 | Co-enzyme biosynthetic process | BP | 1 | 0.051282051 |
| GO:0006733 | Oxidoreduction coenzyme metabolic process | BP | 1 | 0.051282051 |
| GO:0043687 | Post-translational protein modification | BP | 1 | 0.051282051 |
| GO:1901661 | Quinone metabolic process | BP | 1 | 0.051282051 |
| GO:0042181 | Ketone biosynthetic process | BP | 1 | 0.051282051 |
| GO:0051290 | Protein heterotetramerization | BP | 1 | 0.051282051 |
| GO:0051291 | Protein heterooligomerization | BP | 1 | 0.051282051 |
| GO:0006743 | Ubiquinone metabolic process | BP | 1 | 0.051282051 |
| GO:0006744 | Ubiquinone biosynthetic process | BP | 1 | 0.051282051 |
| GO:1901663 | Quinone biosynthetic process | BP | 1 | 0.051282051 |
| GO:0031974 | Membrane-enclosed lumen | CC | 11 | 0.007422402 |
| GO:0070013 | Intracellular organelle lumen | CC | 11 | 0.007422402 |
| GO:0030659 | Cytoplasmic vesicle membrane | CC | 5 | 0.24291498 |
| GO:0044431 | Golgi apparatus part | CC | 5 | 0.24291498 |
| GO:0044429 | Mitochondrial part | CC | 3 | 0.149797571 |
| GO:1902494 | Catalytic complex | CC | 3 | 0.149797571 |
| GO:1990234 | Transferase complex | CC | 2 | 0.101214575 |
| GO:0005788 | Endoplasmic reticulum lumen | CC | 2 | 0.101214575 |
| GO:0030133 | Transport vesicle | CC | 2 | 0.101214575 |
| GO:0005802 | Trans-Golgi network | CC | 1 | 0.051282051 |
| GO:0030135 | Coated vesicle | CC | 1 | 0.051282051 |
| GO:0030136 | Clathrin-coated vesicle | CC | 1 | 0.051282051 |
| GO:0030662 | Coated vesicle membrane | CC | 1 | 0.051282051 |
| GO:0030665 | Clathrin-coated vesicle membrane | CC | 1 | 0.051282051 |
| GO:0005759 | Mitochondrial matrix | CC | 1 | 0.051282051 |
| GO:0016765 | Transferase activity, transferring alkyl, or aryl (other than methyl) groups | MF | 2 | 0.101214575 |
| GO:0046982 | Protein heterodimerization | MF | 1 | 0.051282051 |
| GO:0000010 | Trans-hexaprenyltranstransferase activity | MF | 1 | 0.051282051 |
| GO:0050347 | Trans-octaprenyltranstransferase activity | MF | 1 | 0.051282051 |
| Sm vs. COPD | ||||
| GO:0007623 | Circadian rhythm | BP | 4 | 0.114285714 |
| GO:0048511 | Rhythmic process | BP | 4 | 0.114285714 |
| GO:1901566 | Organonitrogen compound biosynthetic process | BP | 4 | 0.114285714 |
| GO:1901135 | Carbohydrate derivative metabolic process | BP | 3 | 0.085714286 |
| GO:1901137 | Carbohydrate derivative biosynthetic process | BP | 2 | 0.057142857 |
| GO:0006022 | Aminoglycan metabolic process | BP | 1 | 0.028571429 |
| GO:0006023 | Aminoglycan biosynthetic process | BP | 1 | 0.028571429 |
| GO:0006024 | Glycosaminoglycan biosynthetic process | BP | 1 | 0.028571429 |
| GO:0005794 | Glycosaminoglycan metabolic process | BP | 1 | 0.028571429 |
| GO:0030203 | Bounding membrane of organelle | CC | 8 | 0.228571429 |
| GO:0001904 | Organelle sub-compartment | CC | 6 | 0.171428571 |
| GO:0044431 | Golgi apparatus | CC | 6 | 0.171428571 |
| GO:0098588 | Golgi sub-compartment | CC | 5 | 0.142857143 |
| GO:0098791 | Golgi membrane | CC | 4 | 0.114285714 |
| GO:0016740 | Transferase activity | MF | 5 | 0.142857143 |
| GO:0016782 | Transferase activity, transferring sulfur-containing groups | MF | 1 | 0.028571429 |
| GO:0008146 | Sulfotransferase activity | MF | 1 | 0.028571429 |
| GO:0034483 | Heparan sulfate sulfotransferase activity | MF | 1 | 0.028571429 |
| GO:0033871 | (Heparan sulfate)-glucosamine-3-sulfotransferase-2-activity | MF | 1 | 0.028571429 |
| NS vs. IPF | ||||
| GO:0009636 | Response to toxic substance | BP | 2 | 0.142682927 |
| GO:0097324 | Melanocyte migration | BP | 1 | 0.073170732 |
| GO:0097324 | Melanosome organization | BP | 1 | 0.073170732 |
| GO:0014031 | Mesenchymal cell development | BP | 1 | 0.073170732 |
| GO:0034204 | Lipid transport | BP | 1 | 0.073170732 |
| GO:0044429 | Mitochondrial part | CC | 3 | 0.010787992 |
| GO:0005739 | Mitochondrion | CC | 5 | 0.034709193 |
| GO:0030136 | Clathrin-coated vesicle | CC | 1 | 0.073170732 |
| GO:0000785 | Chromatin | CC | 1 | 0.073170732 |
| GO:0005766 | Primary lysosome | CC | 1 | 0.073170732 |
| GO:0000010 | Trans-hexaprenyltranstransferase activity | MF | 1 | 0.073170732 |
| GO:0050347 | Trans-octaprenyltranstransferase activity | MF | 1 | 0.073170732 |
| GO:0016887 | ATPase activity | MF | 1 | 0.073170732 |
| GO:0070412 | R-SMAD binding | MF | 1 | 0.073170732 |
| ID | Term | Ontology | n | p-Value * |
|---|---|---|---|---|
| NS vs. COPD | ||||
| GO:0048514 | Blood vessel morphogenesis | BP | 12 | 0.068181818 |
| GO:0050808 | Synapse organization | BP | 11 | 0.0625 |
| GO:0051962 | Positive regulation of nervous system development | BP | 11 | 0.0625 |
| GO:0044089 | Positive regulation of cellular component biogenesis | BP | 10 | 0.056818182 |
| GO:0044430 | Cytoskeletal part | CC | 9 | 0.051136364 |
| GO:0001525 | Angiogenesis | BP | 8 | 0.045454545 |
| GO:0050803 | Regulation of synapse structure or activity | BP | 8 | 0.045454545 |
| GO:0050807 | Regulation of synapse organization | BP | 8 | 0.045454545 |
| GO:0015630 | Microtubule cytoskeleton | CC | 8 | 0.045454545 |
| GO:0038023 | Signaling receptor activity | MF | 8 | 0.045454545 |
| GO:0060089 | Molecular transducer activity | MF | 8 | 0.045454545 |
| GO:0004888 | Transmembrane signaling receptor activity | MF | 7 | 0.039772727 |
| GO:0004930 | G-protein-coupled receptor activity | MF | 7 | 0.039772727 |
| GO:0019932 | Second-messenger-mediated signaling | BP | 6 | 0.034090909 |
| GO:0007416 | Synapse assembly | BP | 6 | 0.034090909 |
| GO:0045765 | Regulation of angiogenesis | BP | 5 | 0.028409091 |
| GO:1901342 | Regulation of vasculature development | BP | 5 | 0.028409091 |
| GO:0051963 | Regulation of synapse assembly | BP | 5 | 0.028409091 |
| GO:0051965 | Positive regulation of synapse assembly | BP | 4 | 0.022727273 |
| GO:0019722 | Calcium-mediated signaling | BP | 4 | 0.022727273 |
| GO:0005815 | Microtubule organizing center | CC | 4 | 0.022727273 |
| GO:0005813 | Centrosome | CC | 4 | 0.022727273 |
| GO:0016525 | Negative regulation of angiogenesis | BP | 3 | 0.017045455 |
| GO:1901343 | Negative regulation of vasculature development | BP | 3 | 0.017045455 |
| GO:2000181 | Negative regulation of blood vessel morphogenesis | BP | 3 | 0.017045455 |
| GO:0033173 | Calcineurin-NFAT signaling cascade | BP | 2 | 0.011363636 |
| GO:0048016 | Inositol-phosphate-mediated signaling | BP | 2 | 0.011363636 |
| GO:0097720 | Calcineurin-mediated signaling | BP | 2 | 0.011363636 |
| Sm vs. COPD | ||||
| GO:0048514 | Blood vessel morphogenesis | BP | 11 | 0.071428571 |
| GO:0044087 | Regulation of cellular component biogenesis | BP | 11 | 0.071428571 |
| GO:0044089 | Positive regulation of cellular component biogenesis | BP | 10 | 0.064935065 |
| GO:0051962 | Positive regulation of nervous system development | BP | 10 | 0.064935065 |
| GO:0001525 | Angiogenesis | BP | 8 | 0.051948052 |
| GO:0050803 | Regulation of synapse structure or activity | BP | 8 | 0.051948052 |
| GO:0050807 | Regulation of synapse organization | BP | 8 | 0.051948052 |
| GO:0044430 | Cytoskeletal part | CC | 7 | 0.045454545 |
| GO:0038023 | Signaling receptor activity | MF | 7 | 0.045454545 |
| GO:0060089 | Molecular transducer activity | MF | 7 | 0.045454545 |
| GO:0019932 | Second-messenger-mediated signaling | BP | 6 | 0.038961039 |
| GO:0007416 | Synapse assembly | BP | 6 | 0.038961039 |
| GO:0015630 | Microtubule cytoskeleton | CC | 6 | 0.038961039 |
| GO:0004888 | Transmembrane signaling receptor activity | MF | 6 | 0.038961039 |
| GO:0045765 | Regulation of angiogenesis | BP | 5 | 0.032467532 |
| GO:1901342 | Regulation of vasculature development | BP | 5 | 0.032467532 |
| GO:0051963 | Regulation of synapse assembly | BP | 5 | 0.032467532 |
| GO:0051965 | Positive regulation of synapse assembly | BP | 4 | 0.025974026 |
| GO:0019722 | Calcium-mediated signaling | BP | 4 | 0.025974026 |
| GO:0016525 | Negative regulation of angiogenesis | BP | 3 | 0.019480519 |
| GO:1901343 | Negative regulation of vasculature development | BP | 3 | 0.019480519 |
| GO:2000181 | Negative regulation of blood vessel morphogenesis | BP | 3 | 0.019480519 |
| GO:0005815 | Microtubule organizing center | CC | 3 | 0.019480519 |
| GO:0005813 | Centrosome | CC | 3 | 0.019480519 |
| GO:0004930 | G-protein-coupled receptor activity | MF | 3 | 0.019480519 |
| GO:0033173 | Calcineurin-NFAT signaling cascade | BP | 2 | 0.012987013 |
| GO:0048016 | Inositol-phosphate-mediated signaling | BP | 2 | 0.012987013 |
| GO:0097720 | Calcineurin-mediated signaling | BP | 2 | 0.012987013 |
| NS vs. IPF | ||||
| GO:0065008 | Regulation of biological quality | BP | 44 | 0.0327192 |
| GO:0007399 | Nervous system development | BP | 36 | 0.006858696 |
| GO:0048878 | Chemical homeostasis | BP | 18 | 0.003415 |
| GO:0030030 | Cell projection organization | BP | 17 | 0.012599483 |
| GO:0120036 | Plasma-membrane-bounded cell projection organization | BP | 17 | 0.01259943 |
| GO:0044459 | Plasma membrane region | CC | 10 | 0.028105097 |
| GO:0007416 | Synapse assembly | BP | 7 | 0.006212841 |
| GO:0030424 | Axon | CC | 7 | 0.0488352 |
| GO:0150034 | Distal axon | CC | 6 | 0.0305685 |
| GO:0031349 | Positive regulation of defense response | BP | 5 | 0.016747 |
| GO:0044306 | Neuron projection terminus | CC | 4 | 0.007341699 |
| GO:0008092 | Cytoskeletal protein binding | MF | 4 | 0.0073417 |
| GO:0004930 | G-protein-coupled receptor activity | MF | 4 | 0.083811139 |
| GO:0051965 | Positive regulation of synapse assembly | BP | 4 | 0.007341699 |
| GO:0010863 | Positive regulation of phospholipase C activity | BP | 2 | 0.01653348 |
| GO:0043235 | Receptor complex | CC | 2 | 0.01653348 |
| GO:0023026 | MHC class II protein complex binding | MF | 2 | 0.0165335 |
| GO:0005096 | GTPase activator activity | MF | 2 | 0.0165335 |
| GO:1903997 | Positive regulation of non-membrane spanning protein tyrosine kinase activity | BP | 2 | 0.01653348 |
| KEGG Pathway | Selected Pathway |
|---|---|
| COPD ƚ | |
| path:hsa00900 | Terpenoid backbone biosynthesis |
| path:hsa04920 | Adipocytokine signaling pathway |
| path:hsa00520 | Amino sugar and nucleotide sugar metabolism |
| path:hsa04152 | AMPK signaling pathway |
| path:hsa04371 | Apelin signaling pathway |
| path:hsa04140 | Autophagy-animal |
| path:hsa04136 | Autophagy-other |
| path:hsa01040 | Biosynthesis of unsaturated fatty acids |
| path:hsa04024 | cAMP signaling pathway |
| path:hsa04218 | Cellular senescence |
| path:hsa04062 | Chemokine signaling pathway |
| path:hsa00534 * | Glycosaminoglycan biosynthesis |
| IPF # | |
| path:hsa00900 | Terpenoid backbone biosynthesis |
| path:hsa04920 | Adipocytokine signaling pathway |
| path:hsa00520 | Amino sugar and nucleotide sugar metabolism |
| path:hsa04371 | Apelin signaling pathway |
| path:hsa04140 | Autophagy-animal |
| path:hsa04136 | Autophagy-other |
| path:hsa01040 | Biosynthesis of unsaturated fatty acids |
| path:hsa04218 | Cellular senescence |
| path:hsa04062 | Chemokine signaling pathway |
| path: hsa05206 | microRNAs in cancer |
| KEGG Pathway | Selected Pathway |
|---|---|
| COPD ƚ | |
| path:hsa04520 | Adherens junction |
| path:hsa04920 | Adipocytokine signaling pathway |
| path:hsa04261 | Adrenergic signaling in cardiomyocytes |
| path:hsa04960 | Aldosterone-regulated sodium reabsorption |
| path:hsa00520 | Amino sugar and nucleotide sugar metabolism |
| path:hsa04215 | Apoptosis—multiple species |
| path:hsa05310 | Asthma |
| path:hsa05100 | Bacterial invasion of epithelial cells |
| path:hsa01040 | Biosynthesis of unsaturated fatty acids |
| path:hsa04260 | Cardiac muscle contraction |
| path:hsa04022 | cGMP-PKG signaling pathway |
| IPF # | |
| path:hsa04972 * | Pancreatic secretion |
| path:hsa04970 * | Salivary secretion |
| path:hsa04911 * | Insulin secretion |
| path:hsa05416 * | Viral myocarditis |
| path:hsa05310 | Asthma |
| path:hsa01040 | Biosynthesis of unsaturated fatty acids |
| path:hsa04022 | cGMP-PKG signaling pathway |
| path:hsa04014 * | Ras signaling pathway |
| path:hsa04727 * | GABAergic synapse |
| path:hsa05033 * | Nicotine addiction |
| path:hsa04722 * | Neurotrophin signaling pathway |
| path:hsa04010 * | MAPK signaling pathway |
| path:hsa04151 | PI3K-Akt signaling pathway |
| Characteristics | Non-Smokers | Smokers | COPD/Emphysema | IPF | p-Value * |
|---|---|---|---|---|---|
| BALF | |||||
| N | 8 | 8 | 16 | 8 | |
| Age (years), mean (SD) | 49.6 (17.3) | 57.4 (8.9) | 65.9 (13.3) | 76.5 (11.4) | 0.0029 |
| Gender | 0.2952 | ||||
| Male n (%) | 3 (37.5) | 2 (25) | 7 (38.9) | 6 (75) | |
| Not specified | 0 | 0 | 2 | 0 | |
| Smoking status | 0.9719 | ||||
| Current smoker | 0 | 6 | 2 | 0 | |
| Ex-smoker | 0 | 2 | 6 | 4 | |
| N/A | 0 | 0 | 7 | 0 | |
| Lung Tissue | |||||
| N | 8 | 8 | 8 | 8 | |
| Age (years), mean (SD) | 48.3 (16.3) | 53.8 (15.4) | 59.1 (9.9) | 68.9 (9.6) | 0.0688 |
| Gender | 0.981 | ||||
| Male n (%) | 4 (50) | 6 (75) | 3 (37.5) | 5 (62.5) | |
| N/A | 2 | 1 | 0 | 0 | |
| Smoking status | >0.9999 | ||||
| Current smoker | 0 | 7 | 2 | 0 | |
| Ex-smoker | 3 | 1 | 4 | 6 | |
| N/A | 0 | 0 | 1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, G.; Maremanda, K.P.; Campos, M.; Chand, H.S.; Li, F.; Hirani, N.; Haseeb, M.A.; Li, D.; Rahman, I. Distinct Exosomal miRNA Profiles from BALF and Lung Tissue of COPD and IPF Patients. Int. J. Mol. Sci. 2021, 22, 11830. https://doi.org/10.3390/ijms222111830
Kaur G, Maremanda KP, Campos M, Chand HS, Li F, Hirani N, Haseeb MA, Li D, Rahman I. Distinct Exosomal miRNA Profiles from BALF and Lung Tissue of COPD and IPF Patients. International Journal of Molecular Sciences. 2021; 22(21):11830. https://doi.org/10.3390/ijms222111830
Chicago/Turabian StyleKaur, Gagandeep, Krishna Prahlad Maremanda, Michael Campos, Hitendra S. Chand, Feng Li, Nikhil Hirani, M. A. Haseeb, Dongmei Li, and Irfan Rahman. 2021. "Distinct Exosomal miRNA Profiles from BALF and Lung Tissue of COPD and IPF Patients" International Journal of Molecular Sciences 22, no. 21: 11830. https://doi.org/10.3390/ijms222111830
APA StyleKaur, G., Maremanda, K. P., Campos, M., Chand, H. S., Li, F., Hirani, N., Haseeb, M. A., Li, D., & Rahman, I. (2021). Distinct Exosomal miRNA Profiles from BALF and Lung Tissue of COPD and IPF Patients. International Journal of Molecular Sciences, 22(21), 11830. https://doi.org/10.3390/ijms222111830