
Cold Adaptation in Antarctic Notothenioids: Comparative Transcriptomics Reveals Novel Insights in the Peculiar Role of Gills and Highlights Signatures of Cobalamin Deficiency

 , ,
, ,  ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. De Novo Assembly of the C. hamatus Gills Transcriptome
2.2. An Insight into the Highly Specialized Transcriptional Profile of the Gills
2.3. Evidence of Cold Adaptation in the Transcriptomes of Cryonotothenioidea Gills
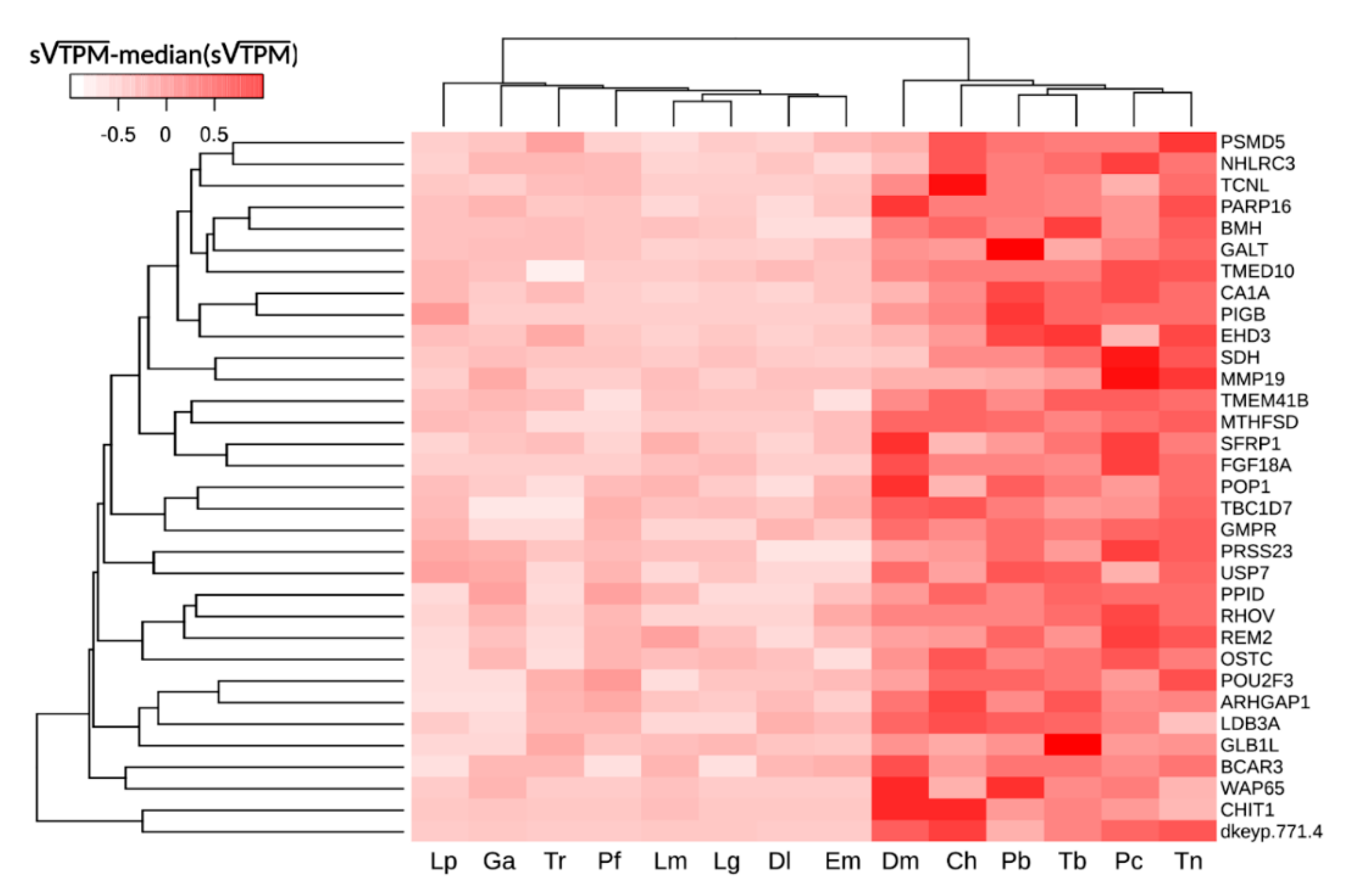
2.4. Comparative Transcriptome Analysis Reveals Signatures of Cobalamin Deficiency in Cryonotothenioidea
2.5. The High Expression of the Carbonic Anhydrase Genes CA1A and CA4A Explains the High Enzymatic Activity Observed in Antarctic Fish Gills
2.6. Constitutive Expression of Chaperones, Co-Chaperones, and the Proteasome Machinery
2.7. Other Remarks and Conclusions
3. Materials and Methods
3.1. Chionodraco Hamatus Transcriptome de Novo Assembly and Annotation
3.2. Assessment and Validation of Tissue-Wide Gene Expression Profile
3.3. Comparative Gene Expression Analysis
3.4. Phylogenetic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hüne, M.; González-Wevar, C.; Poulin, E.; Mansilla, A.; Fernández, D.A.; Barrera-Oro, E. Low Level of Genetic Divergence between Harpagifer Fish Species (Perciformes: Notothenioidei) Suggests a Quaternary Colonization of Patagonia from the Antarctic Peninsula. Polar Biol. 2015, 38, 607–617. [Google Scholar] [CrossRef]
- Coppes Petricorena, Z.L.; Somero, G.N. Biochemical Adaptations of Notothenioid Fishes: Comparisons between Cold Temperate South American and New Zealand Species and Antarctic Species. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 147, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Eastman, J.T.; Giordano, D.; Parisi, E.; Verde, C. Biogeography and Adaptation of Notothenioid Fish: Hemoglobin Function and Globin-Gene Evolution. Gene 2007, 398, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.L.; Halanych, K.M. Evaluating Connectivity in the Brooding Brittle Star Astrotoma Agassizii across the Drake Passage in the Southern Ocean. J. Hered. 2008, 99, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papetti, C.; Windisch, H.S.; La Mesa, M.; Lucassen, M.; Marshall, C.; Lamare, M.D. Non-Antarctic Notothenioids: Past Phylogenetic History and Contemporary Phylogeographic Implications in the Face of Environmental Changes. Mar. Genom. 2016, 25, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Near, T.J.; Dornburg, A.; Harrington, R.C.; Oliveira, C.; Pietsch, T.W.; Thacker, C.E.; Satoh, T.P.; Katayama, E.; Wainwright, P.C.; Eastman, J.T.; et al. Identification of the Notothenioid Sister Lineage Illuminates the Biogeographic History of an Antarctic Adaptive Radiation. BMC Evol. Biol. 2015, 15, 109. [Google Scholar] [CrossRef] [Green Version]
- Peck, L.S.; Webb, K.E.; Bailey, D.M. Extreme Sensitivity of Biological Function to Temperature in Antarctic Marine Species. Funct. Ecol. 2004, 18, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Eastman, J.T. The Nature of the Diversity of Antarctic Fishes. Polar Biol. 2005, 28, 93–107. [Google Scholar] [CrossRef]
- Beers, J.M.; Jayasundara, N. Antarctic Notothenioid Fish: What Are the Future Consequences of “losses” and “Gains” Acquired during Long-Term Evolution at Cold and Stable Temperatures? J. Exp. Biol. 2015, 218, 1834–1845. [Google Scholar] [CrossRef] [Green Version]
- Wells, R.M.G. Blood-Gas Transport and Hemoglobin Function in Polar Fishes: Does Low Temperature Explain Physiological Characters? In Fish Physiology; Academic Press: Cambridge, MA, USA, 2005; Volume 22, pp. 281–316. [Google Scholar]
- Ruud, J.T. Vertebrates without Erythrocytes and Blood Pigment. Nature 1954, 173, 848. [Google Scholar] [CrossRef]
- Near, T.J.; Parker, S.K.; Detrich, H.W. A Genomic Fossil Reveals Key Steps in Hemoglobin Loss by the Antarctic Icefishes. Mol. Biol. Evol. 2006, 23, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, F.; Pellegrino, D.; Amelio, D.; Tota, B. The Antarctic Hemoglobinless Icefish, Fifty Five Years Later: A Unique Cardiocirculatory Interplay of Disaptation and Phenotypic Plasticity. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 154, 10–28. [Google Scholar] [CrossRef]
- Beers, J.M.; Borley, K.A.; Sidell, B.D. Relationship among Circulating Hemoglobin, Nitric Oxide Synthase Activities and Angiogenic Poise in Red- and White-Blooded Antarctic Notothenioid Fishes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 156, 422–429. [Google Scholar] [CrossRef]
- O’Brien, K.M.; Mueller, I.A. The Unique Mitochondrial Form and Function of Antarctic Channichthyid Icefishes. Integr. Comp. Biol. 2010, 50, 993–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Near, T.J.; MacGuigan, D.J.; Parker, E.; Struthers, C.D.; Jones, C.D.; Dornburg, A. Phylogenetic Analysis of Antarctic Notothenioids Illuminates the Utility of RADseq for Resolving Cenozoic Adaptive Radiations. Mol. Phylogenet. Evol. 2018, 129, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-M.; Amores, A.; Kang, S.; Ahn, D.-H.; Kim, J.-H.; Kim, I.-C.; Lee, J.H.; Lee, S.G.; Lee, H.; Lee, J.; et al. Antarctic Blackfin Icefish Genome Reveals Adaptations to Extreme Environments. Nat. Ecol. Evol. 2019, 3, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Di Prisco, G.; Macdonald, J.A.; Brunori, M. Antarctic Fishes Survive Exposure to Carbon Monoxide. Experientia 1992, 48, 473–475. [Google Scholar] [CrossRef]
- Shin, S.C.; Ahn, D.H.; Kim, S.J.; Pyo, C.W.; Lee, H.; Kim, M.-K.; Lee, J.; Lee, J.E.; Detrich, H.W.; Postlethwait, J.H.; et al. The Genome Sequence of the Antarctic Bullhead Notothen Reveals Evolutionary Adaptations to a Cold Environment. Genome Biol. 2014, 15, 468. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.-H.; Shin, S.C.; Kim, B.-M.; Kang, S.; Kim, J.-H.; Ahn, I.; Park, J.; Park, H. Draft Genome of the Antarctic Dragonfish, Parachaenichthys Charcoti. GigaScience 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Bargelloni, L.; Babbucci, M.; Ferraresso, S.; Papetti, C.; Vitulo, N.; Carraro, R.; Pauletto, M.; Santovito, G.; Lucassen, M.; Mark, F.C.; et al. Draft Genome Assembly and Transcriptome Data of the Icefish Chionodraco Myersi Reveal the Key Role of Mitochondria for a Life without Hemoglobin at Subzero Temperatures. Commun. Biol. 2019, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bilyk, K.T.; Cheng, C.-H.C. Model of Gene Expression in Extreme Cold—Reference Transcriptome for the High-Antarctic Cryopelagic Notothenioid Fish Pagothenia Borchgrevinki. BMC Genom. 2013, 14, 634. [Google Scholar] [CrossRef] [Green Version]
- Gerdol, M.; Buonocore, F.; Scapigliati, G.; Pallavicini, A. Analysis and Characterization of the Head Kidney Transcriptome from the Antarctic Fish Trematomus Bernacchii (Teleostea, Notothenioidea): A Source for Immune Relevant Genes. Mar. Genom. 2015, 20, 13–15. [Google Scholar] [CrossRef]
- Coppe, A.; Agostini, C.; Marino, I.A.M.; Zane, L.; Bargelloni, L.; Bortoluzzi, S.; Patarnello, T. Genome Evolution in the Cold: Antarctic Icefish Muscle Transcriptome Reveals Selective Duplications Increasing Mitochondrial Function. Genome Biol. Evol. 2013, 5, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Cai, C.; Hu, X.; Liu, Y.; Guo, Y.; Hu, P.; Chen, Z.; Peng, S.; Zhang, D.; Jiang, S.; et al. Evolutionary Suppression of Erythropoiesis via the Modulation of TGF-β Signalling in an Antarctic Icefish. Mol. Ecol. 2015, 24, 4664–4678. [Google Scholar] [CrossRef]
- Song, W.; Li, L.; Huang, H.; Jiang, K.; Zhang, F.; Wang, L.; Zhao, M.; Ma, L. Tissue-Based Transcriptomics of Chionodraco Hamatus: Sequencing, de Novo Assembly, Annotation and Marker Discovery. J. Fish Biol. 2019, 94, 251–260. [Google Scholar] [CrossRef]
- Bilyk, K.T.; Cheng, C.-H.C. RNA-Seq Analyses of Cellular Responses to Elevated Body Temperature in the High Antarctic Cryopelagic Nototheniid Fish Pagothenia Borchgrevinki. Mar. Genom. 2014, 18, 163–171. [Google Scholar] [CrossRef]
- Huth, T.J.; Place, S.P. RNA-Seq Reveals a Diminished Acclimation Response to the Combined Effects of Ocean Acidification and Elevated Seawater Temperature in Pagothenia Borchgrevinki. Mar. Genom. 2016, 28, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkie, M.P. Mechanisms of Ammonia Excretion Across Fish Gills. Comp. Biochem. Physiol. A Physiol. 1997, 118, 39–50. [Google Scholar] [CrossRef]
- Evans, D.H. Teleost Fish Osmoregulation: What Have We Learned since August Krogh, Homer Smith, and Ancel Keys. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R704–R713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The Mucosal Immune System of Fish: The Evolution of Tolerating Commensals While Fighting Pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [Green Version]
- Hoar, W.S.; Randall, D.J. Gills Anatomy, Gas Transfer, and Acid-Base Regulation; Fish Physiology; Academic Press: Cambridge, MA, USA, 1984; Volume 10. [Google Scholar]
- Nuñez Ortiz, N.; Gerdol, M.; Stocchi, V.; Marozzi, C.; Randelli, E.; Bernini, C.; Buonocore, F.; Picchietti, S.; Papeschi, C.; Sood, N.; et al. T Cell Transcripts and T Cell Activities in the Gills of the Teleost Fish Sea Bass (Dicentrarchus Labrax). Dev. Comp. Immunol. 2014, 47, 309–318. [Google Scholar] [CrossRef]
- Greibe, E.; Fedosov, S.; Nexo, E. The Cobalamin-Binding Protein in Zebrafish Is an Intermediate between the Three Cobalamin-Binding Proteins in Human. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Marques, M.; Ruivo, R.; Delgado, I.; Wilson, J.M.; Aluru, N.; Castro, L.F.C. Basal Gnathostomes Provide Unique Insights into the Evolution of Vitamin B12 Binders. Genome Biol. Evol. 2014, 7, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuerges, J.; Garau, G.; Geremia, S.; Fedosov, S.N.; Petersen, T.E.; Randaccio, L. Structural Basis for Mammalian Vitamin B12 Transport by Transcobalamin. Proc. Natl. Acad. Sci. USA 2006, 103, 4386–4391. [Google Scholar] [CrossRef] [Green Version]
- Wuerges, J.; Geremia, S.; Randaccio, L. Structural Study on Ligand Specificity of Human Vitamin B12 Transporters. Biochem. J. 2007, 403, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-Specific Expression by Genome-Wide Integration of Transcriptomics and Antibody-Based Proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Hall, C.A. Transcobalamins I and II as Natural Transport Proteins of Vitamin B12. J. Clin. Invest. 1975, 56, 1125–1131. [Google Scholar] [CrossRef] [Green Version]
- Zdzitowiecki, K.; Cielecka, D. Digenea of Fishes of the Weddell Sea. II. The Genus Macvicaria (Opecoelidae). Acta Parasitol. 1997, 42, 77–83. [Google Scholar]
- Laskowski, Z.; Rocka, A.; Zdzitowiecki, K.; Ozouf-Costaz, C. Occurence of Endoparasitic Worms in Dusky Notothen, Trematomus Newnesi (Actinopterygii Nototheniidae), at Adélie Land, Antarctica. Pol. Polar Res. 2007, 28, 37–42. [Google Scholar]
- Coscia, M.R.; Oreste, U. Presence of Antibodies Specific for Proteins Of Contracaecum Osculatum (Rudolphi, 1908) in Plasma of Several Antarctic Teleosts. Fish Shellfish Immunol. 1998, 8, 295–302. [Google Scholar] [CrossRef]
- Coscia, M.R.; Oreste, U. Plasma and Bile Antibodies of the Teleost Trematomus Bernacchii Specific for the Nematode Pseudoterranova Decipiens. Dis. Aquat. Organ. 2000, 41, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Zdzitowiecki, K. New Data on the Occurrence of Fish Endoparasitic Worms off Adelie Land, Antarctica. Pol. Polar Res. 2001, 22, 159–165. [Google Scholar]
- Santoro, M.; Mattiucci, S.; Work, T.; Cimmaruta, R.; Nardi, V.; Cipriani, P.; Bellisario, B.; Nascetti, G. Parasitic Infection by Larval Helminths in Antarctic Fishes: Pathological Changes and Impact on the Host Body Condition Index. Dis. Aquat. Organ. 2013, 105, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, M.; Mattiucci, S.; Cipriani, P.; Bellisario, B.; Romanelli, F.; Cimmaruta, R.; Nascetti, G. Parasite Communities of Icefish (Chionodraco Hamatus) in the Ross Sea (Antarctica): Influence of the Host Sex on the Helminth Infracommunity Structure. PLoS ONE 2014, 9, e88876. [Google Scholar] [CrossRef]
- Weinstein, P.P.; Jaffe, J.J. Cobalamin and Folate Metabolism in Helminths. Blood Rev. 1987, 1, 245–253. [Google Scholar] [CrossRef]
- Nyberg, W. The Uptake and Distribution of Co60-Labeled Vitamin B12 by the Fish Tapeworm, Diphyllobothrium Latum. Exp. Parasitol. 1958, 7, 178–190. [Google Scholar] [CrossRef]
- Björkenheim, B. Optic Neuropathy Caused by Vitamin-B12 Deficiency in Carriers of the Fish Tapeworm, Diphyllobothrium Latum. Lancet 1966, 1, 688–690. [Google Scholar] [CrossRef]
- Vuylsteke, P.; Bertrand, C.; Verhoef, G.E.G.; Vandenberghe, P. Case of Megaloblastic Anemia Caused by Intestinal Taeniasis. Ann. Hematol. 2004, 83, 487–488. [Google Scholar] [CrossRef] [PubMed]
- Caterino, M.; Pastore, A.; Strozziero, M.G.; Di Giovamberardino, G.; Imperlini, E.; Scolamiero, E.; Ingenito, L.; Boenzi, S.; Ceravolo, F.; Martinelli, D.; et al. The Proteome of CblC Defect: In Vivo Elucidation of Altered Cellular Pathways in Humans. J. Inherit. Metab. Dis. 2015, 38, 969–979. [Google Scholar] [CrossRef]
- Kuklina, M.M.; Kuklin, V.V. Hematology and Blood Chemistry of the Herring Gull Laurus Argentatus Infested by Cestodes Diphyllobothrium dentriticum (Cestoda: Diphyllobothriiidae). Parazitologiia 2016, 50, 365–375. [Google Scholar]
- Miller, J.W.; Nadeau, M.R.; Smith, D.; Selhub, J. Vitamin B-6 Deficiency vs Folate Deficiency: Comparison of Responses to Methionine Loading in Rats. Am. J. Clin. Nutr. 1994, 59, 1033–1039. [Google Scholar] [CrossRef]
- Jakubowski, H. The Pathophysiological Hypothesis of Homocysteine Thiolactone-Mediated Vascular Disease. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2008, 59, 155–167. [Google Scholar]
- Zimny, J.; Sikora, M.; Guranowski, A.; Jakubowski, H. Protective Mechanisms against Homocysteine Toxicity: The Role of Bleomycin Hydrolase. J. Biol. Chem. 2006, 281, 22485–22492. [Google Scholar] [CrossRef] [Green Version]
- Field, M.S.; Anderson, D.D.; Stover, P.J. Mthfs Is an Essential Gene in Mice and a Component of the Purinosome. Front. Genet. 2011, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Kang, J.; Zhang, D. Microbial Production of Vitamin B12: A Review and Future Perspectives. Microb. Cell Fact. 2017, 16. [Google Scholar] [CrossRef] [Green Version]
- Santovito, G.; Marino, S.M.; Sattin, G.; Cappellini, R.; Bubacco, L.; Beltramini, M. Cloning and Characterization of Cytoplasmic Carbonic Anhydrase from Gills of Four Antarctic Fish: Insights into the Evolution of Fish Carbonic Anhydrase and Cold Adaptation. Polar Biol. 2012, 35, 1587–1600. [Google Scholar] [CrossRef]
- Maffia, M.; Rizzello, A.; Acierno, R.; Rollo, M.; Chiloiro, R.; Storelli, C. Carbonic Anhydrase Activity in Tissues of the Icefish Chionodraco Hamatus and of the Red-Blooded Teleosts Trematomus Bernacchii and Anguilla Anguilla. J. Exp. Biol. 2001, 204, 3983–3992. [Google Scholar]
- Marino, S.; Hayakawa, K.; Hatada, K.; Benfatto, M.; Rizzello, A.; Maffia, M.; Bubacco, L. Structural Features That Govern Enzymatic Activity in Carbonic Anhydrase from a Low-Temperature Adapted Fish, Chionodraco Hamatus. Biophys. J. 2007, 93, 2781–2790. [Google Scholar] [CrossRef] [Green Version]
- Gilmour, K.M. New Insights into the Many Functions of Carbonic Anhydrase in Fish Gills. Respir. Physiol. Neurobiol. 2012, 184, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Egginton, S.; Taylor, E.W.; Wilson, R.W.; Johnston, I.A.; Moon, T.W. Stress Response in the Antarctic Teleosts (Notothenia Neglecta Nybelin and N. Rossii Richardson). J. Fish Biol. 1991, 38, 225–235. [Google Scholar] [CrossRef]
- Harter, T.S.; Sackville, M.A.; Wilson, J.M.; Metzger, D.C.H.; Egginton, S.; Esbaugh, A.J.; Farrell, A.P.; Brauner, C.J. A Solution to Nature’s Haemoglobin Knockout: A Plasma-Accessible Carbonic Anhydrase Catalyses CO2 Excretion in Antarctic Icefish Gills. J. Exp. Biol. 2018, 221. [Google Scholar] [CrossRef] [Green Version]
- Tufts, B.L.; Gervais, M.R.; Staebler, M.; Weaver, J. Subcellular Distribution and Characterization of Gill Carbonic Anhydrase and Evidence for a Plasma Carbonic Anhydrase Inhibitor in Antarctic Fish. J. Comp. Physiol. B 2002, 172, 287–295. [Google Scholar] [CrossRef]
- Cuellar, J.; Yébenes, H.; Parker, S.K.; Carranza, G.; Serna, M.; Valpuesta, J.M.; Zabala, J.C.; Detrich, H.W. Assisted Protein Folding at Low Temperature: Evolutionary Adaptation of the Antarctic Fish Chaperonin CCT and Its Client Proteins. Biol. Open 2014, 3, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, G.E.; Buckley, B.A.; Airaksinen, S.; Keen, J.E.; Somero, G.N. Heat-Shock Protein Expression Is Absent in the Antarctic Fish Trematomus Bernacchii (Family Nototheniidae). J. Exp. Biol. 2000, 203, 2331–2339. [Google Scholar] [PubMed]
- Place, S.P.; Zippay, M.L.; Hofmann, G.E. Constitutive Roles for Inducible Genes: Evidence for the Alteration in Expression of the Inducible Hsp70 Gene in Antarctic Notothenioid Fishes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R429–R436. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Cheng, C.-H.C.; Zhang, J.; Cao, L.; Chen, L.; Zhou, L.; Jin, Y.; Ye, H.; Deng, C.; Dai, Z.; et al. Transcriptomic and Genomic Evolution under Constant Cold in Antarctic Notothenioid Fish. Proc. Natl. Acad. Sci. USA 2008, 105, 12944–12949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönbrunner, E.R.; Schmid, F.X. Peptidyl-Prolyl Cis-Trans Isomerase Improves the Efficiency of Protein Disulfide Isomerase as a Catalyst of Protein Folding. Proc. Natl. Acad. Sci. USA 1992, 89, 4510–4513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, K.; Yamashita, M.; Watabe, S.; Aida, K. The Warm Temperature Acclimation-Related 65-KDa Protein, Wap65, in Goldfish and Its Gene Expression. J. Biol. Chem. 1995, 270, 17087–17092. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.Y.; An, K.W.; Choi, Y.K.; Jo, P.G.; Min, B.H. Expression of Warm Temperature Acclimation-Related Protein 65-KDa (Wap65) MRNA, and Physiological Changes with Increasing Water Temperature in Black Porgy, Acanthopagrus Schlegeli. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2008, 309A, 206–214. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K.; Kern, R.; Mistou, M.-Y.; Dandekar, A.M.; Uratsu, S.L.; Richarme, G. The Chemical Chaperone Proline Relieves the Thermosensitivity of a DnaK Deletion Mutant at 42 Degrees C. J. Bacteriol. 2004, 186, 8149–8152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Ng, D.T.W. Glycosylation-Directed Quality Control of Protein Folding. Nat. Rev. Mol. Cell Biol. 2015, 16, 742. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, N.G.; Surolia, A. Role of Glycosylation in Nucleating Protein Folding and Stability. Biochem. J. 2017, 474, 2333–2347. [Google Scholar] [CrossRef] [PubMed]
- Peck, L.S. A Cold Limit to Adaptation in the Sea. Trends Ecol. Evol. 2016, 31, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Gogliettino, M.; Balestrieri, M.; Riccio, A.; Facchiano, A.; Fusco, C.; Palazzo, V.C.; Rossi, M.; Cocca, E.; Palmieri, G. Uncommon Functional Properties of the First Piscine 26S Proteasome from the Antarctic Notothenioid Trematomus Bernacchii. Biosci. Rep. 2016, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.M.; Grove, T.J.; O’Brien, K.M. Energetic Costs of Protein Synthesis Do Not Differ between Red- and White-Blooded Antarctic Notothenioid Fishes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 187, 177–183. [Google Scholar] [CrossRef]
- Zhang, Y.-A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a Primitive Immunoglobulin Class Specialized in Mucosal Immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef]
- Ametrano, A.; Gerdol, M.; Vitale, M.; Greco, S.; Oreste, U.; Coscia, M.R. Fitting Together the Evolutionary Puzzle Pieces of the Immunoglobulin T Gene from Antarctic Fishes. Preprints 2020. [Google Scholar] [CrossRef]
- Biscotti, M.A.; Gerdol, M.; Canapa, A.; Forconi, M.; Olmo, E.; Pallavicini, A.; Barucca, M.; Schartl, M. The Lungfish Transcriptome: A Glimpse into Molecular Evolution Events at the Transition from Water to Land. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Carniel, F.C.; Gerdol, M.; Montagner, A.; Banchi, E.; De Moro, G.; Manfrin, C.; Muggia, L.; Pallavicini, A.; Tretiach, M. New Features of Desiccation Tolerance in the Lichen Photobiont Trebouxia Gelatinosa Are Revealed by a Transcriptomic Approach. Plant Mol. Biol. 2016, 91, 319–339. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness with Single-Copy Orthologs. Bioinform. Oxf. Engl. 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Infante, C.; Matsuoka, M.P.; Asensio, E.; Cañavate, J.P.; Reith, M.; Manchado, M. Selection of Housekeeping Genes for Gene Expression Studies in Larvae from Flatfish Using Real-Time PCR. BMC Mol. Biol. 2008, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Øvergård, A.-C.; Nerland, A.H.; Patel, S. Evaluation of Potential Reference Genes for Real Time RT-PCR Studies in Atlantic Halibut (Hippoglossus Hippoglossus L.); during Development, in Tissues of Healthy and NNV-Injected Fish, and in Anterior Kidney Leucocytes. BMC Mol. Biol. 2010, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Musser, J.M.; Wagner, G.P. Character Trees from Transcriptome Data: Origin and Individuation of Morphological Characters and the so-Called “Species Signal”. J. Exp. Zoolog. B Mol. Dev. Evol. 2015, 324, 588–604. [Google Scholar] [CrossRef] [PubMed]
- Pankey, M.S.; Minin, V.N.; Imholte, G.C.; Suchard, M.A.; Oakley, T.H. Predictable Transcriptome Evolution in the Convergent and Complex Bioluminescent Organs of Squid. Proc. Natl. Acad. Sci. USA 2014, 111, E4736–E4742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the Model of DNA Substitution. Bioinform. Oxf. Engl. 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinform. Oxf. Engl. 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Description | CS | Qval | FC | Putative Function |
|---|---|---|---|---|---|
| TCNL | transcobalamin-like protein | 48 | 1.81 × 10−21 | 31.20 | cobalamin carrier protein |
| dkeyp-77h1.4 | si:dkeyp-77h1.4 | 48 | 1.02 × 10−20 | 494.00 | unknown |
| GMPR | guanosine monophosphate reductase | 48 | 5.76 × 10−19 | 9.96 | purine biosynthesis |
| MTHFSD | methenyltetrahydrofolate synthetase domain containing | 48 | 1.07 × 10−15 | 11.34 | tetrahydrofolate metabolism regulator |
| CHIT1 | chitotriosidase-I | 48 | 3.45 × 10−14 | 107.97 | defense against nematodes |
| FGF18A | fibroblast growth factor 18a | 48 | 1.85 × 10−10 | 46.61 | cell proliferation regulator |
| OSTC | oligosaccharyltransferase complex subunit | 48 | 1.36 × 10−8 | 5.38 | N-glycosylation of nascent polypeptides |
| BLMH | bleomycin hydrolase | 48 | 2.18 × 10−7 | 4.52 | homocysteine thiolactone detoxyfication |
| TMEM41B | transmembrane protein 41B | 48 | 1.46 × 10−5 | 6.03 | autophagosome formation |
| RHOV | ras homolog family member V | 47 | 1.43 × 10−13 | 7.20 | actin cytoskeleton control |
| PIGB | phosphatidylinositol glycan anchor biosynthesis, class B | 46 | 3.95 × 10−43 | 30.20 | GPI-anchor biosynthesis. |
| CA1A | carbonic anhydrase 1a | 46 | 2.45 × 10−5 | 8.71 | CO2 excretion, cellular pH regulation |
| WAP65 | Warm-temperature acclimation related 65 kDa protein | 46 | 2.75 × 10−5 | 22.85 | stress response gene |
| ARHGAP1 | Rho GTPase activating protein 1 | 45 | 2.49 × 10−10 | 7.37 | GTPase activator |
| LDB3 | LIM domain binding 3 | 44 | 9.36 × 10−15 | 8.38 | scaffolding protein |
| SFRP1 | frizzled-related protein | 44 | 1.47 × 10−8 | 9.20 | Wnt signaling modulator |
| MMP19 | matrix metallopeptidase 19 | 44 | 8.42 × 10−8 | 6.05 | extracellular matrix remodeling |
| NHLRC3 | NHL repeat containing 3 | 44 | 5.65 × 10−7 | 3.83 | unknown |
| GALT | galactose-1-phosphate uridylyltransferase | 44 | 2.81 × 10−3 | 4.44 | galactose catabolism |
| PARP16 | poly (ADP-ribose) polymerase family, member 16 | 44 | 2.83 × 10−2 | 3.56 | unfolded protein response |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansaloni, F.; Gerdol, M.; Torboli, V.; Fornaini, N.R.; Greco, S.; Giulianini, P.G.; Coscia, M.R.; Miccoli, A.; Santovito, G.; Buonocore, F.; et al. Cold Adaptation in Antarctic Notothenioids: Comparative Transcriptomics Reveals Novel Insights in the Peculiar Role of Gills and Highlights Signatures of Cobalamin Deficiency. Int. J. Mol. Sci. 2021, 22, 1812. https://doi.org/10.3390/ijms22041812
Ansaloni F, Gerdol M, Torboli V, Fornaini NR, Greco S, Giulianini PG, Coscia MR, Miccoli A, Santovito G, Buonocore F, et al. Cold Adaptation in Antarctic Notothenioids: Comparative Transcriptomics Reveals Novel Insights in the Peculiar Role of Gills and Highlights Signatures of Cobalamin Deficiency. International Journal of Molecular Sciences. 2021; 22(4):1812. https://doi.org/10.3390/ijms22041812
Chicago/Turabian StyleAnsaloni, Federico, Marco Gerdol, Valentina Torboli, Nicola Reinaldo Fornaini, Samuele Greco, Piero Giulio Giulianini, Maria Rosaria Coscia, Andrea Miccoli, Gianfranco Santovito, Francesco Buonocore, and et al. 2021. "Cold Adaptation in Antarctic Notothenioids: Comparative Transcriptomics Reveals Novel Insights in the Peculiar Role of Gills and Highlights Signatures of Cobalamin Deficiency" International Journal of Molecular Sciences 22, no. 4: 1812. https://doi.org/10.3390/ijms22041812