
Targeting mTOR-CCL20 Signaling May Improve Response to Docetaxel in Head and Neck Squamous Cell Carcinoma

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
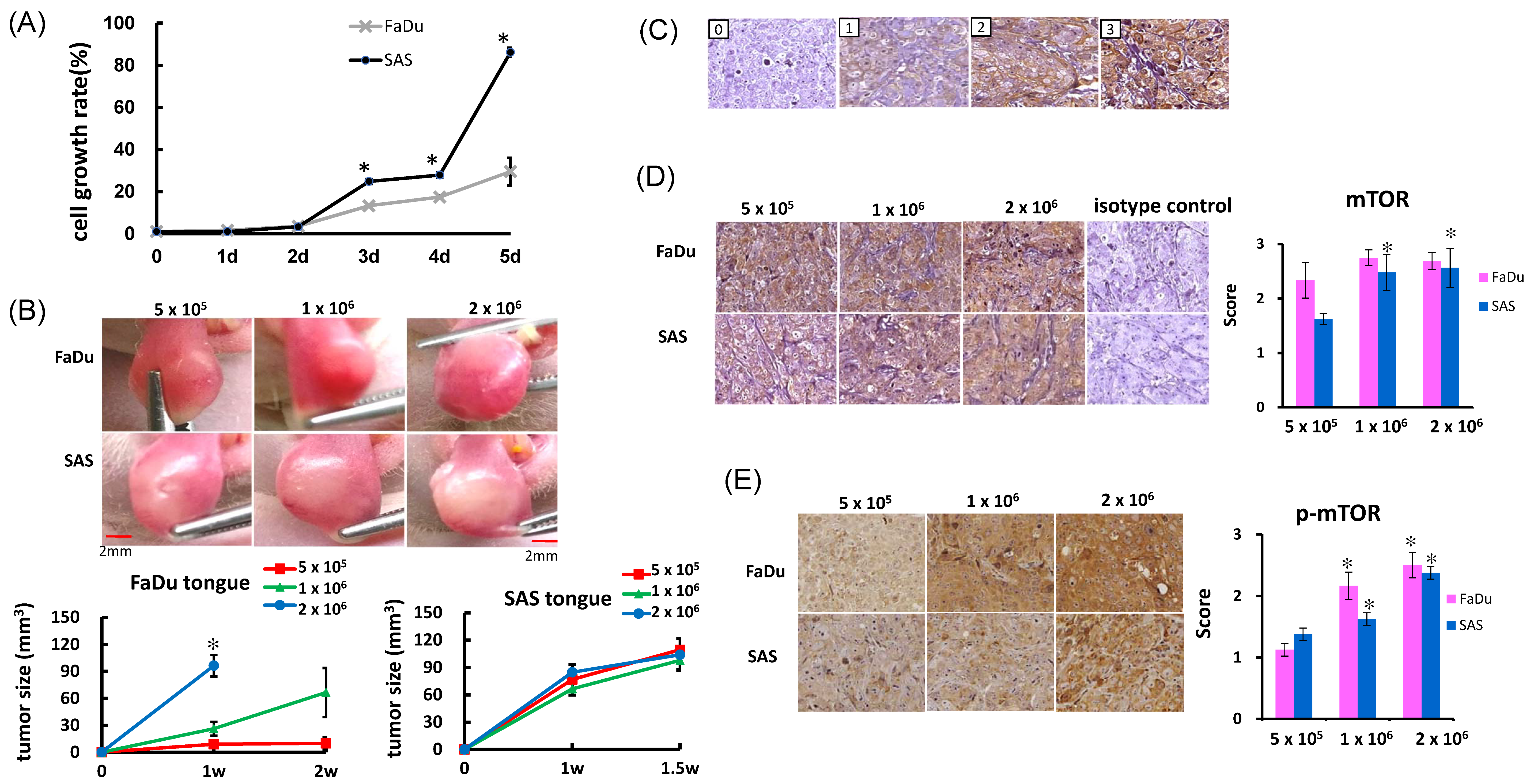
2.1. In Vitro Cell Growth and In Vivo Tumor Growth
2.2. Immunohistochemical Staining for mTOR in NU/NU Mouse Xenograft Models
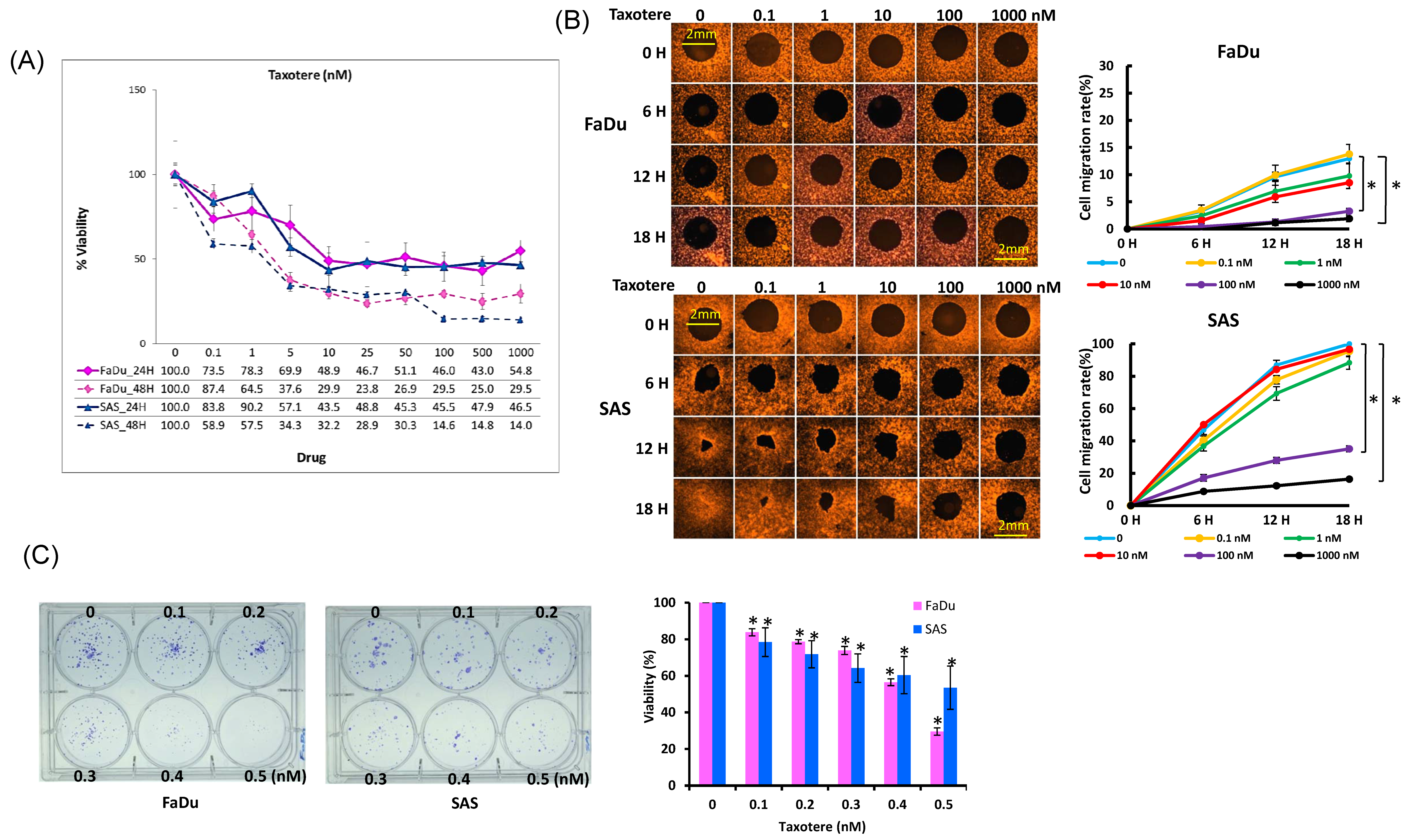
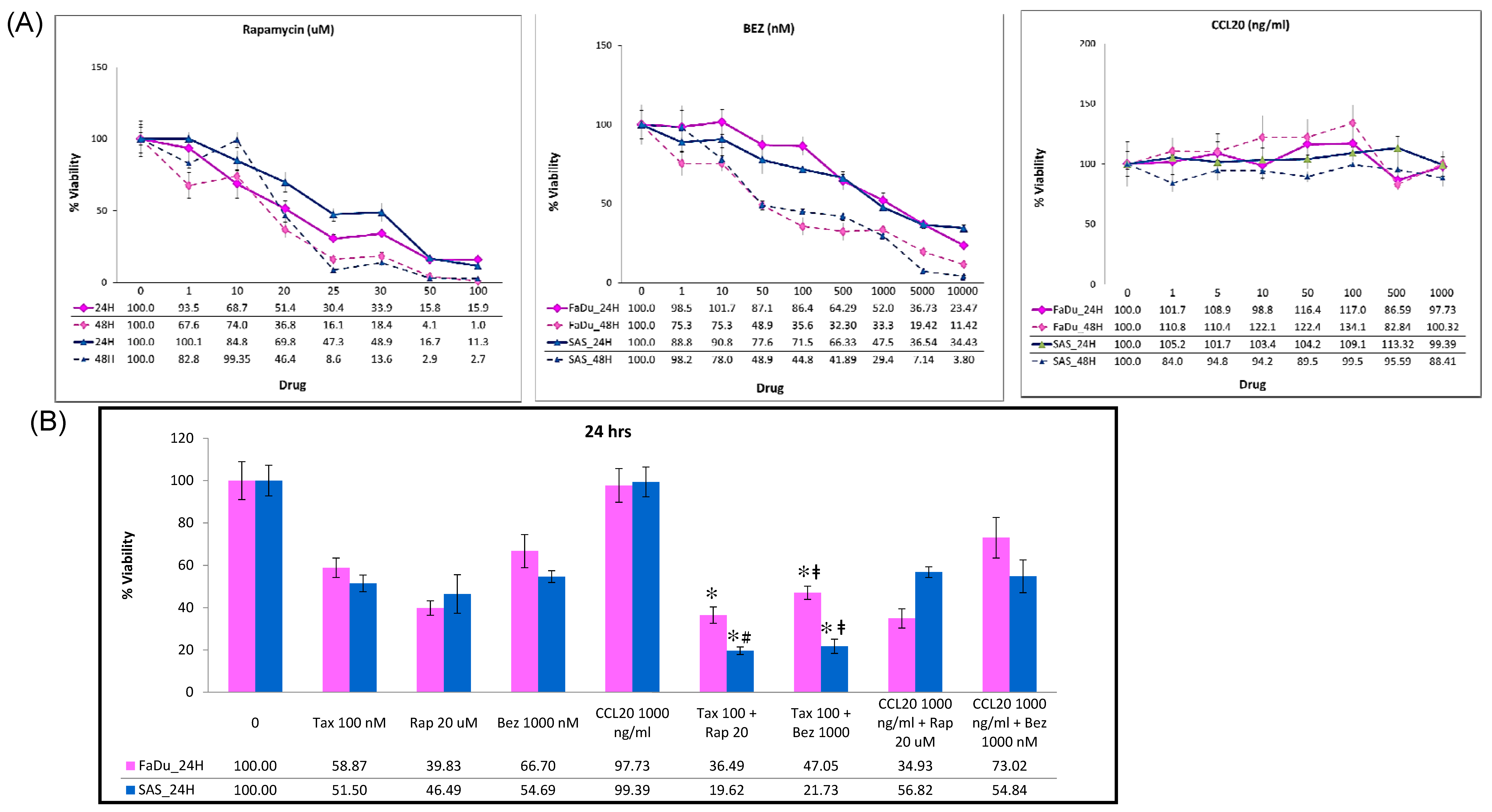
2.3. Cytotoxic Activity of Docetaxel on HNSCC Cell Lines
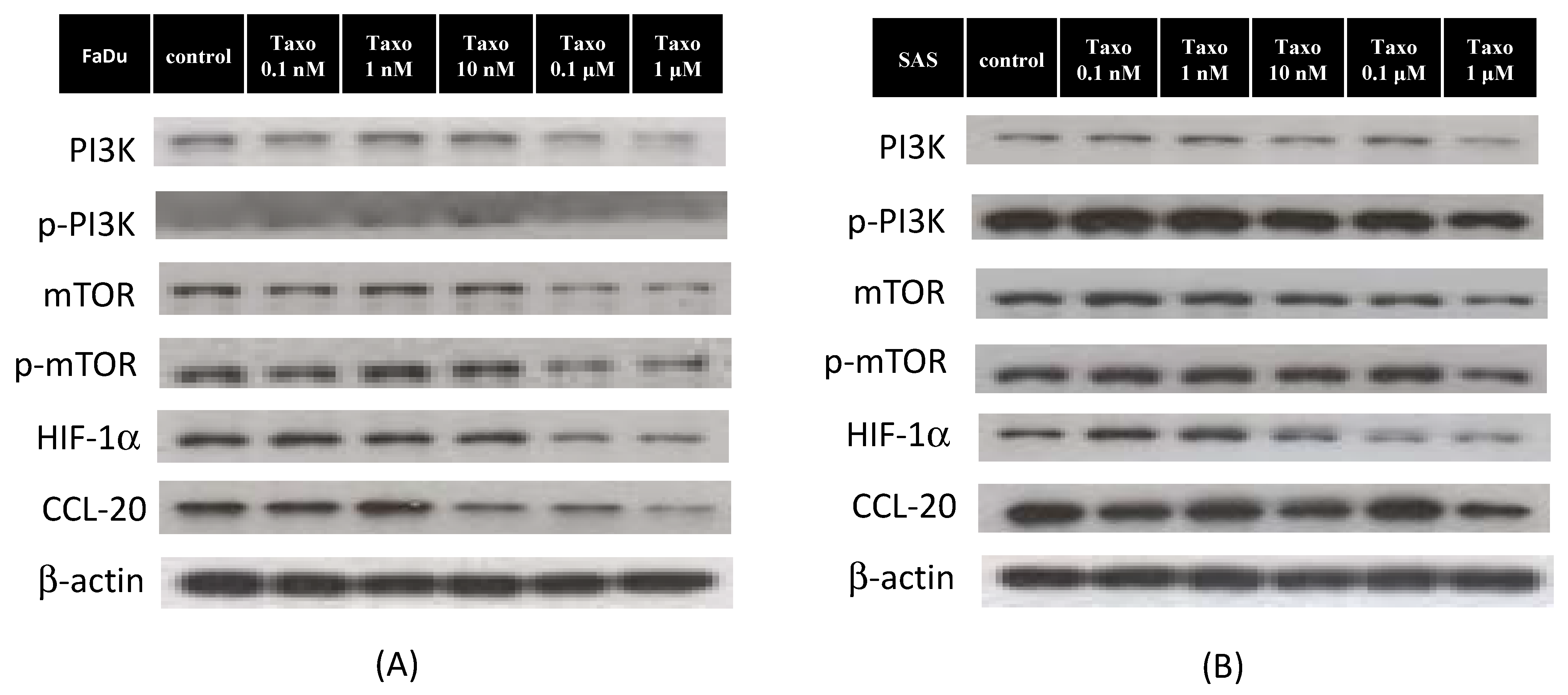
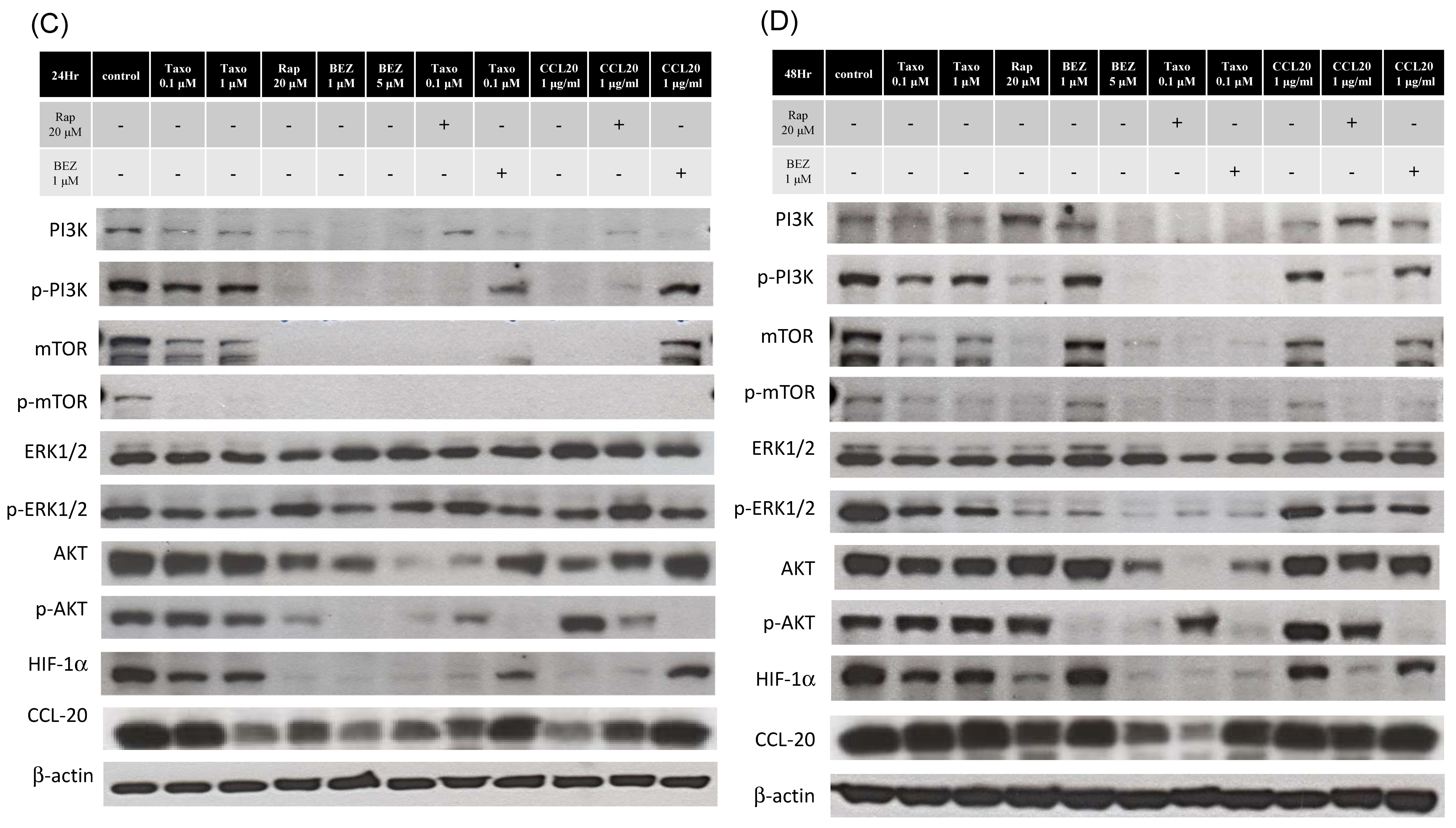
2.4. Docetaxel Inhibits mTOR-CCL20 Expression in HNSCC Cells
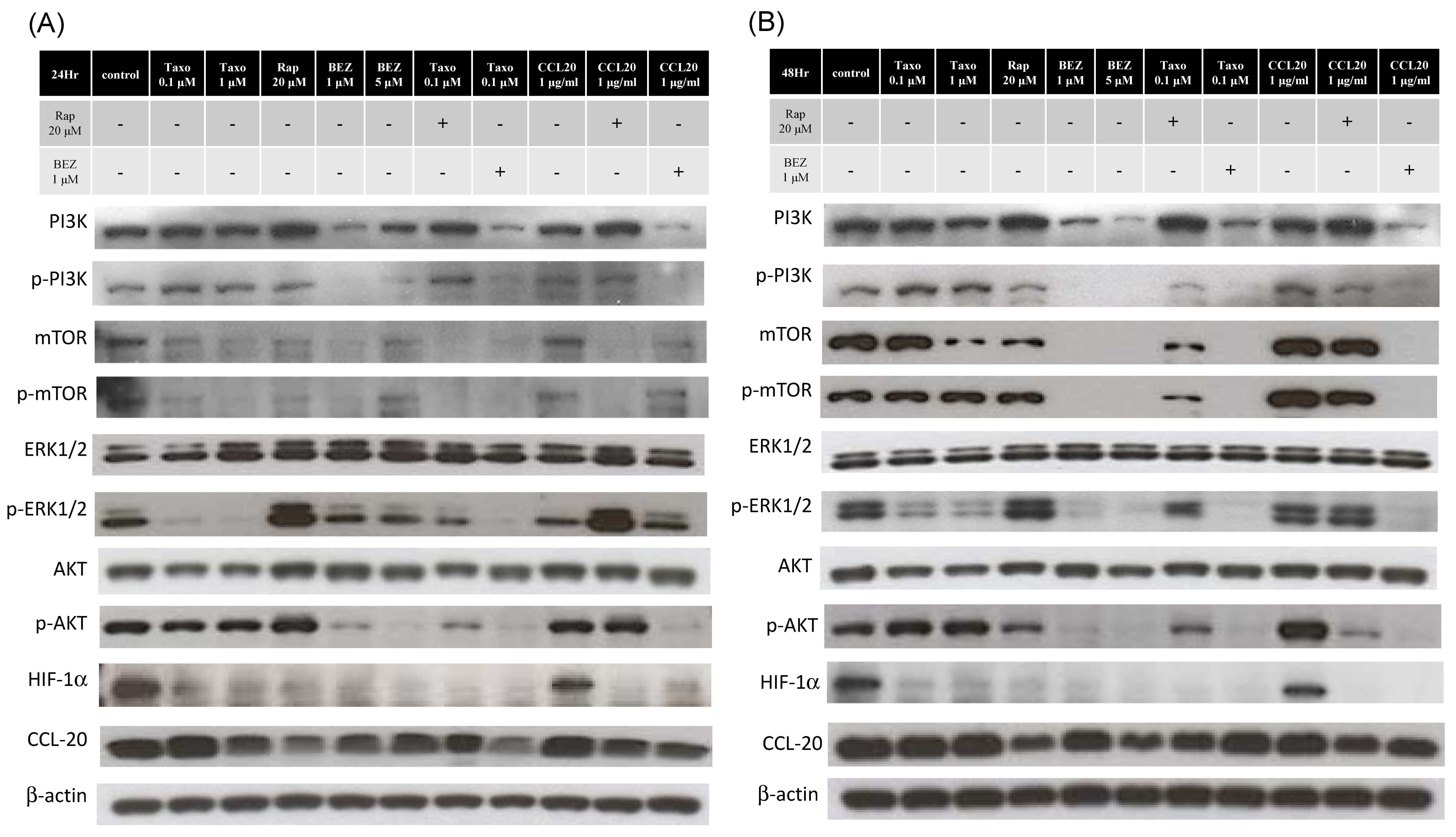
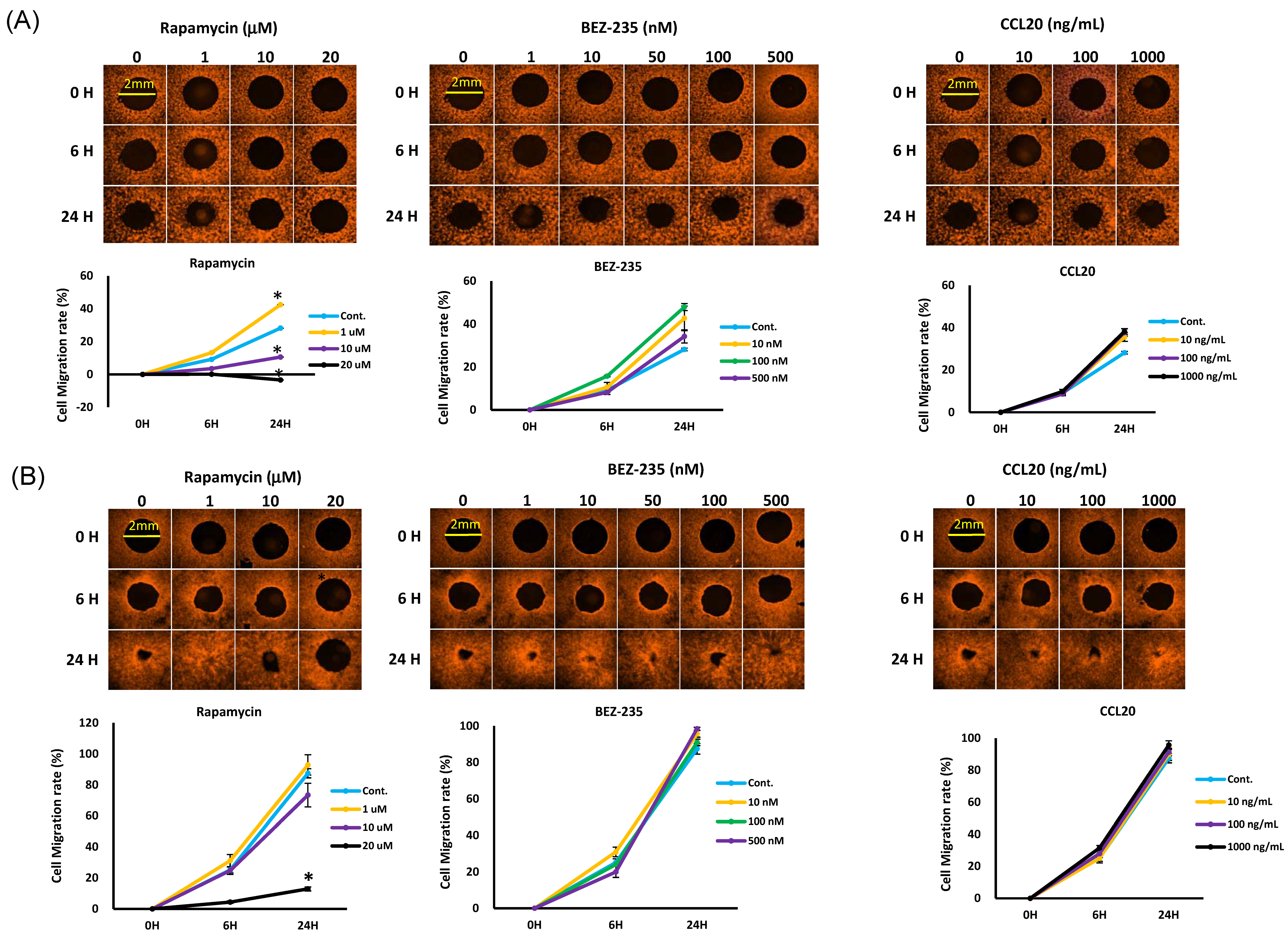
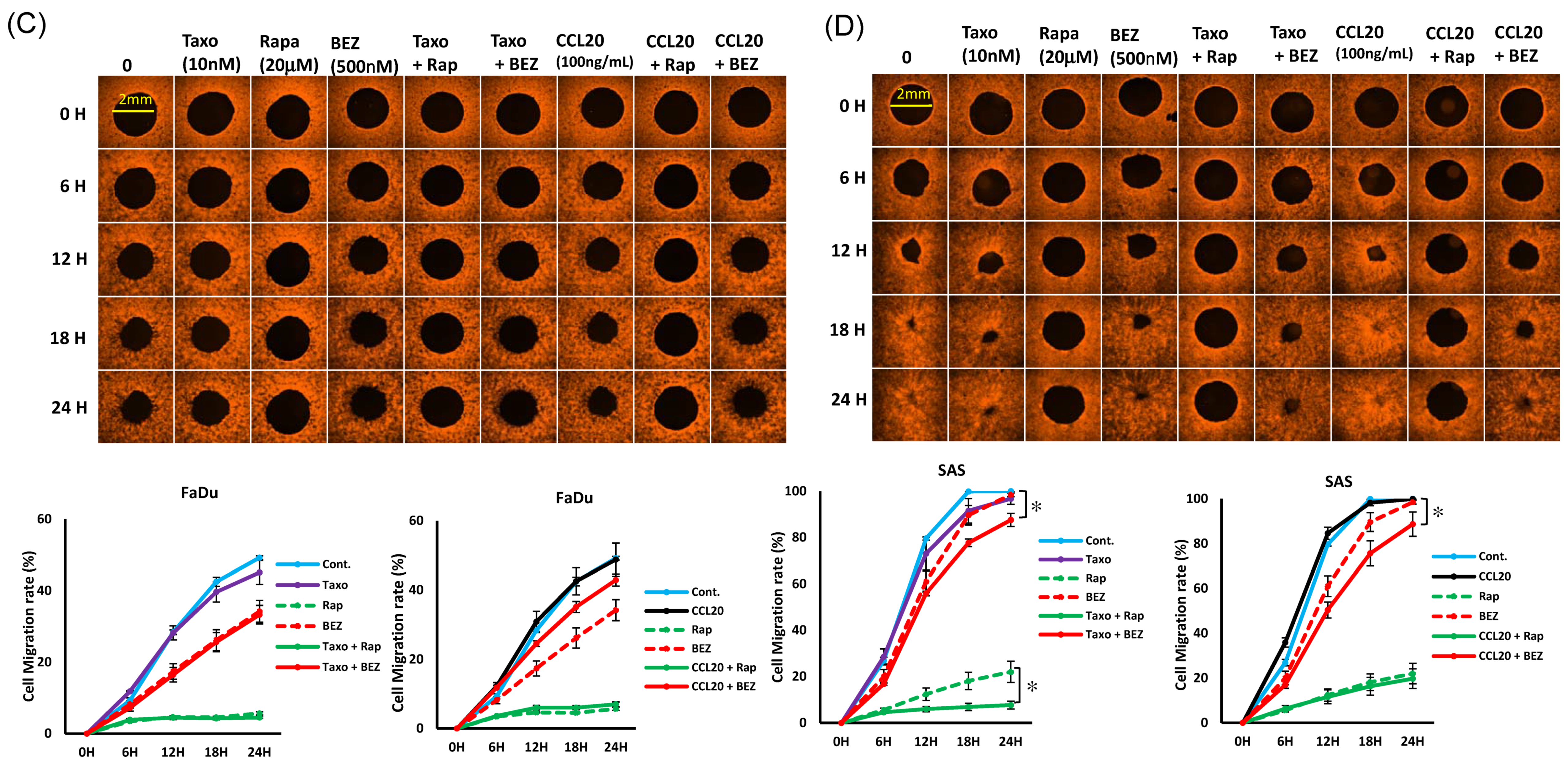
2.5. mTOR Inhibitors Enhance the Effects of Docetaxel to Inhibit PI3K/mTOR/CCL-20 Signaling and HNSCC Cell Proliferation/Migration
3. Discussion
4. Materials and Methods
4.1. HNSCC Cell Cultures
4.2. Cell Growth Assay
4.3. Cell Viability Analysis (WST1)
4.4. Clonogenic Assay
4.5. Western Blot Analysis
4.6. Oris™ Cell Migration Assay
4.7. Animal Care
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HNSCC | head and neck squamous cell carcinoma |
| GLOBOCAN | Global Cancer Observatory CANCER TODAY |
| CRT | chemoradiation therapy |
| PI3K/AKT/mTOR | phosphoinositide 3-kinase/protein kinase B/mammalian target of rapamycin |
| CCL20 | Chemokine (C-C motif) ligand 20 |
| HIF-1α | Hypoxia-Inducible Factor-1α |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.-L.; Kelly, J.Y.; Chiang, C.-J.; Chen, T.-C.; Wang, C.-P. Head and Neck Cancer Incidence Trends in Taiwan, 1980–2014. Int. J. Head Neck Sci. 2017, 1, 180–190. [Google Scholar]
- Hung, L.C.; Kung, P.T.; Lung, C.H.; Tsai, M.H.; Liu, S.A.; Chiu, L.T.; Huang, K.H.; Tsai, W.C. Assessment of the Risk of Oral Cancer Incidence in a High-Risk Population and Establishment of a Predictive Model for Oral Cancer Incidence Using a Population-Based Cohort in Taiwan. Int. J. Environ. Res. Public Health 2020, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posner, M.R.; Hershock, D.M.; Blajman, C.R.; Mickiewicz, E.; Winquist, E.; Gorbounova, V.; Tjulandin, S.; Shin, D.M.; Cullen, K.; Ervin, T.J.; et al. Cisplatin and fluorouracil alone or with docetaxel in head and neck cancer. N. Engl. J. Med. 2007, 357, 1705–1715. [Google Scholar] [CrossRef] [Green Version]
- Vermorken, J.B.; Remenar, E.; van Herpen, C.; Gorlia, T.; Mesia, R.; Degardin, M.; Stewart, J.S.; Jelic, S.; Betka, J.; Preiss, J.H.; et al. Cisplatin, fluorouracil, and docetaxel in unresectable head and neck cancer. N. Engl. J. Med. 2007, 357, 1695–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, H.; Pammer, J.; Minichsdorfer, C.; Posch, D.; Kornek, G.; Aretin, M.B.; Fuereder, T. Modified biweekly cisplatin, docetaxel plus cetuximab (TPEx) as first-line treatment for patients with recurrent/metastatic head and neck cancer. Med. Oncol. 2018, 35, 32. [Google Scholar] [CrossRef] [Green Version]
- Guigay, J.; Fayette, J.; Dillies, A.F.; Sire, C.; Kerger, J.N.; Tennevet, I.; Machiels, J.P.; Zanetta, S.; Pointreau, Y.; Bozec Le Moal, L.; et al. Cetuximab, docetaxel, and cisplatin as first-line treatment in patients with recurrent or metastatic head and neck squamous cell carcinoma: A multicenter, phase II GORTEC study. Ann. Oncol. 2015, 26, 1941–1947. [Google Scholar] [CrossRef] [PubMed]
- Lui, V.W.; Hedberg, M.L.; Li, H.; Vangara, B.S.; Pendleton, K.; Zeng, Y.; Lu, Y.; Zhang, Q.; Du, Y.; Gilbert, B.R.; et al. Frequent mutation of the PI3K pathway in head and neck cancer defines predictive biomarkers. Cancer Discov. 2013, 3, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Bader, A.G.; Vogt, P.K. Phosphatidylinositol 3-kinase mutations identified in human cancer are oncogenic. Proc. Natl. Acad. Sci. USA 2005, 102, 802–807. [Google Scholar] [CrossRef] [Green Version]
- Saidak, Z.; Clatot, F.; Chatelain, D.; Galmiche, A. A gene expression profile associated with perineural invasion identifies a subset of HNSCC at risk of post-surgical recurrence. Oral Oncol. 2018, 86, 53–60. [Google Scholar] [CrossRef]
- Xia, Y.; Jiang, L.; Zhong, T. The role of HIF-1alpha in chemo-/radioresistant tumors. Onco Targets Ther. 2018, 11, 3003–3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guru, S.K.; Pathania, A.S.; Kumar, S.; Ramesh, D.; Kumar, M.; Rana, S.; Kumar, A.; Malik, F.; Sharma, P.R.; Chandan, B.K.; et al. Secalonic Acid-D Represses HIF1alpha/VEGF-Mediated Angiogenesis by Regulating the Akt/mTOR/p70S6K Signaling Cascade. Cancer Res. 2015, 75, 2886–2896. [Google Scholar] [CrossRef] [Green Version]
- Marques, A.E.; Elias, S.T.; Porporatti, A.L.; Castilho, R.M.; Squarize, C.H.; De Luca Canto, G.; Guerra, E.N. mTOR pathway protein immunoexpression as a prognostic factor for survival in head and neck cancer patients: A systematic review and meta-analysis. J. Oral Pathol. Med. 2016, 45, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Jia, Y.; Ma, J.; Wu, S.; Jiang, H.; Cao, Y.; Sun, X.; Yin, X.; Yan, S.; Shang, M.; et al. Tumor-associated macrophage-derived CCL20 enhances the growth and metastasis of pancreatic cancer. Acta Biochim. Biophys. Sin. 2016, 48, 1067–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faes, S.; Demartines, N.; Dormond, O. Resistance to mTORC1 Inhibitors in Cancer Therapy: From Kinase Mutations to Intratumoral Heterogeneity of Kinase Activity. Oxid. Med. Cell Longev. 2017, 2017, 1726078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Li, X.; Zhang, J. mTOR Signaling in Cancer and mTOR Inhibitors in Solid Tumor Targeting Therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Lewis, C.; Nadel, J.A. CCL20/CCR6 feedback exaggerates epidermal growth factor receptor-dependent MUC5AC mucin production in human airway epithelial (NCI-H292) cells. J. Immunol. 2011, 186, 3392–3400. [Google Scholar] [CrossRef] [Green Version]
- Frick, V.O.; Rubie, C.; Kolsch, K.; Wagner, M.; Ghadjar, P.; Graeber, S.; Glanemann, M. CCR6/CCL20 chemokine expression profile in distinct colorectal malignancies. Scand. J. Immunol. 2013, 78, 298–305. [Google Scholar] [CrossRef]
- Du, D.; Liu, Y.; Qian, H.; Zhang, B.; Tang, X.; Zhang, T.; Liu, W. The effects of the CCR6/CCL20 biological axis on the invasion and metastasis of hepatocellular carcinoma. Int. J. Mol. Sci. 2014, 15, 6441–6452. [Google Scholar] [CrossRef] [Green Version]
- Kapur, N.; Mir, H.; Clark Iii, C.E.; Krishnamurti, U.; Beech, D.J.; Lillard, J.W.; Singh, S. CCR6 expression in colon cancer is associated with advanced disease and supports epithelial-to-mesenchymal transition. Br. J. Cancer 2016, 114, 1343–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadomoto, S.; Izumi, K.; Mizokami, A. The CCL20-CCR6 Axis in Cancer Progression. Int. J. Mol. Sci. 2020, 21, 5186. [Google Scholar] [CrossRef]
- Chen, C.H.; Chuang, H.C.; Lin, Y.T.; Fang, F.M.; Huang, C.C.; Chen, C.M.; Lu, H.; Chien, C.Y. Circulating CD105 shows significant impact in patients of oral cancer and promotes malignancy of cancer cells via CCL20. Tumour Biol. 2016, 37, 1995–2005. [Google Scholar] [CrossRef]
- Chuang, H.C.; Chou, M.H.; Chien, C.Y.; Chuang, J.H.; Liu, Y.L. Triggering TLR3 pathway promotes tumor growth and cisplatin resistance in head and neck cancer cells. Oral. Oncol. 2018, 86, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.N.; Holsinger, F.C.; Jasser, S.A.; Bekele, B.N.; Fidler, I.J. An orthotopic nude mouse model of oral tongue squamous cell carcinoma. Clin. Cancer Res. 2002, 8, 293–298. [Google Scholar] [PubMed]
- Haddad, R.I.; Posner, M.; Hitt, R.; Cohen, E.E.W.; Schulten, J.; Lefebvre, J.L.; Vermorken, J.B. Induction chemotherapy in locally advanced squamous cell carcinoma of the head and neck: Role, controversy, and future directions. Ann. Oncol. 2018, 29, 1130–1140. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Kuriyama, H.; Umezu, H.; Tanaka, J.; Yoshimasu, T.; Furukawa, T.; Tanaka, H.; Kagamu, H.; Gejyo, F.; Yoshizawa, H. Class III beta-tubulin expression in tumor cells is correlated with resistance to docetaxel in patients with completely resected non-small-cell lung cancer. Intern. Med. 2009, 48, 203–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, S.A.; Hicks, D.G.; Watroba, N.L.; Murekeyisoni, C.; Hwang, H.; Khoury, T.; Beck, R.A.; Ring, B.Z.; Estopinal, N.C.; Schreeder, M.T.; et al. TLE3 as a candidate biomarker of response to taxane therapy. Breast Cancer Res. 2009, 11, R17. [Google Scholar] [CrossRef] [Green Version]
- Molinolo, A.A.; Hewitt, S.M.; Amornphimoltham, P.; Keelawat, S.; Rangdaeng, S.; Meneses Garcia, A.; Raimondi, A.R.; Jufe, R.; Itoiz, M.; Gao, Y.; et al. Dissecting the Akt/mammalian target of rapamycin signaling network: Emerging results from the head and neck cancer tissue array initiative. Clin. Cancer Res. 2007, 13, 4964–4973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lionello, M.; Blandamura, S.; Loreggian, L.; Ottaviano, G.; Giacomelli, L.; Marchese-Ragona, R.; Velardita, C.; Staffieri, A.; Marioni, G. High mTOR expression is associated with a worse oncological outcome in laryngeal carcinoma treated with postoperative radiotherapy: A pilot study. J. Oral. Pathol. Med. 2012, 41, 136–140. [Google Scholar] [CrossRef]
- Zaytseva, Y.Y.; Valentino, J.D.; Gulhati, P.; Evers, B.M. mTOR inhibitors in cancer therapy. Cancer Lett. 2012, 319, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.A.; Walker, D.; Gillespie, M.B.; Gutkind, J.S.; Day, T.A. mTOR inhibitors and its role in the treatment of head and neck squamous cell carcinoma. Curr. Treat. Opt. Oncol. 2012, 13, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Day, T.A.; Shirai, K.; O’Brien, P.E.; Matheus, M.G.; Godwin, K.; Sood, A.J.; Kompelli, A.; Vick, J.A.; Martin, D.; Vitale-Cross, L.; et al. Inhibition of mTOR Signaling and Clinical Activity of Rapamycin in Head and Neck Cancer in a Window of Opportunity Trial. Clin. Cancer Res. 2019, 25, 1156–1164. [Google Scholar] [CrossRef] [Green Version]
- Kozakiewicz, P.; Grzybowska-Szatkowska, L. Application of molecular targeted therapies in the treatment of head and neck squamous cell carcinoma. Oncol. Lett. 2018, 15, 7497–7505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Li, J.Z.; Chan, J.Y.; Ho, W.K.; Wong, T.S. mTOR Pathway and mTOR Inhibitors in Head and Neck Cancer. ISRN Otolaryngol. 2012, 2012, 953089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Luo, Y.; Huang, S. Updates of mTOR inhibitors. Anticancer Agents Med. Chem. 2010, 10, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Ekshyyan, O.; Moore-Medlin, T.N.; Raley, M.C.; Sonavane, K.; Rong, X.; Brodt, M.A.; Abreo, F.; Alexander, J.S.; Nathan, C.A. Anti-lymphangiogenic properties of mTOR inhibitors in head and neck squamous cell carcinoma experimental models. BMC Cancer 2013, 13, 320. [Google Scholar] [CrossRef] [Green Version]
- Pal, I.; Mandal, M. PI3K and Akt as molecular targets for cancer therapy: Current clinical outcomes. Acta Pharmacol. Sin. 2012, 33, 1441–1458. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, N.A.; McDonald, K.; Tong, L.; von Euw, E.; Kalous, O.; Conklin, D.; Hurvitz, S.A.; di Tomaso, E.; Schnell, C.; Linnartz, R.; et al. Targeting PI3K/mTOR overcomes resistance to HER2-targeted therapy independent of feedback activation of AKT. Clin. Cancer Res. 2014, 20, 3507–3520. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yao, L.; Yang, J.; Wang, Z.; Du, G. PI3K/Akt and HIF1 signaling pathway in hypoxiaischemia (Review). Mol. Med. Rep. 2018, 18, 3547–3554. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.M.; Wang, Y.S.; Zhang, J.; Li, Y.; Xu, J.F.; Zhu, J.; Zhao, W.; Chu, D.K.; Wiedemann, P. Role of PI3K/Akt and MEK/ERK in mediating hypoxia-induced expression of HIF-1alpha and VEGF in laser-induced rat choroidal neovascularization. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Karar, J.; Cerniglia, G.J.; Lindsten, T.; Koumenis, C.; Maity, A. Dual PI3K/mTOR inhibitor NVP-BEZ235 suppresses hypoxia-inducible factor (HIF)-1alpha expression by blocking protein translation and increases cell death under hypoxia. Cancer Biol. Ther. 2012, 13, 1102–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, H.; Jia, B.; Qiu, X.; Pan, J.; Sun, X.; Wang, Z.; Zhao, J. Investigation of proliferation and migration of tongue squamous cell carcinoma promoted by three chemokines, MIP-3alpha, MIP-1beta, and IP-10. Onco Targets Ther. 2017, 10, 4193–4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, M.-H.; Chuang, H.-C.; Lin, Y.-T.; Tsai, M.-H.; Kao, Y.-H.; Lin, I.-C.; Huang, T.-L.; Fang, F.-M.; Chien, C.-Y. Targeting mTOR-CCL20 Signaling May Improve Response to Docetaxel in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 3046. https://doi.org/10.3390/ijms22063046
Chou M-H, Chuang H-C, Lin Y-T, Tsai M-H, Kao Y-H, Lin I-C, Huang T-L, Fang F-M, Chien C-Y. Targeting mTOR-CCL20 Signaling May Improve Response to Docetaxel in Head and Neck Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2021; 22(6):3046. https://doi.org/10.3390/ijms22063046
Chicago/Turabian StyleChou, Ming-Huei, Hui-Ching Chuang, Yu-Tsai Lin, Ming-Hsien Tsai, Ying-Hsien Kao, I-Chun Lin, Tai-Lin Huang, Fu-Min Fang, and Chih-Yen Chien. 2021. "Targeting mTOR-CCL20 Signaling May Improve Response to Docetaxel in Head and Neck Squamous Cell Carcinoma" International Journal of Molecular Sciences 22, no. 6: 3046. https://doi.org/10.3390/ijms22063046
APA StyleChou, M.-H., Chuang, H.-C., Lin, Y.-T., Tsai, M.-H., Kao, Y.-H., Lin, I.-C., Huang, T.-L., Fang, F.-M., & Chien, C.-Y. (2021). Targeting mTOR-CCL20 Signaling May Improve Response to Docetaxel in Head and Neck Squamous Cell Carcinoma. International Journal of Molecular Sciences, 22(6), 3046. https://doi.org/10.3390/ijms22063046