Generation of High-Yield, Functional Oligodendrocytes from a c-myc Immortalized Neural Cell Line, Endowed with Staminal Properties

 , ,
, , 
Abstract
:1. Introduction
2. Results
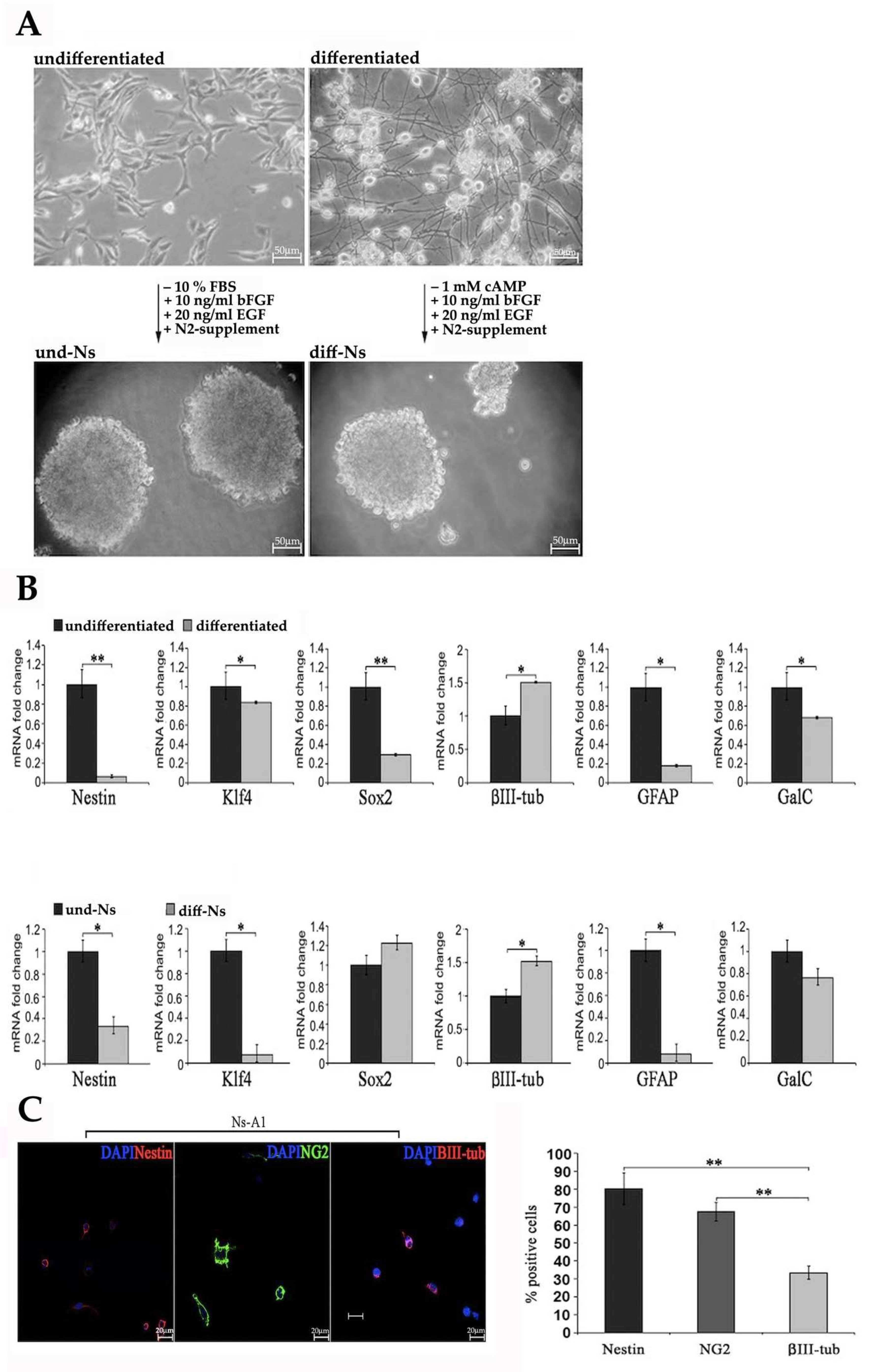
2.1. Expression of Staminal and Neural Markers by und-Ns and diff-Ns
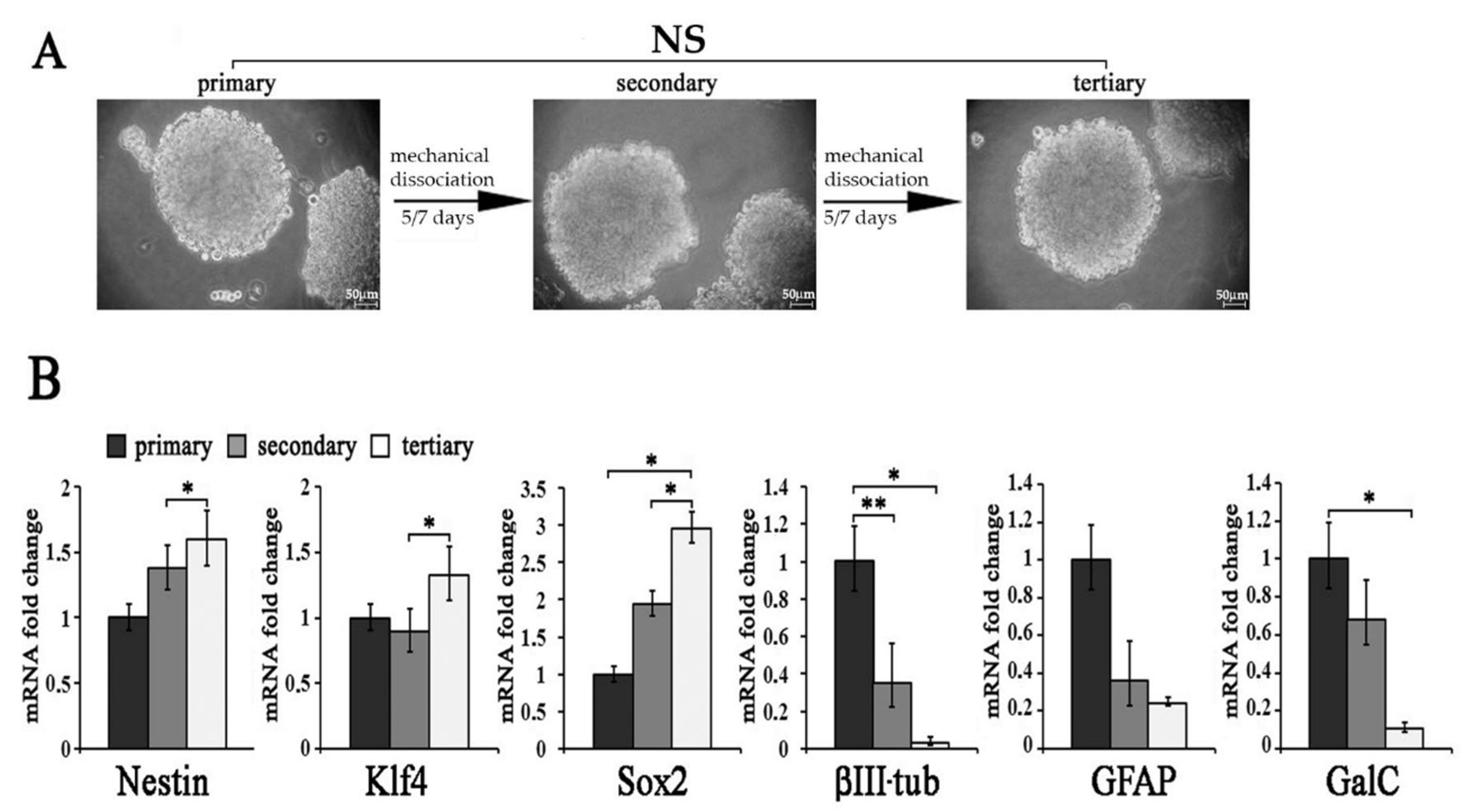
2.2. Molecular Characterization of und-Ns Subcultures
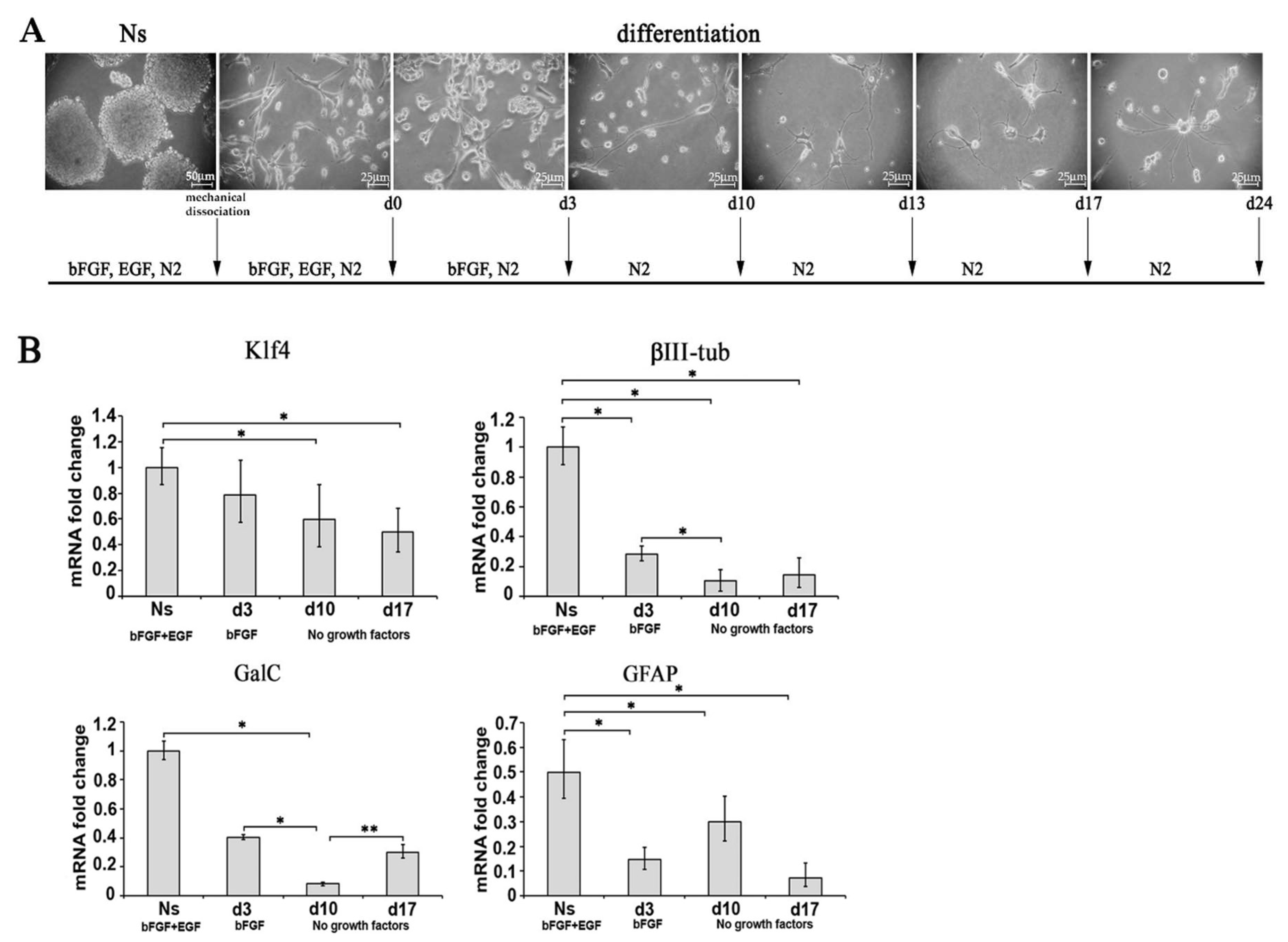
2.3. Differentiation and Characterization of Primary Neurosphere Derived from A1 Undifferentiated Cells (Ns)
2.3.1. Characterization of Neurosphere-Derived A1 Cells by Real-Time PCR
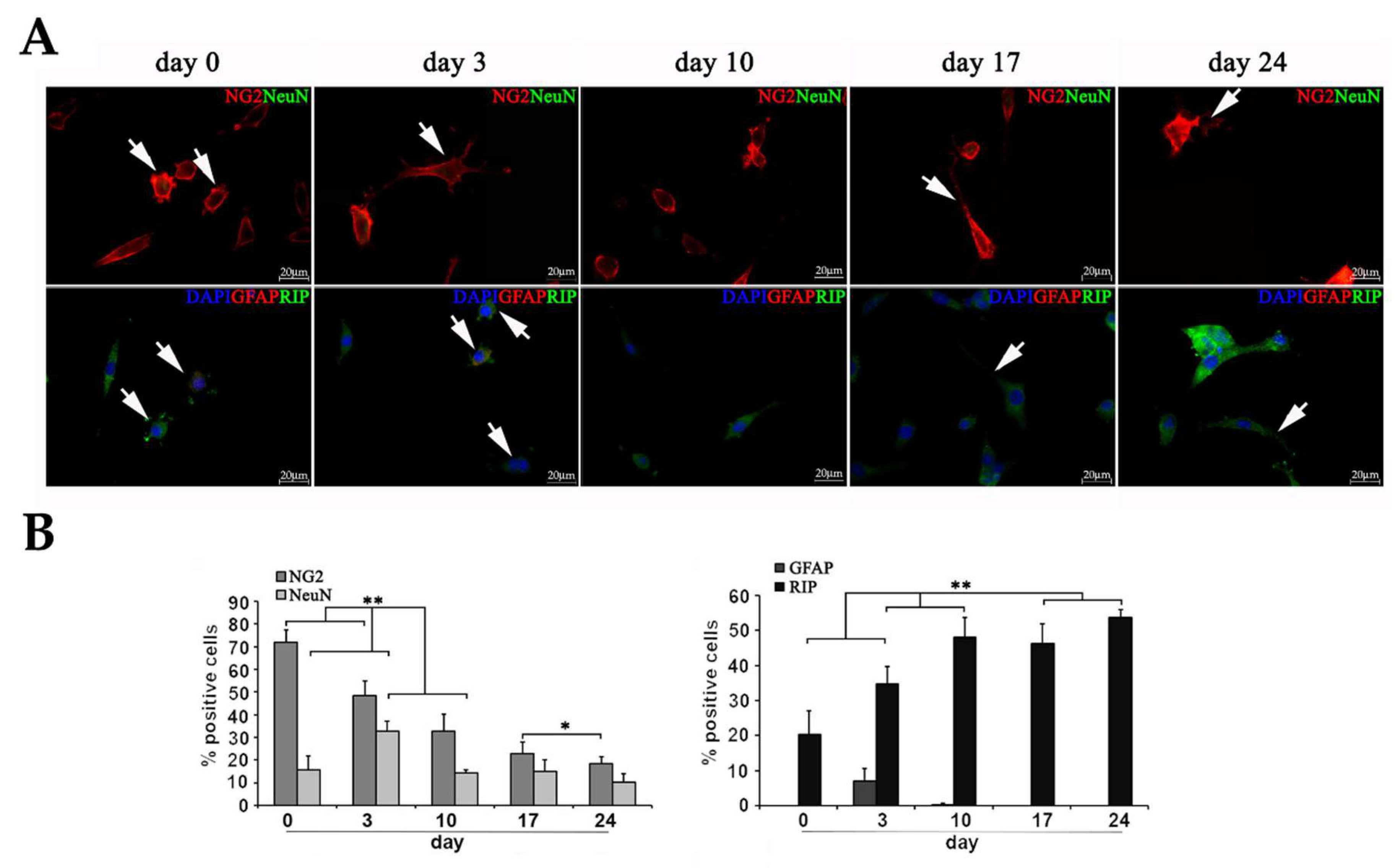
2.3.2. Characterization of diff-Ns by Immunofluorescence
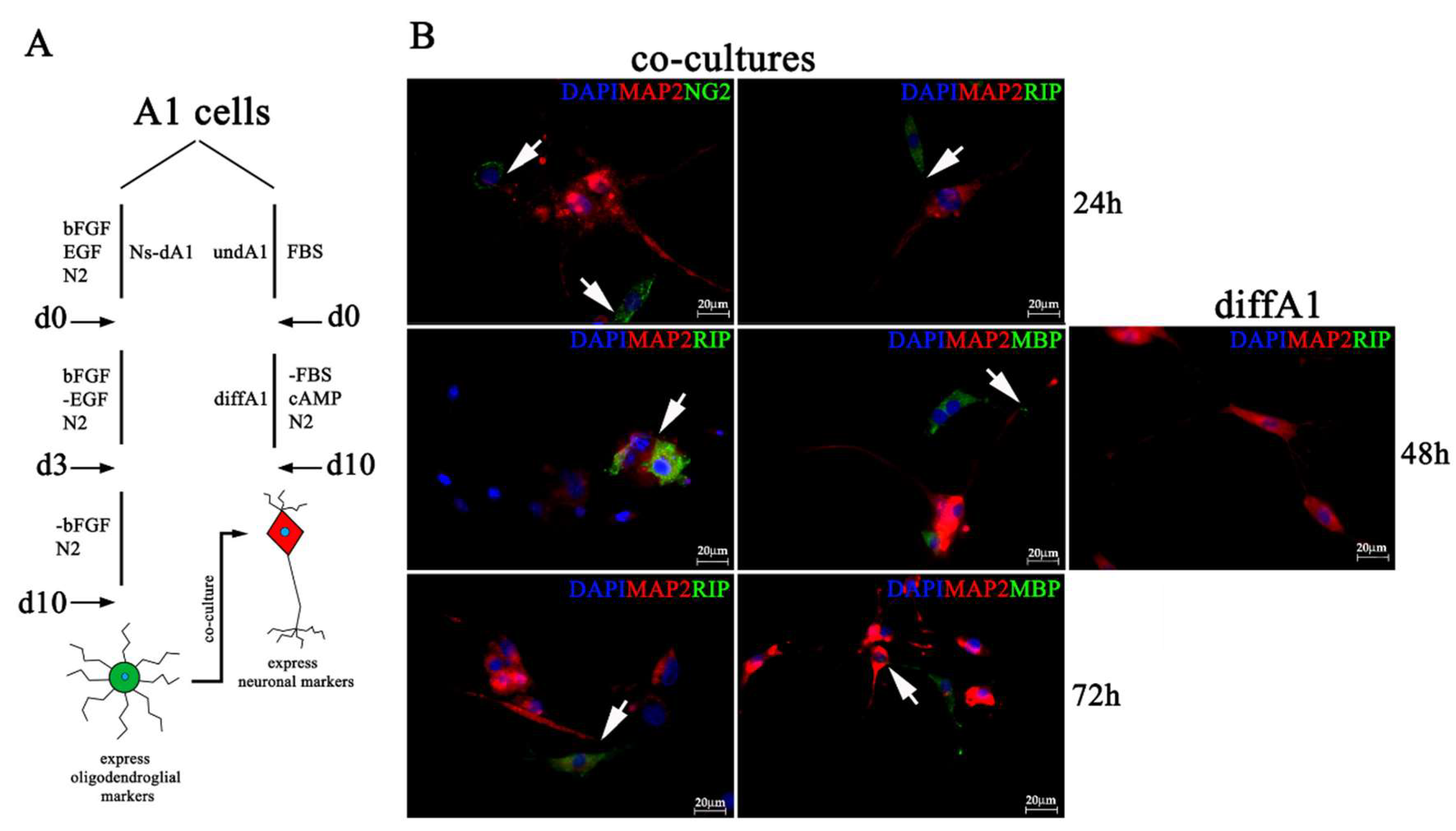
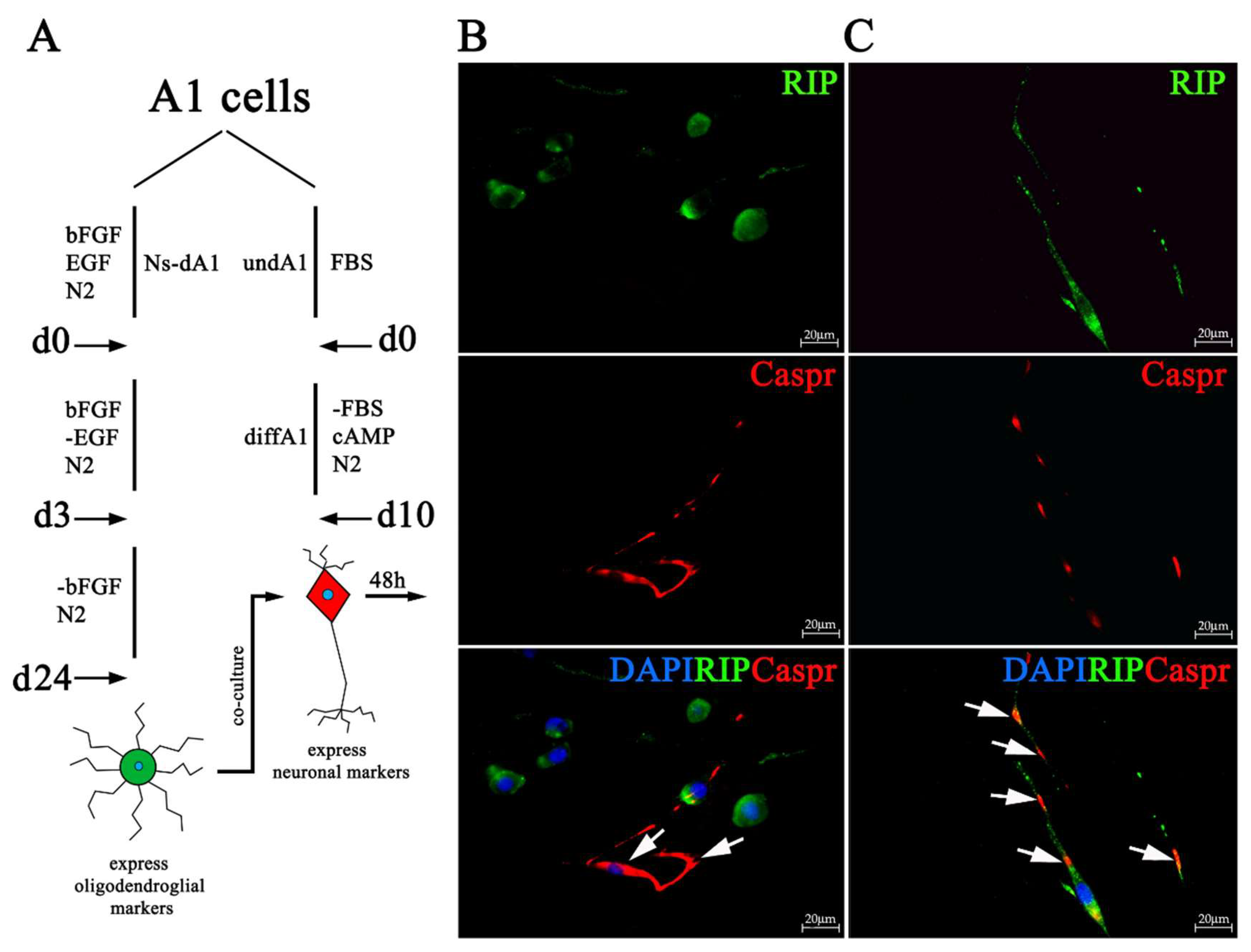
2.4. Neuronal Differentiated A1 Cells When Co-Cultured With Differentiated Ns-A1 Express Oligodendroglial Markers, Redistribute Caspr/Paranodin to Paranodal Regions
3. Discussion
4. Materials and Methods
4.1. A1 Cell Culture Conditions
4.2. Neurosphere-Derived A1 Cell Differentiation
4.3. Co-Culture of Neuronal Differentiated A1 Cells and Differentiated Ns-A1 Expressing Oligodendroglial Markers
4.4. Primary CNS Cell Cultures
4.5. RNA Extraction, Reverse Transcription and Real-Time PCR Analysis
4.6. Immunocytochemistry
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Caspr | Contactin-associated protein |
| bFGF | fibroblast growth factor 2 |
| cAMP | Dibutyryl cyclic adenosine 3,5-monophosphate |
| CNS | Central Nervous System |
| DAPI | 4,6-diamidino-2-phenylindole |
| Diff | Differentiated A1 cells |
| Diff-Ns | primary neurosphere derived from undifferentiated A1 cells |
| EGF | Epidermal Growth Factor |
| GFAP | Glial fibrillary acidic protein |
| GalC | Galactosylcerebrosidase |
| Hprt | Hypoxanthine phosphoribosyltransferase |
| iPS | Induced Pluripotent Stem cells |
| Klf4 | Kruppel-like factor 4 |
| MAP2 | Micro-tubule-associated protein 2 |
| MBP | Myelin basic protein |
| NeuN | Neuronal nuclei |
| NG2 | Neural/glial antigen 2 |
| Ns | Neurospheres |
| NSCs | Neural Stem Cells |
| RIP | Receptor-interacting protein |
| Sox2 | Sex determining region Y-box 2 |
| Und | Undifferentiated A1 cells |
| Und-Ns | Undifferentiated A1 cells-derived neurospheres |
References
- Reynolds, B.A.; Rietze, R.L. Neural stem cells and neurospheres—Re-evaluating the relationship. Nat. Methods 2005, 2, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Rietze, R.L.; Reynolds, B.A. Neural stem cell isolation and characterization. Methods Enzymol. 2006, 419, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Gil-Perotín, S.; Duran-Moreno, M.; Cebrián-Silla, A.; Ramírez, M.; García-Belda, P.; García-Verdugo, J.M. Adult neural stem cells from the subventricular zone: A review of the neurosphere assay. Anat. Rec. (Hoboken) 2013, 296, 1435–1452. [Google Scholar] [CrossRef]
- Bazán, E.; Alonso, F.J.; Redondo, C.; López-Toledano, M.A.; Alfaro, J.M.; Reimers, D.; Herranz, A.S.; Paíno, C.L.; Serrano, A.B.; Cobacho, N.; et al. In vitro and in vivo characterization of neural stem cells. Histol. Histopathol. 2004, 19, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, L.; Binda, E. Concise review: Self-renewal in the central nervous system: Neural stem cells from embryo to adult. Stem Cells Transl. Med. 2012, 1, 298–308. [Google Scholar] [CrossRef]
- Merkle, F.T.; Alvarez-Buylla, A. Neural stem cells in mammalian development. Curr. Opin. Cell Biol. 2006, 18, 704–709. [Google Scholar] [CrossRef]
- De Filippis, L.; Ferrari, D.; Rota Nodari, L.; Amati, B.; Snyder, E.; Vescovi, A.L. Immortalization of human neural stem cells with the c-myc mutant T58A. PLoS ONE 2008, 3, e3310. [Google Scholar] [CrossRef]
- De Filippis, L.; Lamorte, G.; Snyder, E.Y.; Malgaroli, A.; Vescovi, A.L. A novel, immortal, and multipotent human neural stem cell line generating functional neurons and oligodendrocytes. Stem Cells 2007, 25, 2312–2321. [Google Scholar] [CrossRef]
- Cacci, E.; Villa, A.; Parmar, M.; Cavallaro, M.; Mandahl, N.; Lindvall, O.; Martinez-Serrano, A.; Kokaia, Z. Generation of human cortical neurons from a new immortal fetal neural stem cell line. Exp. Cell Res. 2007, 313, 588–601. [Google Scholar] [CrossRef] [Green Version]
- Villa, A.; Liste, I.; Courtois, E.T.; Seiz, E.G.; Ramos, M.; Meyer, M.; Juliusson, B.; Kusk, P.; Martínez-Serrano, A. Generation and properties of a new human ventral mesencephalic neural stem cell line. Exp. Cell Res. 2009, 315, 1860–1874. [Google Scholar] [CrossRef]
- Bartlett, P.F.; Brooker, G.J.; Faux, C.H.; Dutton, R.; Murphy, M.; Turnley, A.; Kilpatrick, T.J. Regulation of neural stem cell differentiation in the forebrain. Immunol. Cell Biol. 1998, 76, 414–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, D.; Jacob, M. Twisting and ignoring facts on hydroxyethyl starch is not very helpful. Scand. J. Trauma Resusc. Emerg. Med. 2013, 21, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.V. c-Myc target genes involved in cell growth, apoptosis, and metabolism. Mol. Cell. Biol. 1999, 19, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoepfler, P.S. Why myc? An unexpected ingredient in the stem cell cocktail. Cell Stem Cell 2008, 2, 18–21. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.M.; Dalton, S. The cell cycle and Myc intersect with mechanisms that regulate pluripotency and reprogramming. Cell Stem Cell 2009, 5, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Zörnig, M.; Evan, G.I. Cell cycle: On target with Myc. Curr. Biol. 1996, 6, 1553–1556. [Google Scholar] [CrossRef] [Green Version]
- Wernig, M.; Meissner, A.; Cassady, J.P.; Jaenisch, R. c-Myc is dispensable for direct reprogramming of mouse fibroblasts. Cell Stem Cell 2008, 2, 10–12. [Google Scholar] [CrossRef] [Green Version]
- Pei, D. Regulation of pluripotency and reprogramming by transcription factors. J. Biol. Chem. 2009, 284, 3365–3369. [Google Scholar] [CrossRef] [Green Version]
- Pino-Barrio, M.J.; García-García, E.; Menéndez, P.; Martínez-Serrano, A. V-myc immortalizes human neural stem cells in the absence of pluripotency-associated traits. PLoS ONE 2015, 10, e0118499. [Google Scholar] [CrossRef] [Green Version]
- Vescovi, A.L.; Parati, E.A.; Gritti, A.; Poulin, P.; Ferrario, M.; Wanke, E.; Frölichsthal-Schoeller, P.; Cova, L.; Arcellana-Panlilio, M.; Colombo, A.; et al. Isolation and cloning of multipotential stem cells from the embryonic human CNS and establishment of transplantable human neural stem cell lines by epigenetic stimulation. Exp. Neurol. 1999, 156, 71–83. [Google Scholar] [CrossRef]
- Colucci-D’Amato, G.L.; Tino, A.; Pernas-Alonso, R.; ffrench-Mullen, J.M.; di Porzio, U. Neuronal and glial properties coexist in a novel mouse CNS immortalized cell line. Exp. Cell Res. 1999, 252, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Einheber, S.; Zanazzi, G.; Ching, W.; Scherer, S.; Milner, T.A.; Peles, E.; Salzer, J.L. The axonal membrane protein Caspr, a homologue of neurexin IV, is a component of the septate-like paranodal junctions that assemble during myelination. J. Cell Biol. 1997, 139, 1495–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindley, R.M.; Hawcutt, D.B.; Connell, M.G.; Edgar, D.H.; Kenny, S.E. Properties of secondary and tertiary human enteric nervous system neurospheres. J. Pediatr. Surg. 2009, 44, 1249–1255; discussion 1255–1256. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Lu, L.; Li, Y.; Hu, J. Neurospheres Induced from Human Adipose-Derived Stem Cells as a New Source of Neural Progenitor Cells. Cell Transplant. 2019, 28, 66S–75S. [Google Scholar] [CrossRef] [PubMed]
- Osterberg, N.; Roussa, E. Characterization of primary neurospheres generated from mouse ventral rostral hindbrain. Cell Tissue Res. 2009, 336, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Salzer, J.L. Polarized domains of myelinated axons. Neuron 2003, 40, 297–318. [Google Scholar] [CrossRef] [Green Version]
- Bonnon, C.; Goutebroze, L.; Denisenko-Nehrbass, N.; Girault, J.A.; Faivre-Sarrailh, C. The paranodal complex of F3/contactin and caspr/paranodin traffics to the cell surface via a non-conventional pathway. J. Biol. Chem. 2003, 278, 48339–48347. [Google Scholar] [CrossRef] [Green Version]
- Charles, P.; Tait, S.; Faivre-Sarrailh, C.; Barbin, G.; Gunn-Moore, F.; Denisenko-Nehrbass, N.; Guennoc, A.M.; Girault, J.A.; Brophy, P.J.; Lubetzki, C. Neurofascin is a glial receptor for the paranodin/Caspr-contactin axonal complex at the axoglial junction. Curr. Biol. 2002, 12, 217–220. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, D.; Zalfa, C.; Nodari, L.R.; Gelati, M.; Carlessi, L.; Delia, D.; Vescovi, A.L.; De Filippis, L. Differential pathotropism of non-immortalized and immortalized human neural stem cell lines in a focal demyelination model. Cell Mol. Life Sci. 2012, 69, 1193–1210. [Google Scholar] [CrossRef]
- Cocks, G.; Romanyuk, N.; Amemori, T.; Jendelova, P.; Forostyak, O.; Jeffries, A.R.; Perfect, L.; Thuret, S.; Dayanithi, G.; Sykova, E.; et al. Conditionally immortalized stem cell lines from human spinal cord retain regional identity and generate functional V2a interneurons and motorneurons. Stem Cell Res. Ther. 2013, 4, 69. [Google Scholar] [CrossRef] [Green Version]
- Ishii, T.; Kawakami, E.; Endo, K.; Misawa, H.; Watabe, K. Myelinating cocultures of rodent stem cell line-derived neurons and immortalized Schwann cells. Neuropathology 2017, 37, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Colucci-D’Amato, L.; Farina, A.; Vissers, J.P.; Chambery, A. Quantitative neuroproteomics: Classical and novel tools for studying neural differentiation and function. Stem Cell Rev. Rep. 2011, 7, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Severino, V.; Farina, A.; Colucci-D’Amato, L.; Reccia, M.G.; Volpicelli, F.; Parente, A.; Chambery, A. Secretome profiling of differentiated neural mes-c-myc A1 cell line endowed with stem cell properties. Biochim. Biophys. Acta 2013, 1834, 2385–2395. [Google Scholar] [CrossRef] [PubMed]
- Florio, E.; Keller, S.; Coretti, L.; Affinito, O.; Scala, G.; Errico, F.; Fico, A.; Boscia, F.; Sisalli, M.J.; Reccia, M.G.; et al. Tracking the evolution of epialleles during neural differentiation and brain development: D-Aspartate oxidase as a model gene. Epigenetics 2017, 12, 41–54. [Google Scholar] [CrossRef]
- Chambery, A.; Farina, A.; Di Maro, A.; Rossi, M.; Abbondanza, C.; Moncharmont, B.; Malorni, L.; Cacace, G.; Pocsfalvi, G.; Malorni, A.; et al. Proteomic analysis of MCF-7 cell lines expressing the zinc-finger or the proline-rich domain of retinoblastoma-interacting-zinc-finger protein. J. Proteome Res. 2006, 5, 1176–1185. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A.; Poulet, S.A.; Carotenuto, Y.; Buttino, I.; Romano, G.; Casotti, R.; Pohnert, G.; Wichard, T.; Colucci-D’Amato, L.; et al. Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 2004, 429, 403–407. [Google Scholar] [CrossRef]
- Di Lieto, A.; Leo, D.; Volpicelli, F.; di Porzio, U.; Colucci-D’Amato, L. FLUOXETINE modifies the expression of serotonergic markers in a differentiation-dependent fashion in the mesencephalic neural cell line A1 mes c-myc. Brain Res. 2007, 1143, 1–10. [Google Scholar] [CrossRef]
- Gentile, M.T.; Nawa, Y.; Lunardi, G.; Florio, T.; Matsui, H.; Colucci-D’Amato, L. Tryptophan hydroxylase 2 (TPH2) in a neuronal cell line: Modulation by cell differentiation and NRSF/rest activity. J. Neurochem. 2012, 123, 963–970. [Google Scholar] [CrossRef]
- Gentile, M.T.; Ciniglia, C.; Reccia, M.G.; Volpicelli, F.; Gatti, M.; Thellung, S.; Florio, T.; Melone, M.A.; Colucci-D’Amato, L. Ruta graveolens L. induces death of glioblastoma cells and neural progenitors, but not of neurons, via ERK 1/2 and AKT activation. PLoS ONE 2015, 10, e0118864. [Google Scholar] [CrossRef]
- Nizzari, M.; Barbieri, F.; Gentile, M.T.; Passarella, D.; Caorsi, C.; Diaspro, A.; Taglialatela, M.; Pagano, A.; Colucci-D’Amato, L.; Florio, T.; et al. Amyloid-β protein precursor regulates phosphorylation and cellular compartmentalization of microtubule associated protein tau. J. Alzheimers Dis. 2012, 29, 211–227. [Google Scholar] [CrossRef]
- Nawa, Y.; Kaneko, H.; Oda, M.; Tsubonoya, M.; Hiroi, T.; Gentile, M.T.; Colucci-D’Amato, L.; Takahashi, R.; Matsui, H. Functional characterization of the neuron-restrictive silencer element in the human tryptophan hydroxylase 2 gene expression. J. Neurochem. 2017, 142, 827–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favaro, R.; Valotta, M.; Ferri, A.L.; Latorre, E.; Mariani, J.; Giachino, C.; Lancini, C.; Tosetti, V.; Ottolenghi, S.; Taylor, V.; et al. Hippocampal development and neural stem cell maintenance require Sox2-dependent regulation of Shh. Nat. Neurosci. 2009, 12, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Ellis, P.; Fagan, B.M.; Magness, S.T.; Hutton, S.; Taranova, O.; Hayashi, S.; McMahon, A.; Rao, M.; Pevny, L. SOX2, a persistent marker for multipotential neural stem cells derived from embryonic stem cells, the embryo or the adult. Dev. Neurosci. 2004, 26, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Pevny, L.H.; Nicolis, S.K. Sox2 roles in neural stem cells. Int. J. Biochem. Cell Biol. 2010, 42, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Andrianakos, R.; Yang, Y.; Liu, C.; Lu, W. Kruppel-like factor 4 (Klf4) prevents embryonic stem (ES) cell differentiation by regulating Nanog gene expression. J. Biol. Chem. 2010, 285, 9180–9189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.; Guo, G.; Wray, J.; Eyres, I.; Nichols, J.; Grotewold, L.; Morfopoulou, S.; Humphreys, P.; Mansfield, W.; Walker, R.; et al. Oct4 and LIF/Stat3 additively induce Krüppel factors to sustain embryonic stem cell self-renewal. Cell Stem Cell 2009, 5, 597–609. [Google Scholar] [CrossRef]
- Jiang, J.; Chan, Y.S.; Loh, Y.H.; Cai, J.; Tong, G.Q.; Lim, C.A.; Robson, P.; Zhong, S.; Ng, H.H. A core Klf circuitry regulates self-renewal of embryonic stem cells. Nat. Cell Biol. 2008, 10, 353–360. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Liu, M.; Niu, W.; Zhang, C.L. Dysregulation of Kruppel-like factor 4 during brain development leads to hydrocephalus in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 21117–21121. [Google Scholar] [CrossRef] [Green Version]
- Caiazzo, M.; Colucci-D’Amato, L.; Esposito, M.T.; Parisi, S.; Stifani, S.; Ramirez, F.; di Porzio, U. Transcription factor KLF7 regulates differentiation of neuroectodermal and mesodermal cell lineages. Exp. Cell Res. 2010, 316, 2365–2376. [Google Scholar] [CrossRef]
- Caiazzo, M.; Colucci-D’Amato, L.; Volpicelli, F.; Speranza, L.; Petrone, C.; Pastore, L.; Stifani, S.; Ramirez, F.; Bellenchi, G.C.; di Porzio, U. Krüppel-like factor 7 is required for olfactory bulb dopaminergic neuron development. Exp. Cell Res. 2011, 317, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugas, J.C.; Ibrahim, A.; Barres, B.A. The T3-induced gene KLF9 regulates oligodendrocyte differentiation and myelin regeneration. Mol. Cell. Neurosci. 2012, 50, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laitman, B.M.; Asp, L.; Mariani, J.N.; Zhang, J.; Liu, J.; Sawai, S.; Chapouly, C.; Horng, S.; Kramer, E.G.; Mitiku, N.; et al. The Transcriptional Activator Krüppel-like Factor-6 Is Required for CNS Myelination. PLoS Biol. 2016, 14, e1002467. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.H.; Fukaya, M.; Yang, J.K.; Rothstein, J.D.; Bergles, D.E. NG2+ CNS glial progenitors remain committed to the oligodendrocyte lineage in postnatal life and following neurodegeneration. Neuron 2010, 68, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampton, D.W.; Rhodes, K.E.; Zhao, C.; Franklin, R.J.; Fawcett, J.W. The responses of oligodendrocyte precursor cells, astrocytes and microglia to a cortical stab injury, in the brain. Neuroscience 2004, 127, 813–820. [Google Scholar] [CrossRef]
- Magnus, T.; Carmen, J.; Deleon, J.; Xue, H.; Pardo, A.C.; Lepore, A.C.; Mattson, M.P.; Rao, M.S.; Maragakis, N.J. Adult glial precursor proliferation in mutant SOD1G93A mice. Glia 2008, 56, 200–208. [Google Scholar] [CrossRef]
- Sellers, D.L.; Maris, D.O.; Horner, P.J. Postinjury niches induce temporal shifts in progenitor fates to direct lesion repair after spinal cord injury. J. Neurosci. 2009, 29, 6722–6733. [Google Scholar] [CrossRef] [Green Version]
- Suda, H.; Hosokawa, T.; Ohno, R.; Hamaguchi, K.; Tsukada, Y. 2’,3’-Cyclic nucleotide 3’-phosphodiesterase activity in the cerebrospinal fluid of patients with demyelinating diseases. Neurochem. Pathol. 1984, 2, 85–102. [Google Scholar] [CrossRef]
- Kuhn, S.; Gritti, L.; Crooks, D.; Dombrowski, Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells 2019, 8, 1424. [Google Scholar] [CrossRef] [Green Version]
- Kerman, B.E.; Kim, H.J.; Padmanabhan, K.; Mei, A.; Georges, S.; Joens, M.S.; Fitzpatrick, J.A.; Jappelli, R.; Chandross, K.J.; August, P.; et al. In vitro myelin formation using embryonic stem cells. Development 2015, 142, 2213–2225. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Fex Svenningsen, A.; Shan, W.S.; Colman, D.R.; Pedraza, L. Rapid method for culturing embryonic neuron-glial cell cocultures. J. Neurosci. Res. 2003, 72, 565–573. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| und vs. diff | und-Ns vs. diff-Ns | diff Ns-A1 d3 | diff Ns-A1 d10 | diff Ns-A1 d13 | diff Ns-A1 d17 | diff Ns-A1 d24 | |||
|---|---|---|---|---|---|---|---|---|---|
| Klf4 | ++ | + | ++ | + | + | + | N.D. | + | N.D. |
| GalC | ++ | + | + | + | ++ | + | ++ | N.D. | N.D. |
| ßIII-tub | + | ++ | + | ++ | ++ | + | + | N.D. | N.D. |
| GFAP | ++ | + | ++ | + | - | - | N.D. | - | N.D. |
| Gene of Interest | Primers Sequence |
|---|---|
| Nestin | Forward 5′- AGCAACTGGCACACCTCAAGA- 3′ Reverse 5′- CTCAGCCTCCAGCAGAGTCC- 3′ |
| βIII-tubulin | Forward 5′- CGTGGGCTCAAAATGTCATC- 3′ Reverse 5’- TGGCTGTGAACTGCTCCGAGAT- 3’ |
| GFAP | Forward 5′- GAGGGACAACTTTGCACAGGA- 3′ Reverse 5′- CCAGCCTCAGGTTGGTTTCAT- 3′ |
| Sox2 | Forward 5′- AGGGCTGGACTGCGAACTG- 3′ Reverse 5′- TTTGCACCCCTCCCAATTC- 3′ |
| HPRT | Forward 5′- TGCCCTTGACTATAATGAGTACTTCAG- 3′ Reverse 5′- TTGGCTTTTCCAGTTTCACTAATG- 3′ |
| GalC | Forward 5′- GCCTTTCATTCCAAATCCCAA- 3′ Reverse 5′- AGCTCACTGCCTACTATGTTG- 3′ |
| Klf4 | Forward 5′- CCCTCCTTTCCTGCCAGACC- 3′ Reverse 5′- ACCTTCTTCCCCTCTTTGGCTTG- 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reccia, M.G.; Volpicelli, F.; Benedikz, E.; Svenningsen, Å.F.; Colucci-D’Amato, L. Generation of High-Yield, Functional Oligodendrocytes from a c-myc Immortalized Neural Cell Line, Endowed with Staminal Properties. Int. J. Mol. Sci. 2021, 22, 1124. https://doi.org/10.3390/ijms22031124
Reccia MG, Volpicelli F, Benedikz E, Svenningsen ÅF, Colucci-D’Amato L. Generation of High-Yield, Functional Oligodendrocytes from a c-myc Immortalized Neural Cell Line, Endowed with Staminal Properties. International Journal of Molecular Sciences. 2021; 22(3):1124. https://doi.org/10.3390/ijms22031124
Chicago/Turabian StyleReccia, Mafalda Giovanna, Floriana Volpicelli, Eirkiur Benedikz, Åsa Fex Svenningsen, and Luca Colucci-D’Amato. 2021. "Generation of High-Yield, Functional Oligodendrocytes from a c-myc Immortalized Neural Cell Line, Endowed with Staminal Properties" International Journal of Molecular Sciences 22, no. 3: 1124. https://doi.org/10.3390/ijms22031124
APA StyleReccia, M. G., Volpicelli, F., Benedikz, E., Svenningsen, Å. F., & Colucci-D’Amato, L. (2021). Generation of High-Yield, Functional Oligodendrocytes from a c-myc Immortalized Neural Cell Line, Endowed with Staminal Properties. International Journal of Molecular Sciences, 22(3), 1124. https://doi.org/10.3390/ijms22031124