
Some New Methodological and Conceptual Aspects of the “Acid Growth Theory” for the Auxin Action in Maize (Zea mays L.) Coleoptile Segments: Do Acid- and Auxin-Induced Rapid Growth Differ in Their Mechanisms?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
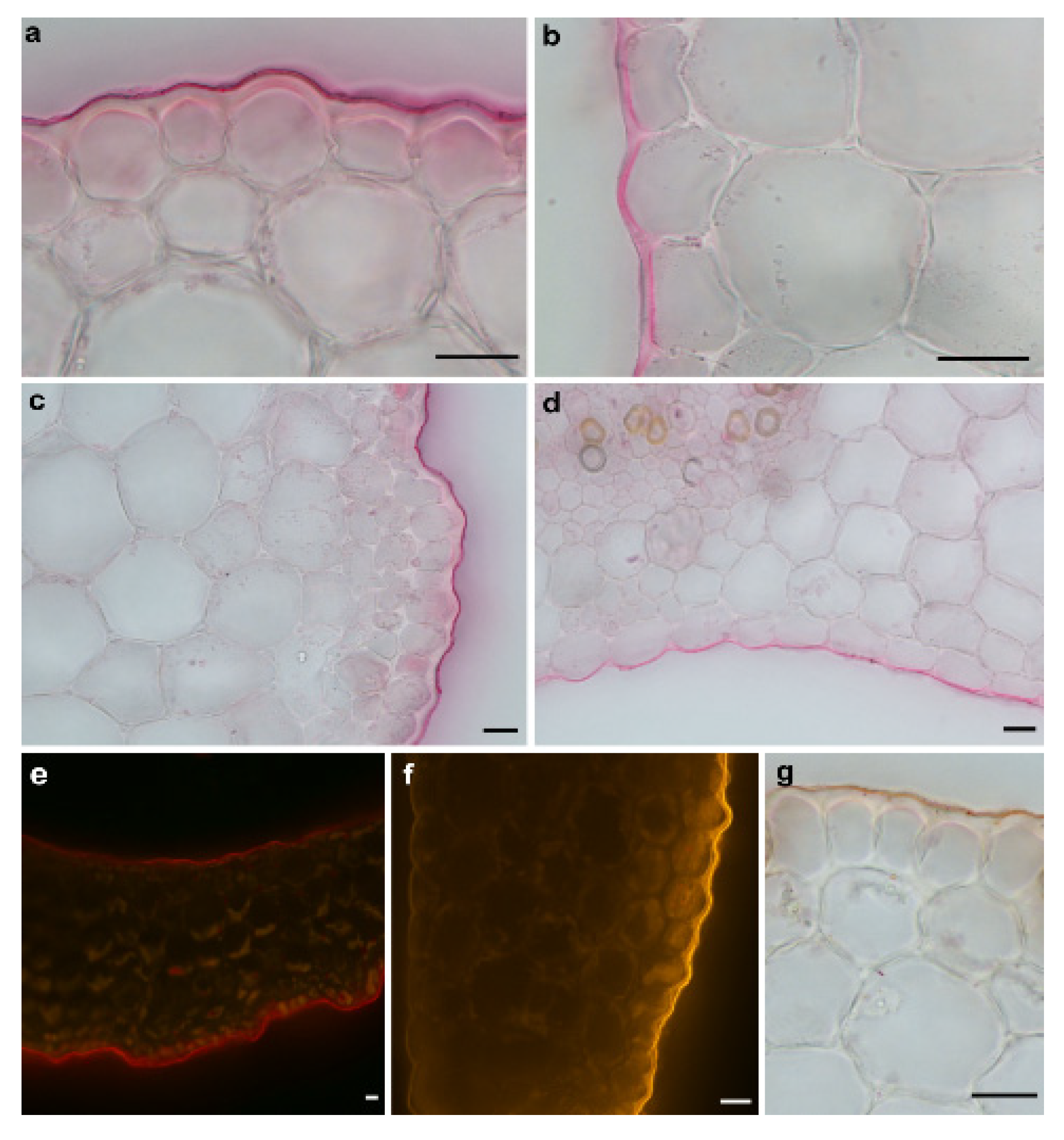
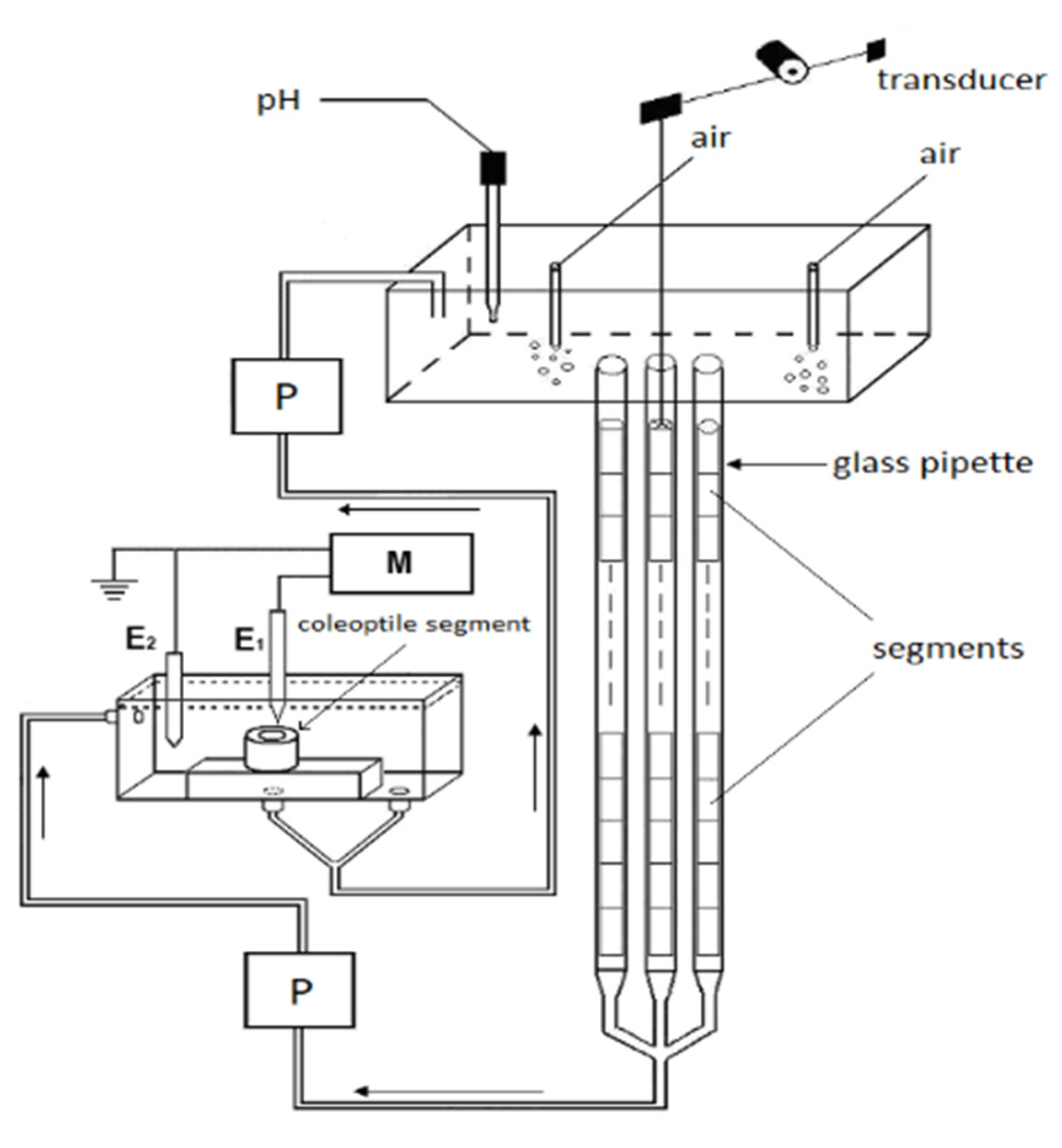
2.1. Methodological Aspects of the Experiments with Maize Coleoptile Segments
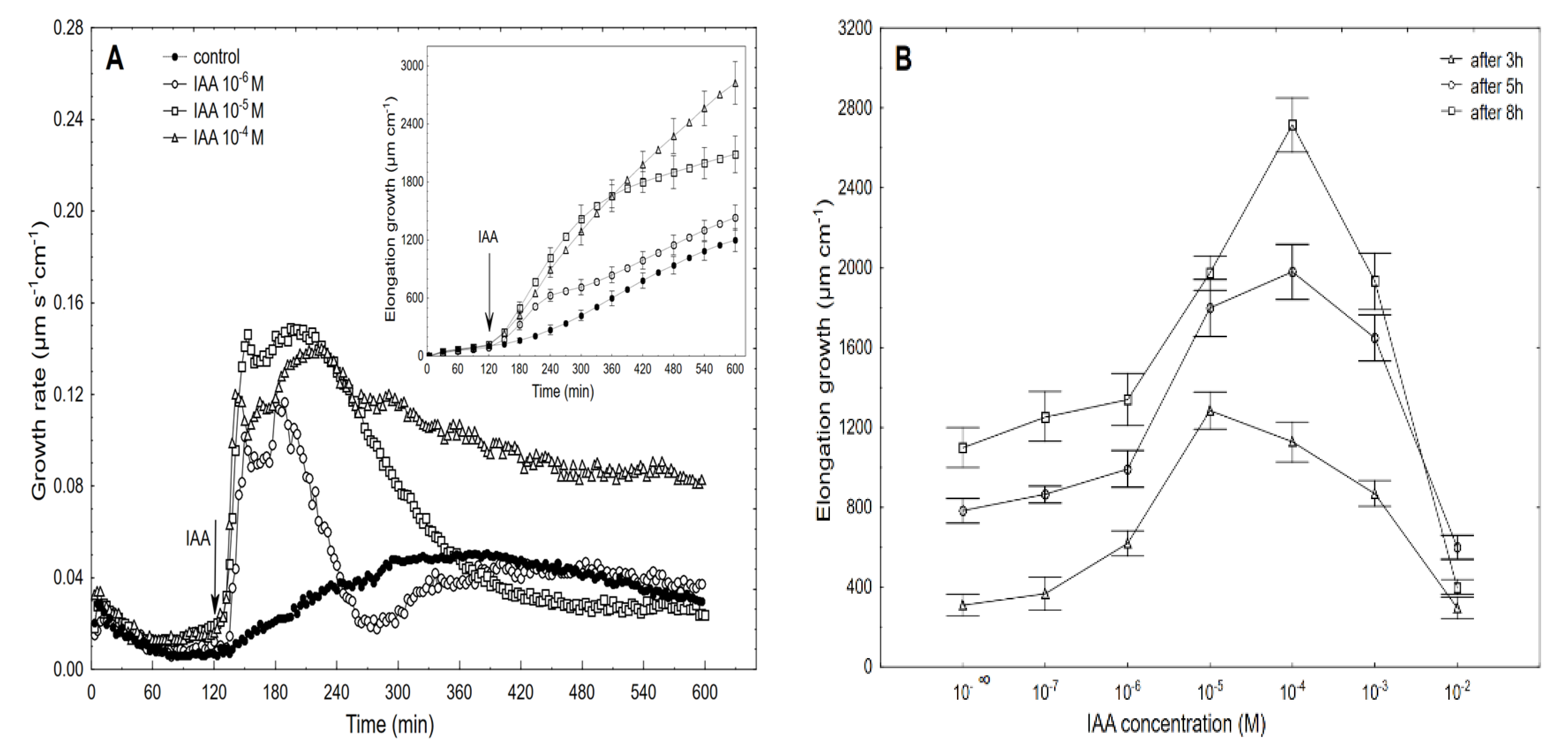
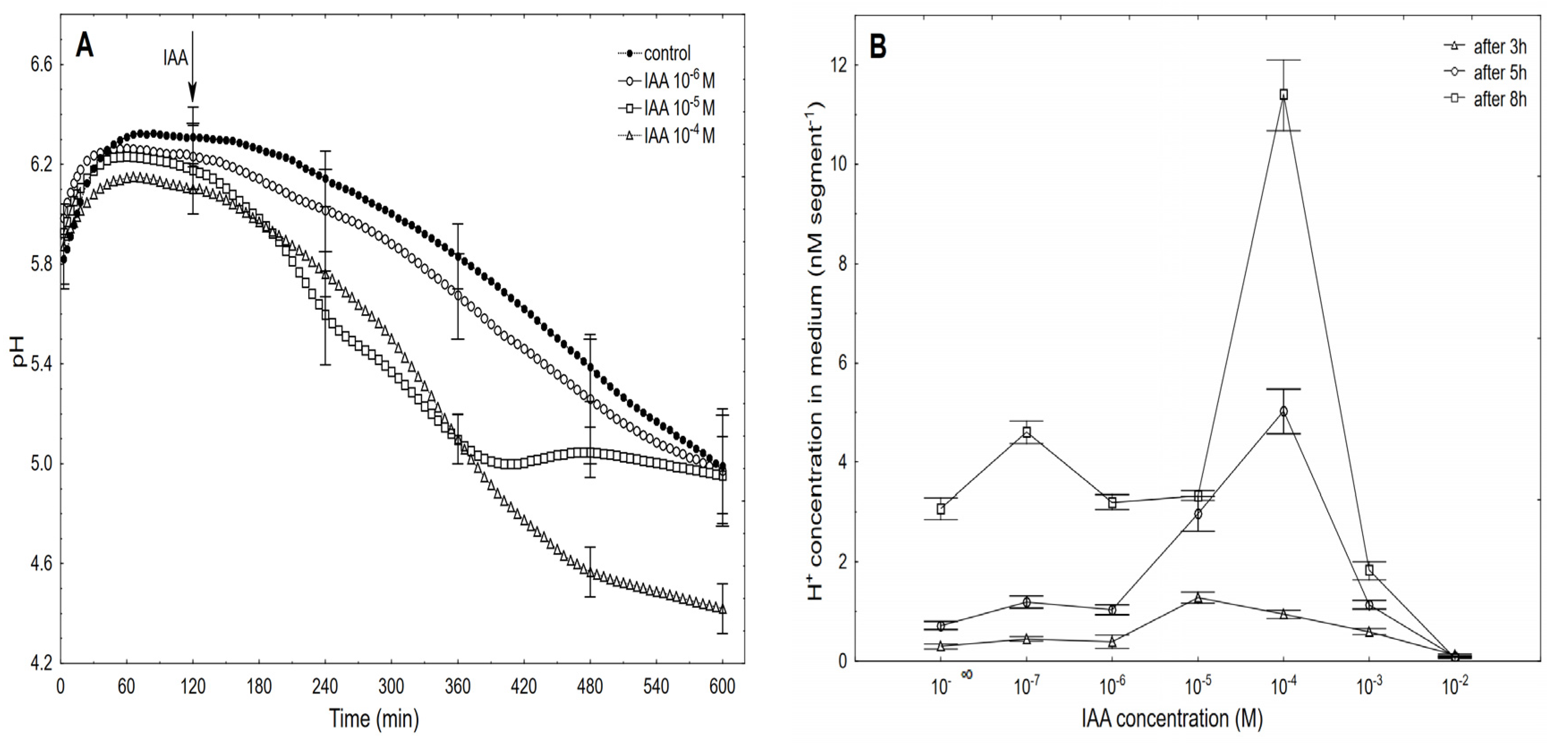
2.2. The Dose-Response Curves for the IAA-Induced Elongation Growth of Coleoptile Segments and Medium pH Measured Simultaneously with the Growth
2.3. Effect of Auxin on the Membrane Potential (Em) of the Parenchymal Cells Simultaneously Measured with the Growth and Medium pH of the Coleoptile Segments.
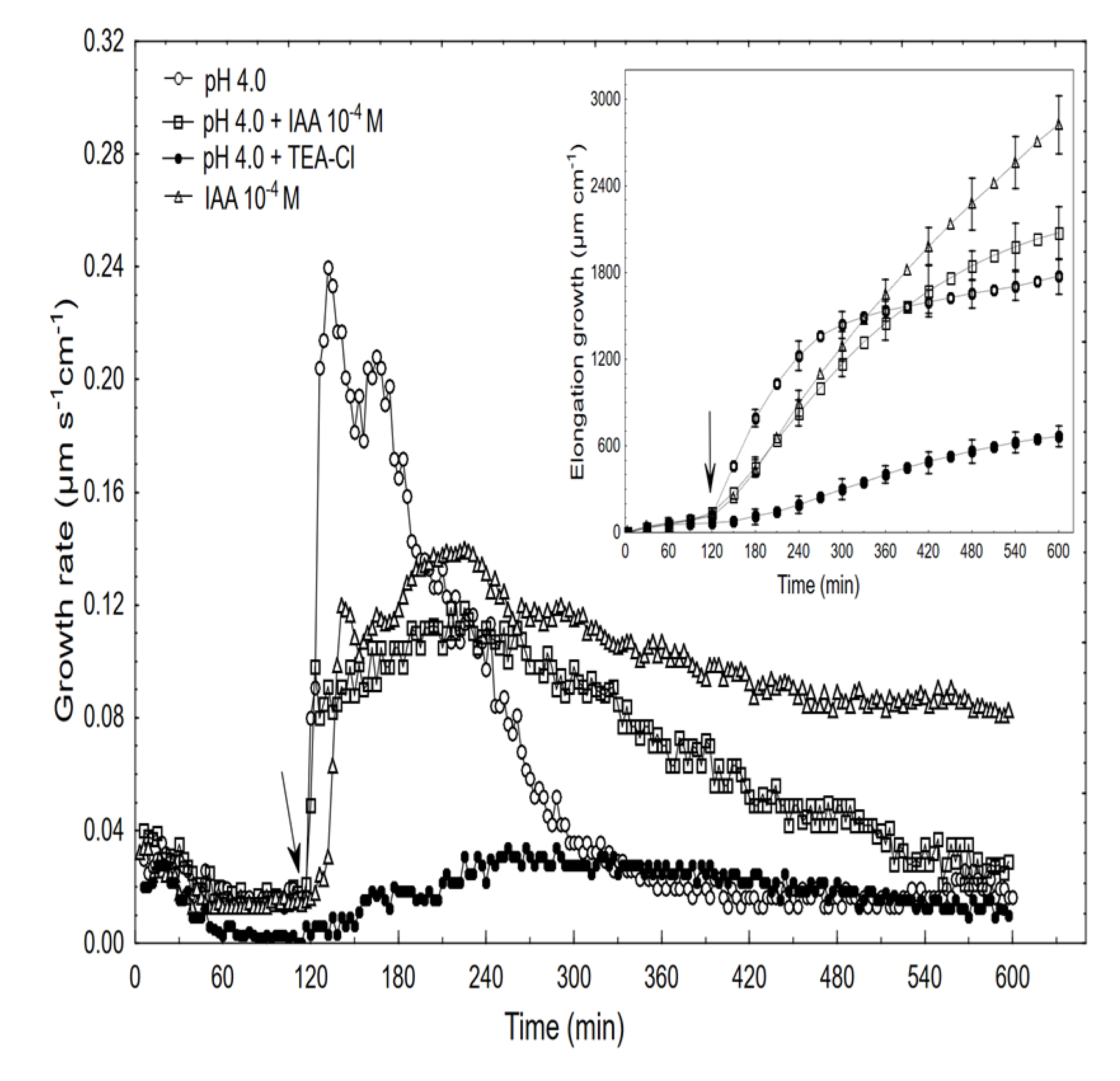
2.4. The Effects of Acid and Acid Combined with Auxin on the Growth and Membrane Potential of Coleoptile
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Growth, Medium pH and Membrane Potential Measurements
4.3. Histochemical Procedures and Staining
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rayle, D.L.; Cleland, R.E. Enhancement of wall loosening and elongation by acid solutions. Plant Physiol. 1970, 46, 250–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleland, R.E. Cell wall extension. Annu. Rev. Plant Physiol. 1971, 22, 197–222. [Google Scholar] [CrossRef]
- Hager, A.; Menzel, H.; Krauss, A. Versuche und Hypothese zur Primärwirkung des Auxins beim Streckungswachstum. Planta 1971, 100, 1–15. [Google Scholar] [CrossRef]
- Hager, A. Role of the plasma membrane H+-ATPase in auxin-induced elongation growth: Historical and new aspects. J. Plant Res. 2003, 116, 483–505. [Google Scholar] [CrossRef]
- Karcz, W.; Stolarek, J.; Pietruszka, M.; Malkowski, E. The dose–response curves for IAA-induced elongation growth and acidification of the incubation medium of Zea mays L. coleoptile segments. Physiol. Plant 1990, 80, 257–261. [Google Scholar] [CrossRef]
- Lüthen, H.; Bigdon, M.; Böttger, M. Re-examination of the acid-growth theory of auxin action. Plant Physiol. 1990, 93, 931–939. [Google Scholar] [CrossRef] [Green Version]
- Cleland, R.E.; Buckley, G.; Nowbar, S.; Lew, N.; Stinemetz, C.; Evans, M.; Rayle, D. The pH profile for acid-induced elongation of coleoptile and epicotyl section is consistent with the acid-growth theory. Planta 1991, 186, 70–74. [Google Scholar] [CrossRef]
- Rayle, D.L.; Cleland, R.E. The acid-growth theory of auxin induced cell elongation is alive and well. Plant Physiol. 1992, 99, 1271–1274. [Google Scholar] [CrossRef] [Green Version]
- Pope, D.G. Does indoleacetic acid promote growth via wall acidification? Planta 1978, 140, 137–142. [Google Scholar] [CrossRef]
- Kutschera, U.; Schopfer, P. Evidence against the acid-growth theory of auxin action. Planta 1985, 163, 483–493. [Google Scholar] [CrossRef]
- Schopfer, P. pH-dependence of extension growth in Avena coleoptiles and its implications for the mechanism of auxin action. Plant Physiol. 1989, 90, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Kutschera, U. The current status of the acid-growth hypothesis. New Phytol. 1994, 126, 549–569. [Google Scholar] [CrossRef]
- Kutschera, U. Acid growth and plant development. Science 2006, 311, 952–953. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, U.; Wang, Z.-Y. Growth-limiting proteins in maize coleoptiles and the auxin-brassinosteroid hypothesis of mesocotyl elongation. Protoplasma 2015, 253, 3–14. [Google Scholar] [CrossRef]
- Kutschera, U.; Khanna, R. Auxin action in developing maize coleoptiles: Challenges and open questions. Plant Signal. Behav. 2020. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Hedrich, R. Channeling auxin action: Modulation of ion transport by indole-3-acetic acid. Plant Mol. Biol. 2002, 49, 349–356. [Google Scholar] [CrossRef]
- Sauer, M.; Robert, S.; Kleine-Vehn, J. Auxin: Simply complicated. J. Exp. Bot. 2013, 64, 2565–2577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fendrych, M.; Leung, J.; Friml, J. TIR1/AFB-Aux/IAA auxin perception mediates rapid cel wall acidification and growth of Arabidopsis hypocotyls. eLife 2016. [Google Scholar] [CrossRef]
- Arsuffi, G.; Braybrook, S. Acid growth: An ongoing trip. J. Exp. Bot. 2019, 69, 137–146. [Google Scholar] [CrossRef]
- Ma, Q.; Grones, P.; Robert, S. Auxin signaling: A big question to be addressed by small molecules. J. Exp. Bot. 2018, 69, 313–328. [Google Scholar] [CrossRef] [Green Version]
- Cleland, R.E. Auxin and cell elongation. Physiology, biochemistry, and molecular biology. In Plant Hormones; Davies, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 214–217. [Google Scholar]
- Cleland, R.E.; Prins, H.B.A.; Harper, J.R.; Higinbotham, N. Rapid hormone-induced hyperpolarization of the oat coleoptile transmembrane potential. Plant Physiol. 1977, 59, 395–397. [Google Scholar] [CrossRef] [Green Version]
- Felle, H.H.; Peters, W.S.; Palme, K. The electrical response of maize to auxins. Biochim. Biophys. Acta 1991, 1064, 199–204. [Google Scholar] [CrossRef]
- Keller, C.P.; Van Volkenburgh, E. The electrical response of Avena coleoptile cortex to auxins: Evidence in vivo for activation of a Cl− conductance. Planta 1996, 198, 404–412. [Google Scholar] [CrossRef]
- Karcz, W.; Burdach, Z. A comparison of the effects of IAA and 4-Cl-IAA on growth, proton secretion and membrane potential in maize coleoptile segments. J. Exp. Bot. 2002, 53, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Siemieniuk, A.; Karcz, W. Effect of K+ and Ca2+ on the indole-3-acetic acid- and fusicoccin-induced growth and membrane potential in maize coleoptile cells. AoB Plants 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rücke, A.; Palme, K.; Venis, M.A.; Napier, R.M.; Felle, H.H. Patch-clamp analysis establishes a role for an auxin binding protein in the auxin stimulation of plasma membrane current in Zea mays protoplasts. Plant J. 1993, 4, 41–46. [Google Scholar] [CrossRef]
- Philippar, K.; Fuchs, I.; Lüthen, H.; Hoth, S.; Bauer, C.S.; Haga, K.; Thiel, G.; Ljung, K.; Sandberg, G.; Böttger, M.; et al. Auxin-induced K+ channel expression an essential step in coleoptile growth and gravitropism. Proc. Nat. Acad. Sci. USA 1999, 96, 12186–12191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiel, G.; Weise, R. Auxin augments conductance of K+ inward rectifier in maize protoplasts. Planta 1999, 208, 38–45. [Google Scholar] [CrossRef]
- Philippar, K.; Ivashikina, N.; Ache, P.; Christian, M.; Lüthen, H.; Palme, K.; Hedrich, R. Auxin activates KAT1 and KAT2, two K+- channel genes expressed in seedlings of Arabidopsis thaliana. Plant J. 2004, 37, 815–827. [Google Scholar] [CrossRef]
- Frias, I.; Caldeira, M.T.; Perez-Castineira, J.R.; Navarro-Avino, J.P.; Culianzez-Macia, F.; Kuppinger, O.; Stransky, H.; Pages, M.; Hager, A.; Serrano, R. A major isoform of the maize plasma membrane H+-ATPase: Characterization and induction by auxin in coleoptiles. Plant Cell 1996, 8, 1533–1544. [Google Scholar] [PubMed] [Green Version]
- Burdach, Z.; Siemieniuk, A.; Trela, Z.; Kurtyka, R.; Karcz, W. Role of auxin (IAA) in the regulation of slow vacuolar (SV) channels and the volume of red beet taproot vacuoles. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Burdach, Z.; Siemieniuk, A.; Karcz, W. Effect of auxin (IAA) on the fast vacuolar (FV) channels in red beet (Beta vulgaris L.) taproot vacuoles. Inter. J. Mol. Sci. 2020, 21, 4876. [Google Scholar] [CrossRef]
- Karcz, W.; Stolarek, J.; Lekacz, H.; Kurtyka, R.; Burdach, Z. Comparative investigation of auxin and fusicoccin-induced growth and H+-extrusion in coleoptile of Zea mays L. Acta Physiol. Plant. 1995, 17, 3–8. [Google Scholar]
- Hager, A.; Debus, G.; Edel, H.G.; Stransky, H.; Serrano, R. Auxin induces exocytosis and the rapid synthesis of a high-turnover pool of plasma membrane H+-ATPase. Planta 1991, 185, 527–537. [Google Scholar] [CrossRef]
- Park, W.J.; Schäfer, A.; Prinsen, E.; van Onckelen, H.; Kang, B.G.; Hertel, R. Auxin-induced elongation of short maize coleoptile segments is supported by 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one. Planta 2001, 213, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Jeffree, C.E. The fine structure of the plant cuticle. In Biology of the Plant Cuticle; Riederer, M., Muller, C., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 16–49. [Google Scholar]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Peters, W.S.; Felle, H. Control of apoplast pH in corn coleoptile segments. II. The effect of various auxins and auxin analogues. J. Plant Physiol. 1991, 137, 691–696. [Google Scholar] [CrossRef]
- Claussen, M.; Lüthen, H.; Blatt, M.; Böttger, M. Auxin-induced growth and its linkage to potassium channels. Planta 1997, 201, 227–234. [Google Scholar] [CrossRef]
- Tode, K.; Lüthen, H. Fusicoccin- and IAA-induced elongation growth share the same pattern of K+ dependence. J. Exp. Bot. 2001, 52, 251–255. [Google Scholar]
- Burdach, Z.; Kurtyka, R.; Siemieniuk, A.; Karcz, W. Role of chloride ions in the promotion of auxin-induced growth of maize coleoptile segments. Ann. Bot. 2014, 114, 1023–1034. [Google Scholar] [CrossRef] [Green Version]
- Kurtyka, R.; Burdach, Z.; Siemieniuk, A.; Karcz, W. Single and combined effects of Cd and Pb on the growth, medium pH, membrane potential and metal content in maize (Zea mays L.) coleoptile segments. Ecotox. Environ. Saf. 2018, 161, 8–16. [Google Scholar] [CrossRef]
- Falhof, J.; Pedersen, J.T.; Fuglsang, A.T.; Palmgren, M. Plasma membrane H+-ATPase regulation in the centre of plant physiology. Mol. Plant 2016, 9, 323–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Navarro, A. Potassium transport in fungi and plants. Biochim. Biophys. Acta 2000, 1469, 1–30. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology; Sinauer Associates, Inc.: Sunderland, MA, USA, 2010; p. 782. [Google Scholar]
- Kitasato, H. The influence of H+ on the membrane potential and ion fluxes of Nitella. J. Gen. Physiol. 1968, 52, 60–87. [Google Scholar] [CrossRef] [Green Version]
- Thibaud, J.B.; Soler, A.; Grignon, C. H+ and K+ electrogenic exchanges in corn roots. Plant Physiol. 1986, 81, 847–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopfer, P. Hydroxyl radical-induced cell-wall loosening in vitro and in vivo: Implications for the control of elongation growth. Plant J. 2001, 28, 679–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticle. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef]
- Dominguez, E.; Heredia-Guerrero, J.A.; Heredia, A. The biophysical design of plant cuticle: An overview. New Phytol. 2011, 189, 938–949. [Google Scholar] [CrossRef]
- Zeisler-Diehl, V.V.; Migdal, B.; Schreiber, L. Quantitative characterization of cuticular barrier properties: Methods, requirements, and problems. J. Exp. Bot. 2017, 68, 5281–5291. [Google Scholar] [CrossRef] [Green Version]
- Cleland, R.E. The dose-response curve for auxin-induced cell elongation. A re-evaluation. Planta 1972, 104, 1–9. [Google Scholar] [CrossRef]
- Nissen, P. Dose responses of auxins. Physiol. Plant. 1985, 65, 357–374. [Google Scholar] [CrossRef]
- Karcz, W.; Lüthen, H.; Böttger, M. Effect of IAA and 4-Cl-IAA on growth rate in maize coleoptile segments. Acta Physiol. Plant. 1999, 21, 133–139. [Google Scholar] [CrossRef]
- Polak, M.; Tukaj, Z.; Karcz, W. Effect of temperature on the dose-response curves for auxin-induced elongation growth in maize coleoptile segments. Acta Physiol. Plant. 2011, 33, 437–442. [Google Scholar] [CrossRef]
- Rudnicka, M.; Ludynia, M.; Karcz, W. The effect of naphthazarin on the growth electrogenicity, oxidative stress, and microtubule array in Z. mays coleoptile cells treated with IAA. Front. Plant Sci. 2019. [Google Scholar] [CrossRef]
- Rudnicka, M.; Ludynia, M.; Karcz, W. Effects of naphthazarin (DHNQ) combined with lawsone (NQ-2-OH) or 1,4 naphthoquinone (NQ) on the auxin-induced growth of Zea mays L. coleoptile segments. Int. J. Mol. Sci. 2019, 20, 1788. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Bregante, M.; Dreyer, I.; Gambale, F. The voltage-dependent potassium-uptake channel of corn coleoptiles has permeation properties different from other K+ channels. Planta 1995, 197, 193–199. [Google Scholar] [CrossRef]
- Bates, G.W.; Goldsmith, M.H.M. Rapid response of the plasma-membrane potential of oat coleoptiles to auxin and other weak acids. Planta 1983, 159, 231–237. [Google Scholar] [CrossRef]
- Paponov, I.A.; Dindas, J.; Król, E.; Friz, T.; Budryk, V.; Teale, W.; Paponov, M.; Hedrich, R.; Palme, K. Auxin-induced plasma membrane depolarization is regulated by auxin transport and not by AUXIN BINDING PROTEIN1. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, I.; Michard, E. High- and low-affinity transport in plants from a thermodynamic point of view. Front. Plant Sci. 2020. [Google Scholar] [CrossRef] [Green Version]
- Spalding, E.P.; Hirsch, R.E.; Lewis, D.R.; Qi, Z.; Sussman, M.R.; Lewis, B.D. Potassium uptake supporting plant growth in the absence of AKT1 channel activity–inhibition by ammonium and stimulation by sodium. J. Gen. Physiol. 1999, 113, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, L.M. Auxin-induced growth in cut segments of grass coleoptiles or dicot stems. In Plant Growth and Development: Hormones and Environment; Academic Press: New York, NY, USA; Elsevier Science: New York, NY, USA, 2002; pp. 341–378. [Google Scholar]
- Polak, M.; Zaborska, W.; Tukaj, Z.; Karcz, W. Effect of thiosulphinates contained in Garlic extract on growth, proton fluxes and membrane potential in maize (Zea mays L.) coleoptile segments. Acta Physiol. Plant 2012, 34, 41–52. [Google Scholar] [CrossRef]
- Buda, G.J.; Isaacson, T.; Matas, A.J.; Paolillo, D.J.; Rose, J.K.C. Three-dimensional imaging of plant cuticle architecture using confocal scanning laser microscopy. Plant J. 2009, 60, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Potocka, I.; Baldwin, T.C.; Kurczyńska, E.U. Distribution of lipid transfer protein 1 (LTP1) epitopes associated with morphogenic events during somatic embryogenesis of Arabidopsis thaliana. Plant Cell Rep. 2012, 31, 2031–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Giorgi, J.; Piskurewicz, U.; Loubery, S.; Utz-Pugin, A.; Bailly, C.; Mène-Saffrané, L.; Lopez-Molina, L. An endosperm-associated cuticle is required for arabidopsis seed viability, dormancy and early control of germination. PLoS Gen. 2015, 11, e1005708. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polak, M.; Karcz, W. Some New Methodological and Conceptual Aspects of the “Acid Growth Theory” for the Auxin Action in Maize (Zea mays L.) Coleoptile Segments: Do Acid- and Auxin-Induced Rapid Growth Differ in Their Mechanisms? Int. J. Mol. Sci. 2021, 22, 2317. https://doi.org/10.3390/ijms22052317
Polak M, Karcz W. Some New Methodological and Conceptual Aspects of the “Acid Growth Theory” for the Auxin Action in Maize (Zea mays L.) Coleoptile Segments: Do Acid- and Auxin-Induced Rapid Growth Differ in Their Mechanisms? International Journal of Molecular Sciences. 2021; 22(5):2317. https://doi.org/10.3390/ijms22052317
Chicago/Turabian StylePolak, Małgorzata, and Waldemar Karcz. 2021. "Some New Methodological and Conceptual Aspects of the “Acid Growth Theory” for the Auxin Action in Maize (Zea mays L.) Coleoptile Segments: Do Acid- and Auxin-Induced Rapid Growth Differ in Their Mechanisms?" International Journal of Molecular Sciences 22, no. 5: 2317. https://doi.org/10.3390/ijms22052317