B Cells with a Senescent-Associated Secretory Phenotype Accumulate in the Adipose Tissue of Individuals with Obesity

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
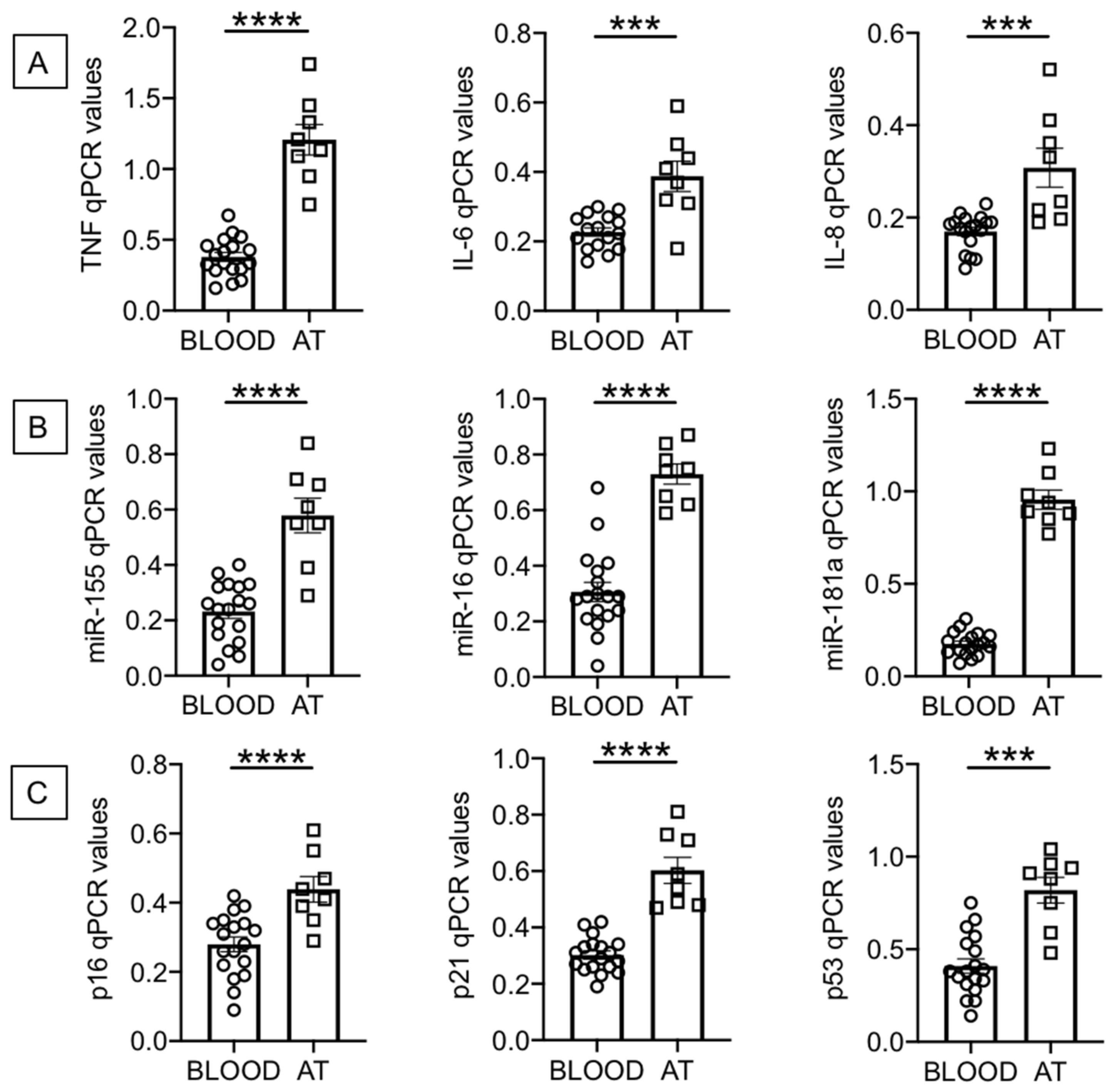
2.1. Increased Expression of SASP Markers in B Cells from the AT of Individuals with Obesity as Compared to the Peripheral Blood
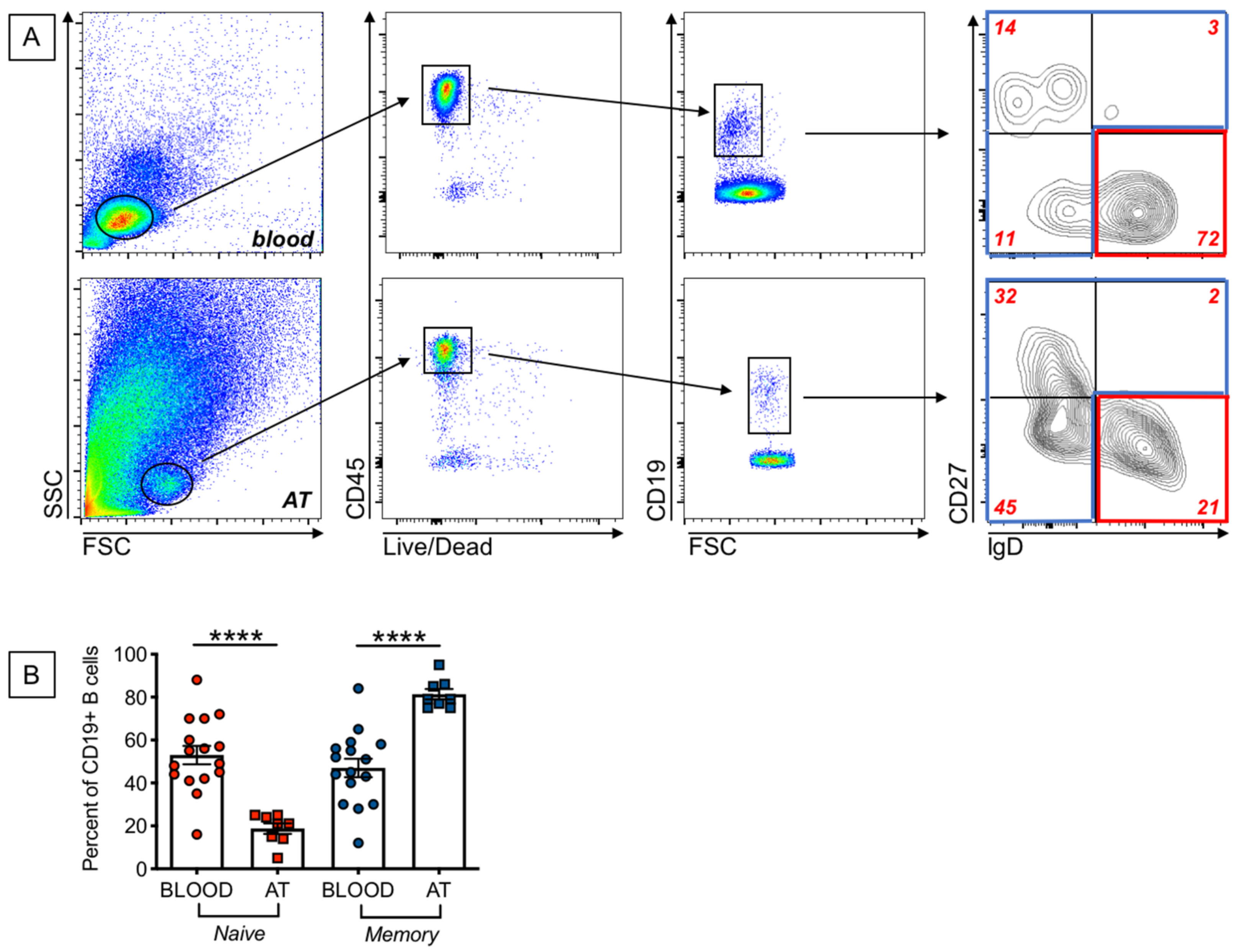
2.2. Increased Frequencies of Memory B Cells, and Decreased Frequencies of Naïve B Cells, in the AT of Individuals with Obesity as Compared to the Peripheral Blood
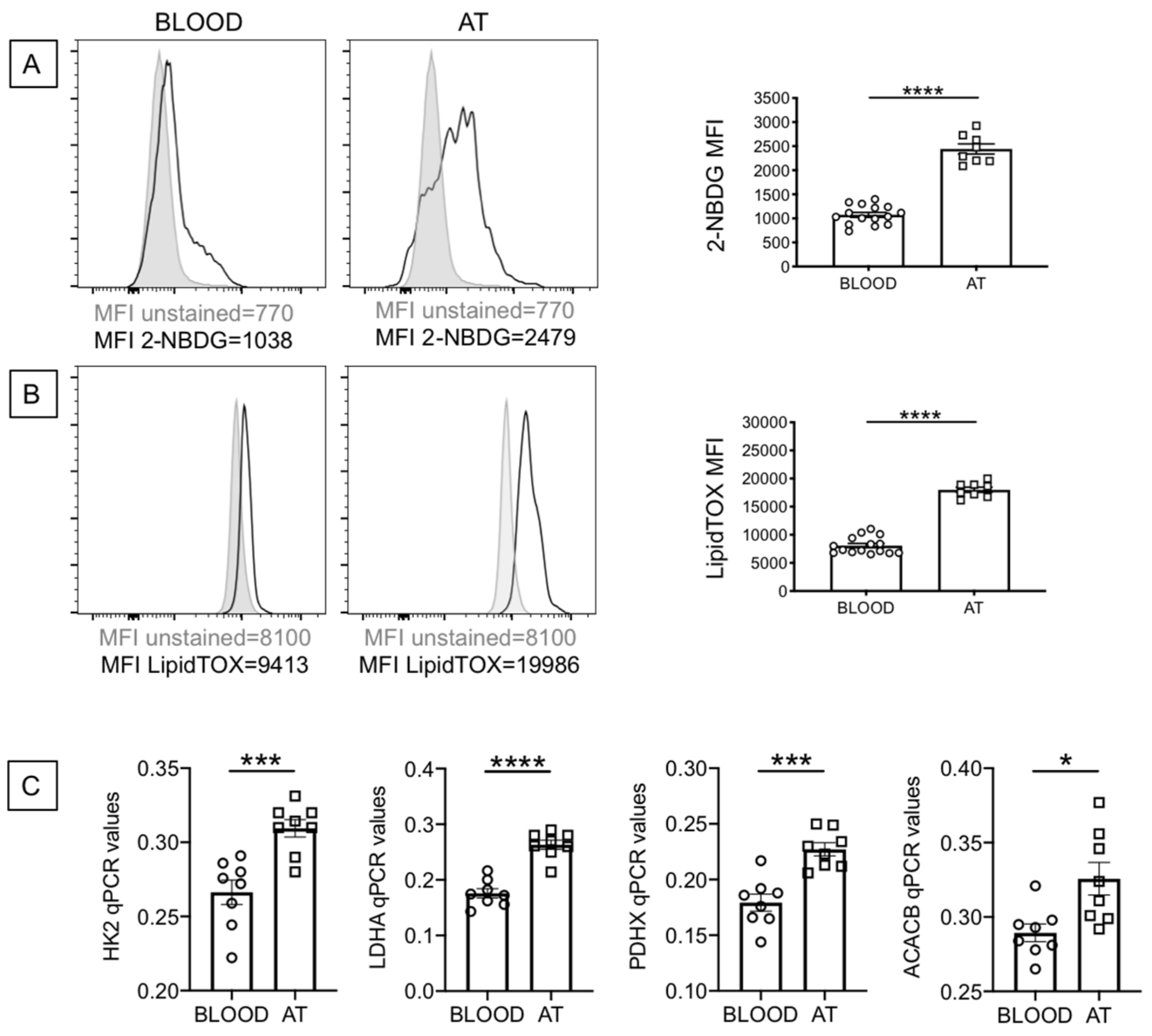
2.3. B Cells from the AT of Individuals with Obesity Are Highly Metabolic as Compared to Those from the Peripheral Blood
3. Discussion and Conclusions
4. Materials and Methods
4.1. Subjects
4.2. Isolation and Processing of the AT
4.3. Flow Cytometry
4.4. B Cell Sorting
4.5. RNA Extraction and Quantitative (q)PCR
4.6. Glucose and Lipid Uptake Measurements
4.7. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Childs, B.G.; Durik, M.; Baker, D.J.; Van Deursen, J.M. Cellular Senescence in Aging and Age-Related Disease: From Mechanisms to Therapy. Nat. Med. 2015, 21, 1424–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Deursen, J.M. The Role of Senescent Cells in Ageing. Nat. Cell Biol. 2014, 509, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Campisi, J. Cellular Senescence: Putting the Paradoxes in Perspective. Curr. Opin. Genet. Dev. 2011, 21, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikora, E.; Arendt, T.; Bennett, M.; Narita, M. Impact of Cellular Senescence Signature on Ageing Research. Ageing Res. Rev. 2011, 10, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J. Cellular Senescence and Apoptosis: How Cellular Responses Might Influence Aging Phenotypes. Exp. Gerontol. 2003, 38, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Passos, J.F.; Nelson, G.; Wang, C.; Richter, T.; Simillion, C.; Proctor, C.J.; Miwa, S.; Olijslagers, S.; Hallinan, J.; Wipat, A.; et al. Feedback between p21 and Reactive Oxygen Production Is Necessary for Cell Senescence. Mol. Syst. Biol. 2010, 6, 347. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Blomberg, B.B. Adipose Tissue, Immune Aging, and Cellular Senescence. Semin. Immunopathol. 2020, 42, 573–587. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, K.K.; Jiang, X.; Xu, A.; Cheng, K.K. The Role of Adipose Tissue Senescence in Obesity- and Ageing-Related Metabolic Disorders. Clin. Sci. 2020, 134, 315–330. [Google Scholar] [CrossRef]
- Tchkonia, T.; Morbeck, D.E.; Von Zglinicki, T.; Van Deursen, J.; Lustgarten, J.; Scrable, H.; Khosla, S.; Jensen, M.D.; Kirkland, J.L. Fat Tissue, Aging, and Cellular Senescence. Aging Cell 2010, 9, 667–684. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflammaging: An Evolutionary Perspective on Immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and Metabolic Disorders. Nat. Cell Biol. 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Johnson, A.M.; Olefsky, J.M. The Origins and Drivers of Insulin Resistance. Cell 2013, 152, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and Insulin Resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Apovian, C.M.; Gokce, N. Obesity and Cardiovascular Disease. Circulation 2012, 125, 1178–1182. [Google Scholar] [CrossRef] [Green Version]
- Renehan, A.G.; Tyson, M.; Egger, M.; Heller, R.F.; Zwahlen, M. Body-Mass Index and Incidence of Cancer: A Systematic Review and Meta-Analysis of Prospective Observational Studies. Lancet 2008, 371, 569–578. [Google Scholar] [CrossRef]
- Casas, R.; Sacanella, E.; Estruch, R. The Immune Protective Effect of the Mediterranean Diet against Chronic Low-Grade Inflammatory Diseases. Endocrine Metab. Immune Disord. Drug Targets 2014, 14, 245–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hass, D.J.; Brensinger, C.M.; Lewis, J.D.; Lichtenstein, G.R. The Impact of Increased Body Mass Index on the Clinical Course of Crohn’s Disease. Clin. Gastroenterol. Hepatol. 2006, 4, 482–488. [Google Scholar] [CrossRef]
- Frasca, D.; Ferracci, F.; Diaz, A.; Romero, M.; Lechner, S.; Blomberg, B.B. Obesity Decreases B Cell Responses in Young and Elderly Individuals. Obesity 2016, 24, 615–625. [Google Scholar] [CrossRef]
- Ovsyannikova, I.G.; White, S.J.; Larrabee, B.R.; Grill, D.E.; Jacobson, R.M.; Poland, G.A. Leptin and Leptin-Related Gene Polymorphisms, Obesity, and Influenza A/H1N1 Vaccine-Induced Immune Responses in Older Individuals. Vaccine 2014, 32, 881–887. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, P.A.; Paich, H.A.; Handy, J.; Karlsson, E.A.; Hudgens, M.G.; Sammon, A.B.; Holland, L.A.; Weir, S.; Noah, T.L.; Beck, M.A. Obesity Is Associated with Impaired Immune Response to Influenza Vaccination in Humans. Int. J. Obes. 2011, 36, 1072–1077. [Google Scholar] [CrossRef] [Green Version]
- George, M.; Baker, J.F. The Obesity Epidemic and Consequences for Rheumatoid Arthritis Care. Curr. Rheumatol. Rep. 2016, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ottaviani, S.; Allanore, Y.; Tubach, F.; Forien, M.; Gardette, A.; Pasquet, B.; Palazzo, E.; Meunier, M.; Hayem, G.; Job-Deslandre, C.; et al. Body Mass Index Influences the Response to Infliximab in Ankylosing Spondylitis. Arthritis Res. Ther. 2012, 14, R115. [Google Scholar] [CrossRef] [Green Version]
- Sandberg, M.E.C.; Bengtsson, C.; Källberg, H.; Wesley, A.; Klareskog, L.; Alfredsson, L.; Saevarsdottir, S. Overweight Decreases the Chance of Achieving Good Response and Low Disease Activity in Early Rheumatoid Arthritis. Ann. Rheum. Dis. 2014, 73, 2029–2033. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, M.; Kinoshita, K.; Fagarasan, S.; Yamada, S.; Shinkai, Y.; Honjo, T. Class Switch Recombination and Hypermutation Require Activation-Induced Cytidine Deaminase (AID), a Potential RNA Editing Enzyme. Cell 2000, 102, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Diaz, A.; Romero, M.; Landin, A.M.; Phillips, M.; Lechner, S.C.; Ryan, J.G.; Blomberg, B.B. Intrinsic Defects in B Cell Response to Seasonal Influenza Vaccination in Elderly Humans. Vaccine 2010, 28, 8077–8084. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Diaz, A.; Romero, M.; Phillips, M.; Mendez, N.V.; Landin, A.M.; Blomberg, B.B. Unique Biomarkers for B-Cell Function Predict the Serum Response to Pandemic H1N1 Influenza Vaccine. Int. Immunol. 2012, 24, 175–182. [Google Scholar] [CrossRef]
- Khurana, S.; Frasca, D.; Blomberg, B.; Golding, H. AID Activity in B Cells Strongly Correlates with Polyclonal Antibody Affinity Maturation In-Vivo Following Pandemic 2009-H1N1 Vaccination in Humans. PLOS Pathog. 2012, 8, e1002920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-H.; Kim, S.R.; Han, D.H.; Yu, H.T.; Han, Y.D.; Kim, J.H.; Kim, S.H.; Lee, C.J.; Min, B.-H.; Kim, D.-H.; et al. Senescent T Cells Predict the Development of Hyperglycemia in Humans. Diabetes 2019, 68, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Diaz, A.; Romero, M.; Garcia, D.; Jayram, D.; Thaller, S.; Piqueras, M.D.C.; Bhattacharya, S.; Blomberg, B.B. Identification and Characterization of Adipose Tissue-Derived Human Antibodies With “Anti-Self” Specificity. Front. Immunol. 2020, 11, 392. [Google Scholar] [CrossRef] [Green Version]
- Frasca, D.; Diaz, A.; Romero, M.; Thaller, S.; Blomberg, B.B. Secretion of Autoimmune Antibodies in the Human Subcutaneous Adipose Tissue. PLoS ONE 2018, 13, e0197472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, C.D.; Campisi, J. From Ancient Pathways to Aging Cells—Connecting Metabolism and Cellular Senescence. Cell Metab. 2016, 23, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Hamon, M.; Ahmed, E.K.; Baraibar, M.A.; Friguet, B. Proteome Oxidative Modifications and Impairment of Specific Metabolic Pathways During Cellular Senescence and Aging. Proteomics 2019, 20, e1800421. [Google Scholar] [CrossRef] [PubMed]
- Henneken, M.; Dörner, T.; Burmester, G.-R.; Berek, C. Differential Expression of Chemokine Receptors on Peripheral Blood B Cells from Patients with Rheumatoid Arthritis and Systemic Lupus Erythematosus. Arthritis Res. Ther. 2005, 7, R1001–R1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, P.; Kulyté, A. MicroRNA Regulatory Networks in Human Adipose Tissue and Obesity. Nat. Rev. Endocrinol. 2015, 11, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Deiuliis, A.J. MicroRNAs as Regulators of Metabolic Disease: Pathophysiologic Significance and Emerging Role as Biomarkers and Therapeutics. Int. J. Obes. 2016, 40, 88–101. [Google Scholar] [CrossRef] [Green Version]
- Iacomino, G.; Siani, A. Role of MicroRNAs in Obesity and Obesity-Related Diseases. Genes Nutr. 2017, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Diaz, A.; Romero, M.; Blomberg, B.B. Human Peripheral Late/Exhausted Memory B Cells Express a Senescent-Associated Secretory Phenotype and Preferentially Utilize Metabolic Signaling Pathways. Exp. Gerontol. 2017, 87, 113–120. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity and Its Impact on Metabolic Syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Passos, J.F.; Simillion, C.; Hallinan, J.; Wipat, A.; Von Zglinicki, T. Cellular Senescence: Unravelling Complexity. AGE 2009, 31, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Valdes, A.M.; Andrew, T.; Gardner, J.P.; Kimura, M.; Oelsner, E.; Cherkas, L.F.; Aviv, A.; Spector, T.D. Obesity, Cigarette Smoking, and Telomere Length in Women. Lancet 2005, 366, 662–664. [Google Scholar] [CrossRef]
- Camell, C.D.; Günther, P.; Lee, A.; Goldberg, E.L.; Spadaro, O.; Youm, Y.-H.; Bartke, A.; Hubbard, G.B.; Ikeno, Y.; Ruddle, N.H.; et al. Aging Induces an Nlrp3 Inflammasome-Dependent Expansion of Adipose B Cells That Impairs Metabolic Homeostasis. Cell Metab. 2019, 30, 1024–1039.e6. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Diaz, A.; Romero, M.; Vazquez, T.; Blomberg, B.B. Obesity Induces Pro-inflammatory B Cells and Impairs B Cell Function in Old Mice. Mech. Ageing Dev. 2017, 162, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winer, D.A.; Winer, S.; Shen, L.; Wadia, P.P.; Yantha, J.; Paltser, G.; Tsui, H.; Wu, P.; Davidson, M.G.; Alonso, M.N.; et al. B Cells Promote Insulin Resistance through Modulation of T Cells and Production of Pathogenic Igg Antibodies. Nat. Med. 2011, 17, 610–617. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frasca, D.; Romero, M.; Diaz, A.; Garcia, D.; Thaller, S.; Blomberg, B.B. B Cells with a Senescent-Associated Secretory Phenotype Accumulate in the Adipose Tissue of Individuals with Obesity. Int. J. Mol. Sci. 2021, 22, 1839. https://doi.org/10.3390/ijms22041839
Frasca D, Romero M, Diaz A, Garcia D, Thaller S, Blomberg BB. B Cells with a Senescent-Associated Secretory Phenotype Accumulate in the Adipose Tissue of Individuals with Obesity. International Journal of Molecular Sciences. 2021; 22(4):1839. https://doi.org/10.3390/ijms22041839
Chicago/Turabian StyleFrasca, Daniela, Maria Romero, Alain Diaz, Denisse Garcia, Seth Thaller, and Bonnie B. Blomberg. 2021. "B Cells with a Senescent-Associated Secretory Phenotype Accumulate in the Adipose Tissue of Individuals with Obesity" International Journal of Molecular Sciences 22, no. 4: 1839. https://doi.org/10.3390/ijms22041839
APA StyleFrasca, D., Romero, M., Diaz, A., Garcia, D., Thaller, S., & Blomberg, B. B. (2021). B Cells with a Senescent-Associated Secretory Phenotype Accumulate in the Adipose Tissue of Individuals with Obesity. International Journal of Molecular Sciences, 22(4), 1839. https://doi.org/10.3390/ijms22041839