
Distinct Regulation of Dopamine D3 Receptor in the Basolateral Amygdala and Dentate Gyrus during the Reinstatement of Cocaine CPP Induced by Drug Priming and Social Stress

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
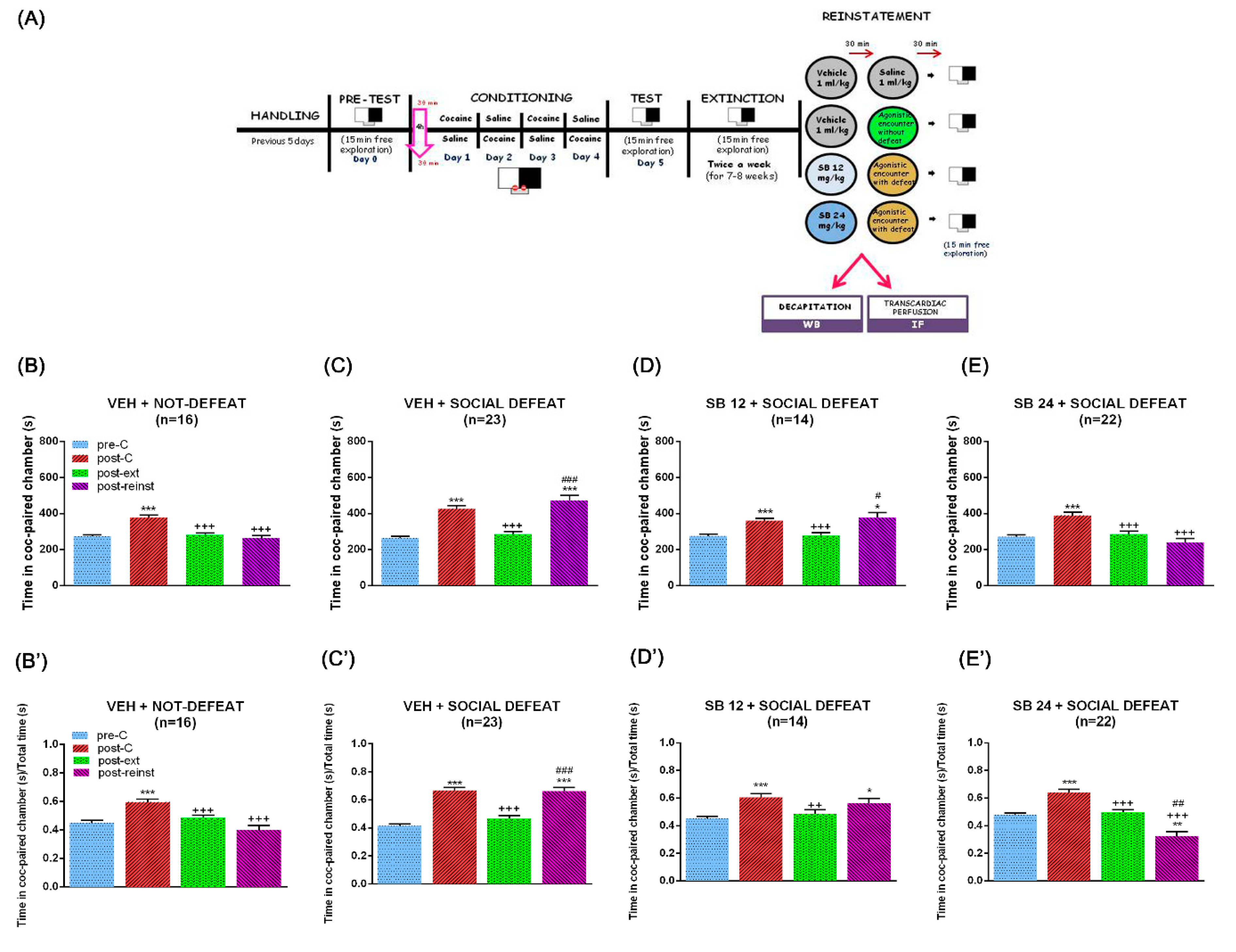
2.1. D3 Antagonism Did Not Prevent the Reinstatement of the CPP Induced by a Cocaine Prime but Did Block the Reinstatement of the CPP Induced by Social Defeat
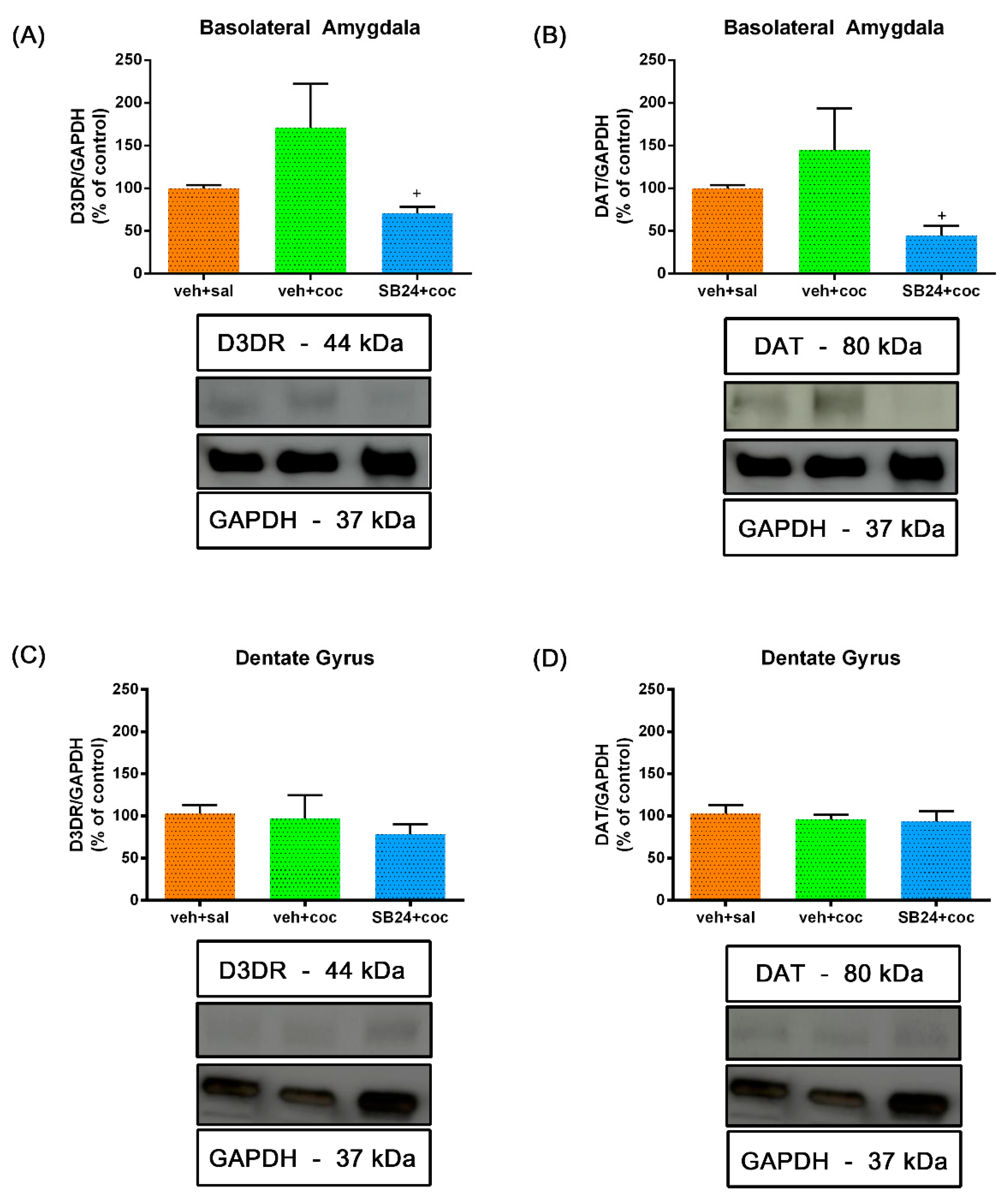
2.2. The Administration of SB-277011-A Prior Drug Prime and Social Stress Altered D3R and DAT Expression in the BLA but Not in the Hippocampal DG
2.3. The Reinstatement of Cocaine CPP Induced by Both a Drug Prime and Social Stress Decreased mTOR Phosphorylation Ratio in BLA and DG
2.4. Characterization of D3R-Expressing Neurons
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Reagents
4.3. Conditioned Place Preference Paradigm
4.4. Experimental Group
4.4.1. Experiment 1: Effect of D3R Antagonism on the Reinstatement of Cocaine-Induced CPP Evoked by a Cocaine Prime
4.4.2. Experiment 2: Effect of D3R Antagonism on the Reinstatement of Cocaine-Induced CPP Evoked by Social Stress
4.5. Tissue Collection and Western Blot Analysis
4.6. Immunofluorescence
4.7. Confocal Analysis
4.8. Data Collection and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mantsch, J.R.; Baker, D.A.; Funk, D.; Lê, A.D.; Shaham, Y. Stress-Induced Reinstatement of Drug Seeking: 20 Years of Progress. Neuropsychopharmacology 2016, 41, 335–356. [Google Scholar] [CrossRef]
- Farrell, M.R.; Schoch, H.; Mahler, S.V. Modeling cocaine relapse in rodents: Behavioral considerations and circuit mechanisms. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 87, 33–47. [Google Scholar] [CrossRef]
- Dong, Y.; Taylor, J.R.; Wolf, M.E.; Shaham, Y. Circuit and Synaptic Plasticity Mechanisms of Drug Relapse. J. Neurosci. 2017, 37, 10867–10876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchis-Segura, C.; Spanagel, R. Review: Behavioural assessment of drug reinforcement and addictive features in rodents: An overview. Addict. Biol. 2006, 11, 2–38. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, D.H.; Brown, R.; Lawrence, A.J.; Rezvani, A.H. Chapter 16—Overview of Animal Models of Drug Addiction: Commonalities to Human Addiction. In Biological Research on Addiction; Miller, P.M., Ed.; Academic Press: San Diego, CA, USA, 2013; pp. 149–157. [Google Scholar] [CrossRef]
- Tzschentke, T.M. Review on CPP: Measuring reward with the conditioned place preference (CPP) paradigm: Update of the last decade. Addict. Biol. 2007, 12, 227–462. [Google Scholar] [CrossRef]
- Rutten, K.; Rutten, K.; van der Kam, E.L.; De Vry, J.; Tzschentke, T.M. Critical Evaluation of the Use of Extinction Paradigms for the Assessment of Opioid-Induced Conditioned Place Preference in Rats. Pharmacology 2011, 87, 286–296. [Google Scholar] [CrossRef]
- Cunningham, C.L.; Gremel, C.M.; Groblewski, P.A. Drug-induced conditioned place preference and aversion in mice. Nat. Protoc. 2006, 1, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Kelley, A.E. Memory and Addiction: Shared Neural Circuitry and Molecular Mechanisms. Neuron 2004, 44, 161–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torregrossa, M.M.; Corlett, P.R.; Taylor, J.R. Aberrant learning and memory in addiction. Neurobiol. Learn. Mem. 2011, 96, 609–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hainmueller, T.; Bartos, M. Dentate gyrus circuits for encoding, retrieval and discrimination of episodic memories. Nat. Rev. Neurosci. 2020, 21, 153–168. [Google Scholar] [CrossRef]
- Garcia-Perez, D.; Ferenczi, S.; Kovacs, K.J.; Laorden, M.L.; Milanes, M.V.; Nunez, C. Glucocorticoid Homeostasis in the Dentate Gyrus Is Essential for Opiate Withdrawal-Associated Memories. Mol. Neurobiol. 2017, 54, 6523–6541. [Google Scholar] [CrossRef]
- Goodman, J.; Packard, M.G. Memory Systems and the Addicted Brain. Front. Psychiatry 2016, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Nestler, E.J. Cellular basis of memory for addiction. Dialogues Clin. Neurosci. 2013, 15, 431–443. [Google Scholar] [PubMed]
- García-Pérez, D.; Ferenczi, S.; Kovács, K.J.; Laorden, M.L.; Milanés, M.V.; Núñez, C. Different contribution of glucocorticoids in the basolateral amygdala to the formation and expression of opiate withdrawal-associated memories. Psychoneuroendocrinology 2016, 74, 350–362. [Google Scholar] [CrossRef]
- Belin, D.; Belin-Rauscent, A.; Murray, J.E.; Everitt, B.J. Addiction: Failure of control over maladaptive incentive habits. Curr. Opin. Neurobiol. 2013, 23, 564–572. [Google Scholar] [CrossRef]
- Rosen, L.G.; Rushlow, W.J.; Laviolette, S.R. Opiate exposure state controls dopamine D3 receptor and cdk5/calcineurin signaling in the basolateral amygdala during reward and withdrawal aversion memory formation. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2017, 79, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Sokoloff, P.; Le Foll, B. The dopamine D3 receptor, a quarter century later. Eur. J. Neurosci. 2017, 45, 2–19. [Google Scholar] [CrossRef] [PubMed]
- Takashima, Y.; Tseng, J.; Fannon, M.J.; Purohit, D.C.; Quach, L.W.; Terranova, M.J.; Kharidia, K.M.; Oliver, R.J.; Mandyam, C.D. Sex Differences in Context-Driven Reinstatement of Methamphetamine Seeking is Associated with Distinct Neuroadaptations in the Dentate Gyrus. Brain Sci. 2018, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Torres, G.E. The dopamine transporter proteome. J. Neurochem. 2006, 97, 3–10. [Google Scholar] [CrossRef]
- Dayas, C.V.; Smith, D.W.; Dunkley, P.R. An emerging role for the Mammalian target of rapamycin in "pathological" protein translation: Relevance to cocaine addiction. Front. Pharmacol. 2012, 3, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neasta, J.; Barak, S.; Hamida, S.B.; Ron, D. mTOR complex 1: A key player in neuroadaptations induced by drugs of abuse. J. Neurochem. 2014, 130, 172–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeffer, C.A.; Klann, E. mTOR signaling: At the crossroads of plasticity, memory and disease. Trends Neurosci. 2010, 33, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Bautista, R.; Do Couto, B.R.; Hidalgo, J.M.; Cárceles-Moreno, F.J.; Molina, G.; Laorden, M.L.; Núñez, C.; Milanés, M.V. Modulation of stress—And cocaine prime-induced reinstatement of conditioned place preference after memory extinction through dopamine D3 receptor. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2019, 92, 308–320. [Google Scholar] [CrossRef]
- Guerrero-Bautista, R.; Franco-García, A.; Hidalgo, J.M.; Fernández-Gómez, F.; Milanés, M.V.; Núñez, C. Blockade of D3 receptor prevents changes in DAT and D3R expression in the mesolimbic dopaminergic circuit produced by social stress—and cocaine prime-induced reinstatement of cocaine-CPP. J. Psychopharmacol. 2020, 34, 1300–1315. [Google Scholar] [CrossRef] [PubMed]
- Solinas, M.; Belujon, P.; Fernagut, P.O.; Jaber, M.; Thiriet, N. Dopamine and addiction: What have we learned from 40 years of research. J. Neural Transm. 2019, 126, 481–516. [Google Scholar] [CrossRef]
- Roozendaal, B.; McEwen, B.S.; Chattarji, S. Stress, memory and the amygdala. Nat. Rev. Neurosci. 2009, 10, 423–433. [Google Scholar] [CrossRef]
- Lynch, W.J. Modeling the development of drug addiction in male and female animals. Pharmacol. Biochem. Behav. 2018, 164, 50–61. [Google Scholar] [CrossRef]
- Steketee, J.D.; Kalivas, P.W. Drug Wanting: Behavioral Sensitization and Relapse to Drug-Seeking Behavior. Pharmacol. Rev. 2011, 63, 348–365. [Google Scholar] [CrossRef]
- Montagud-Romero, S.; Blanco-Gandía, M.C.; Reguilón, M.D.; Ferrer-Pérez, C.; Ballestín, R.; Miñarro, J.; Rodríguez-Arias, M. Social defeat stress: Mechanisms underlying the increase in rewarding effects of drugs of abuse. Eur. J. Neurosci. 2018, 48, 2948–2970. [Google Scholar] [CrossRef]
- Galaj, E.; Ewing, S.; Ranaldi, R. Dopamine D1 and D3 receptor polypharmacology as a potential treatment approach for substance use disorder. Neurosci. Biobehav. Rev. 2018, 89, 13–28. [Google Scholar] [CrossRef]
- Ashby, C.R., Jr.; Rice, O.V.; Heidbreder, C.A.; Gardner, E.L. The selective dopamine D3 receptor antagonist SB-277011A attenuates drug—Or food-deprivation reactivation of expression of conditioned place preference for cocaine in male sprague-dawley rats. Synapse 2015, 69, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.-X.; Gilbert, J.; Campos, A.C.; Kline, N.; Ashby, C.R.; Hagan, J.J.; Heidbreder, C.A.; Gardner, E.L. Blockade of mesolimbic dopamine D3 receptors inhibits stress-induced reinstatement of cocaine-seeking in rats. Psychopharmacology 2004, 176, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higley, A.E.; Kiefer, S.W.; Li, X.; Gaál, J.; Xi, Z.-X.; Gardner, E.L. Dopamine D(3) receptor antagonist SB-277011A inhibits methamphetamine self-administration and methamphetamine-induced reinstatement of drug-seeking in rats. Eur. J. Pharmacol. 2011, 659, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreoli, M.; Tessari, M.; Pilla, M.; Valerio, E.; Hagan, J.J.; Heidbreder, C.A. Selective Antagonism at Dopamine D3 Receptors Prevents Nicotine-Triggered Relapse to Nicotine-Seeking Behavior. Neuropsychopharmacology 2003, 28, 1272–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vengeliene, V.; Leonardi-Essmann, F.; Perreau-Lenz, S.; Gebicke-Haerter, P.; Drescher, K.; Gross, G.; Spanagel, R. The dopamine D3 receptor plays an essential role in alcohol-seeking and relapse. FASEB J. 2006, 20, 2223–2233. [Google Scholar] [CrossRef]
- Rice, O.V.; Heidbreder, C.A.; Gardner, E.L.; Schonhar, C.D.; Ashby, C.R., Jr. The selective D₃ receptor antagonist SB-277011A attenuates morphine-triggered reactivation of expression of cocaine-induced conditioned place preference. Synapse 2013, 67, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.-X.; Newman, A.H.; Gilbert, J.G.; Pak, A.C.; Peng, X.-Q.; Ashby, C.R.; Gitajn, L.; Gardner, E.L. The Novel Dopamine D3 Receptor Antagonist NGB 2904 Inhibits Cocaine’s Rewarding Effects and Cocaine-Induced Reinstatement of Drug-Seeking Behavior in Rats. Neuropsychopharmacology 2006, 31, 1393–1405. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.-X.; Gilbert, J.G.; Pak, A.C.; Ashby, C.R., Jr.; Heidbreder, C.A.; Gardner, E.L. Selective dopamine D3 receptor antagonism by SB-277011A attenuates cocaine reinforcement as assessed by progressive-ratio and variable-cost–variable-payoff fixed-ratio cocaine self-administration in rats. Eur. J. Neurosci. 2005, 21, 3427–3438. [Google Scholar] [CrossRef] [Green Version]
- Sokoloff, P.; Giros, B.; Martres, M.-P.; Bouthenet, M.-L.; Schwartz, J.-C. Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 1990, 347, 146–151. [Google Scholar] [CrossRef]
- Everitt, B.J.; Robbins, T.W. Neural systems of reinforcement for drug addiction: From actions to habits to compulsion. Nat. Neurosci. 2005, 8, 1481–1489. [Google Scholar] [CrossRef]
- Staley, J.K.; Mash, D.C. Adaptive Increase in D3 Dopamine Receptors in the Brain Reward Circuits of Human Cocaine Fatalities. J. Neurosci. 1996, 16, 6100–6106. [Google Scholar] [CrossRef] [Green Version]
- Le Foll, B.; Francès, H.; Diaz, J.; Schwartz, J.-C.; Sokoloff, P. Role of the dopamine D3 receptor in reactivity to cocaine-associated cues in mice. Eur. J. Neurosci. 2002, 15, 2016–2026. [Google Scholar] [CrossRef]
- Chen, P.C.; Lao, C.L.; Chen, J.C. The D(3) dopamine receptor inhibits dopamine release in PC-12/hD3 cells by autoreceptor signaling via PP-2B, CK1, and Cdk-5. J. Neurochem. 2009, 110, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Grace, A.A. The tonic/phasic model of dopamine system regulation and its implications for understanding alcohol and psychostimulant craving. Addiction 2000, 95, 119–128. [Google Scholar] [CrossRef]
- Berton, O.; McClung, C.A.; Dileone, R.J.; Krishnan, V.; Renthal, W.; Russo, S.J.; Graham, D.; Tsankova, N.M.; Bolanos, C.A.; Rios, M.; et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 2006, 311, 864–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holly, E.N.; DeBold, J.F.; Miczek, K.A. Increased mesocorticolimbic dopamine during acute and repeated social defeat stress: Modulation by corticotropin releasing factor receptors in the ventral tegmental area. Psychopharmacology 2015, 232, 4469–4479. [Google Scholar] [CrossRef]
- Tidey, J.W.; Miczek, K.A. Social defeat stress selectively alters mesocorticolimbic dopamine release: An in vivo microdialysis study. Brain Res. 1996, 721, 140–149. [Google Scholar] [CrossRef]
- Nygard, S.K.; Hourguettes, N.J.; Sobczak, G.G.; Carlezon, W.A.; Bruchas, M.R. Stress-Induced Reinstatement of Nicotine Preference Requires Dynorphin/Kappa Opioid Activity in the Basolateral Amygdala. J. Neurosci. 2016, 36, 9937–9948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokoloff, P.; Diaz, J.; Le Foll, B.; Guillin, O.; Leriche, L.; Bezard, E.; Gross, C. The dopamine D3 receptor: A therapeutic target for the treatment of neuropsychiatric disorders. CNS Neurol. Disord. Drug Targets 2006, 5, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Twining, R.C.; Wheeler, D.S.; Ebben, A.L.; Jacobsen, A.J.; Robble, M.A.; Mantsch, J.R.; Wheeler, R.A. Aversive stimuli drive drug seeking in a state of low dopamine tone. Biol. Psychiatry 2015, 77, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Urban, N.B.; Martinez, D. Neurobiology of addiction: Insight from neurochemical imaging. Psychiatr. Clin. 2012, 35, 521–541. [Google Scholar] [CrossRef]
- Izenwasser, S. The role of the dopamine transporter in cocaine abuse. Neurotox. Res. 2004, 6, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Zapata, A.; Kivell, B.; Han, Y.; Javitch, J.A.; Bolan, E.A.; Kuraguntla, D.; Jaligam, V.; Oz, M.; Jayanthi, L.D.; Samuvel, D.J.; et al. Regulation of dopamine transporter function and cell surface expression by D3 dopamine receptors. J. Biol. Chem. 2007, 282, 35842–35854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, A.; Shippenberg, T.S. D3 receptor ligands modulate extracellular dopamine clearance in the nucleus accumbens. J. Neurochem. 2002, 81, 1035–1042. [Google Scholar] [CrossRef]
- Castro-Hernández, J.; Afonso-Oramas, D.; Cruz-Muros, I.; Salas-Hernández, J.; Barroso-Chinea, P.; Moratalla, R.; Millan, M.J.; González-Hernández, T. Prolonged treatment with pramipexole promotes physical interaction of striatal dopamine D3 autoreceptors with dopamine transporters to reduce dopamine uptake. Neurobiol. Dis. 2015, 74, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Seeman, P.; Lee, T.; Chau-Wong, M.; Wong, K. Antipsychotic drug doses and neuroleptic/dopamine receptors. Nature 1976, 261, 717–719. [Google Scholar] [CrossRef]
- Hess, E.; Norman, A.; Creese, I. Chronic treatment with dopamine receptor antagonists: Behavioral and pharmacologic effects on D1 and D2 dopamine receptors. J. Neurosci. 1988, 8, 2361–2370. [Google Scholar] [CrossRef]
- Tzeng, W.Y.; Chuang, J.Y.; Lin, L.C.; Cherng, C.G.; Lin, K.Y.; Chen, L.H.; Su, C.C.; Yu, L. Companions reverse stressor-induced decreases in neurogenesis and cocaine conditioning possibly by restoring BDNF and NGF levels in dentate gyrus. Psychoneuroendocrinology 2013, 38, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Takashima, Y.; Fannon, M.J.; Galinato, M.H.; Steiner, N.L.; An, M.; Zemljic-Harpf, A.E.; Somkuwar, S.S.; Head, B.P.; Mandyam, C.D. Neuroadaptations in the dentate gyrus following contextual cued reinstatement of methamphetamine seeking. Brain Struct. Funct. 2018, 223, 2197–2211. [Google Scholar] [CrossRef] [PubMed]
- Pourhamzeh, M.; Mozafari, R.; Jamali, S.; Motamedi, F.; Ahadi, R.; Haghparast, A. Involvement of orexin receptors within the hippocampal dentate gyrus in morphine-induced reinstatement in food-deprived rats. Behav. Brain Res. 2019, 375, 112155. [Google Scholar] [CrossRef]
- Ebrahimian, F.; Naghavi, F.S.; Yazdi, F.; Sadeghzadeh, F.; Taslimi, Z.; Haghparast, A. Differential roles of orexin receptors within the dentate gyrus in stress- and drug priming-induced reinstatement of conditioned place preference in rats. Behav. Neurosci. 2016, 130, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Khakpour-Taleghani, B.; Reisi, Z.; Haghparast, A. The Blockade of D1/D2-Like Dopamine Receptors within the Dentate Gyrus of Hippocampus Decreased the Reinstatement of Morphine-Extinguished Conditioned Place Preference in Rats. Basic Clin. Neurosci. 2015, 6, 73–82. [Google Scholar] [PubMed]
- Norozpour, Y.; Zarrabian, S.; Rezaee, L.; Haghparast, A. D1- and D2-like receptors in the dentate gyrus region of the hippocampus are involved in the reinstatement induced by a subthreshold dose of morphine and forced swim stress in extinguished morphine-CPP in rats. Behav. Neurosci. 2019, 133, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Sutton, L.P.; Caron, M.G. Essential role of D1R in the regulation of mTOR complex1 signaling induced by cocaine. Neuropharmacology 2015, 99, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Chiamulera, C.; Di Chio, M.; Cavalleri, L.; Venniro, M.; Padovani, L.; Collo, G. Ketamine effects on mammalian target of rapamycin signaling in the mouse limbic system depend on functional dopamine D3 receptors. Neuroreport 2018, 29, 615–620. [Google Scholar] [CrossRef]
- Barroso-Chinea, P.; Luis-Ravelo, D.; Fumagallo-Reading, F.; Castro-Hernandez, J.; Salas-Hernandez, J.; Rodriguez-Nuñez, J.; Febles-Casquero, A.; Cruz-Muros, I.; Afonso-Oramas, D.; Abreu-Gonzalez, P.; et al. DRD3 (dopamine receptor D3) but not DRD2 activates autophagy through MTORC1 inhibition preserving protein synthesis. Autophagy 2020, 16, 1279–1295. [Google Scholar] [CrossRef] [PubMed]
- Martel, J.C.; Gatti McArthur, S. Dopamine Receptor Subtypes, Physiology and Pharmacology: New Ligands and Concepts in Schizophrenia. Front. Pharmacol. 2020, 11, 1003. [Google Scholar] [CrossRef]
- Puighermanal, E.; Marsicano, G.; Busquets-Garcia, A.; Lutz, B.; Maldonado, R.; Ozaita, A. Cannabinoid modulation of hippocampal long-term memory is mediated by mTOR signaling. Nat. Neurosci. 2009, 12, 1152–1158. [Google Scholar] [CrossRef]
- Puighermanal, E.; Busquets-Garcia, A.; Gomis-González, M.; Marsicano, G.; Maldonado, R.; Ozaita, A. Dissociation of the pharmacological effects of THC by mTOR blockade. Neuropsychopharmacology 2013, 38, 1334–1343. [Google Scholar] [CrossRef] [Green Version]
- Neasta, J.; Ben Hamida, S.; Yowell, Q.; Carnicella, S.; Ron, D. Role for mammalian target of rapamycin complex 1 signaling in neuroadaptations underlying alcohol-related disorders. Proc. Natl. Acad. Sci. USA 2010, 107, 20093–20098. [Google Scholar] [CrossRef] [Green Version]
- Collo, G.; Bono, F.; Cavalleri, L.; Plebani, L.; Mitola, S.; Merlo Pich, E.; Millan, M.J.; Zoli, M.; Maskos, U.; Spano, P.; et al. Nicotine-Induced Structural Plasticity in Mesencephalic Dopaminergic Neurons Is Mediated by Dopamine D3 Receptors and Akt-mTORC1 Signaling. Mol. Pharmacol. 2013, 83, 1176–1189. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, J.; Baptista, S.; Olesen, M.V.; Fontes-Ribeiro, C.; Malva, J.O.; Woldbye, D.P.; Silva, A.P. Methamphetamine-induced changes in the mice hippocampal neuropeptide Y system: Implications for memory impairment. J. Neurochem. 2012, 123, 1041–1053. [Google Scholar] [CrossRef]
- Ben Hamida, S.; Laguesse, S.; Morisot, N.; Park, J.-H.; Phuamluong, K.; Berger, A.L.; Park, K.D.; Ron, D. Mammalian target of rapamycin complex 1 and its downstream effector collapsin response mediator protein-2 drive reinstatement of alcohol reward seeking. Addict. Biol. 2019, 24, 908–920. [Google Scholar] [CrossRef]
- Ucha, M.; Coria, S.M.; Núñez, A.E.; Santos-Toscano, R.; Roura-Martínez, D.; Fernández-Ruiz, J.; Higuera-Matas, A.; Ambrosio, E. Morphine self-administration alters the expression of translational machinery genes in the amygdala of male Lewis rats. J. Psychopharmacol. 2019, 33, 882–893. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, X.Q.; Cui, Y.; Xin, W.J.; Jing, J.; Liu, X.G. Activation of phosphatidylinositol 3-kinase/Akt-mammalian target of Rapamycin signaling pathway in the hippocampus is essential for the acquisition of morphine-induced place preference in rats. Neuroscience 2010, 171, 134–143. [Google Scholar] [CrossRef]
- Deli, A.; Schipany, K.; Rosner, M.; Höger, H.; Pollak, A.; Li, L.; Hengstschläger, M.; Lubec, G. Blocking mTORC1 activity by rapamycin leads to impairment of spatial memory retrieval but not acquisition in C57BL/6J mice. Behav. Brain Res. 2012, 229, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Al-Ali, H.; Ding, Y.; Slepak, T.; Wu, W.; Sun, Y.; Martinez, Y.; Xu, X.M.; Lemmon, V.P.; Bixby, J.L. The mTOR Substrate S6 Kinase 1 (S6K1) Is a Negative Regulator of Axon Regeneration and a Potential Drug Target for Central Nervous System Injury. J. Neurosci. 2017, 37, 7079–7095. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, Y.; Tran, J.; O’Neal, P.; Baudry, M.; Bi, X. mTORC1-S6K1 inhibition or mTORC2 activation improves hippocampal synaptic plasticity and learning in Angelman syndrome mice. Cell. Mol. Life Sci. 2016, 73, 4303–4314. [Google Scholar] [CrossRef]
- Rodriguez-Arias, M.; Minarro, J.; Aguilar, M.A.; Pinazo, J.; Simon, V.M. Effects of risperidone and SCH 23390 on isolation-induced aggression in male mice. Eur. Neuropsychopharmacol. 1998, 8, 95–103. [Google Scholar] [CrossRef]
- Valverde, O.; Fournie-Zaluski, M.C.; Roques, B.P.; Maldonado, R. The CCKB antagonist PD-134,308 facilitates rewarding effects of endogenous enkephalins but does not induce place preference in rats. Psychopharmacology 1996, 123, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, C.; Rodriguez-Arias, M.; Castillo, A.; Aguilar, M.A.; Minarro, J. Effect of memantine and CNQX in the acquisition, expression and reinstatement of cocaine-induced conditioned place preference. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2007, 31, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Arias, M.; Montagud-Romero, S.; Rubio-Araiz, A.; Aguilar, M.A.; Martin-Garcia, E.; Cabrera, R.; Maldonado, R.; Porcu, F.; Colado, M.I.; Minarro, J. Effects of repeated social defeat on adolescent mice on cocaine-induced CPP and self-administration in adulthood: Integrity of the blood-brain barrier. Addict. Biol. 2017, 22, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Arias, M.; Castillo, A.; Daza-Losada, M.; Aguilar, M.A.; Minarro, J. Effects of extended cocaine conditioning in the reinstatement of place preference. Physiol. Behav. 2009, 96, 620–630. [Google Scholar] [CrossRef]
- Ribeiro Do Couto, B.; Aguilar, M.A.; Lluch, J.; Rodriguez-Arias, M.; Minarro, J. Social experiences affect reinstatement of cocaine-induced place preference in mice. Psychopharmacology 2009, 207, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Miczek, K.A.; Nikulina, E.M.; Shimamoto, A.; Covington, H.E., 3rd. Escalated or suppressed cocaine reward, tegmental BDNF, and accumbal dopamine caused by episodic versus continuous social stress in rats. J. Neurosci. 2011, 31, 9848–9857. [Google Scholar] [CrossRef] [Green Version]
- Miczek, K.A.; Thompson, M.L.; Shuster, L. Opioid-like analgesia in defeated mice. Science 1982, 215, 1520–1522. [Google Scholar] [CrossRef]
- Frankin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: San Diego, CA, USA, 2008. [Google Scholar]
- Leng, A.; Feldon, J.; Ferger, B. Long-term social isolation and medial prefrontal cortex: Dopaminergic and cholinergic neurotransmission. Pharmacol. Biochem. Behav. 2004, 77, 371–379. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerrero-Bautista, R.; Franco-García, A.; Hidalgo, J.M.; Fernández-Gómez, F.J.; Ribeiro Do Couto, B.; Milanés, M.V.; Núñez, C. Distinct Regulation of Dopamine D3 Receptor in the Basolateral Amygdala and Dentate Gyrus during the Reinstatement of Cocaine CPP Induced by Drug Priming and Social Stress. Int. J. Mol. Sci. 2021, 22, 3100. https://doi.org/10.3390/ijms22063100
Guerrero-Bautista R, Franco-García A, Hidalgo JM, Fernández-Gómez FJ, Ribeiro Do Couto B, Milanés MV, Núñez C. Distinct Regulation of Dopamine D3 Receptor in the Basolateral Amygdala and Dentate Gyrus during the Reinstatement of Cocaine CPP Induced by Drug Priming and Social Stress. International Journal of Molecular Sciences. 2021; 22(6):3100. https://doi.org/10.3390/ijms22063100
Chicago/Turabian StyleGuerrero-Bautista, Rocío, Aurelio Franco-García, Juana M. Hidalgo, Francisco José Fernández-Gómez, Bruno Ribeiro Do Couto, M. Victoria Milanés, and Cristina Núñez. 2021. "Distinct Regulation of Dopamine D3 Receptor in the Basolateral Amygdala and Dentate Gyrus during the Reinstatement of Cocaine CPP Induced by Drug Priming and Social Stress" International Journal of Molecular Sciences 22, no. 6: 3100. https://doi.org/10.3390/ijms22063100