
Sesquiterpene Lactone Deoxyelephantopin Isolated from Elephantopus scaber and Its Derivative DETD-35 Suppress BRAFV600E Mutant Melanoma Lung Metastasis in Mice

 , and
, and 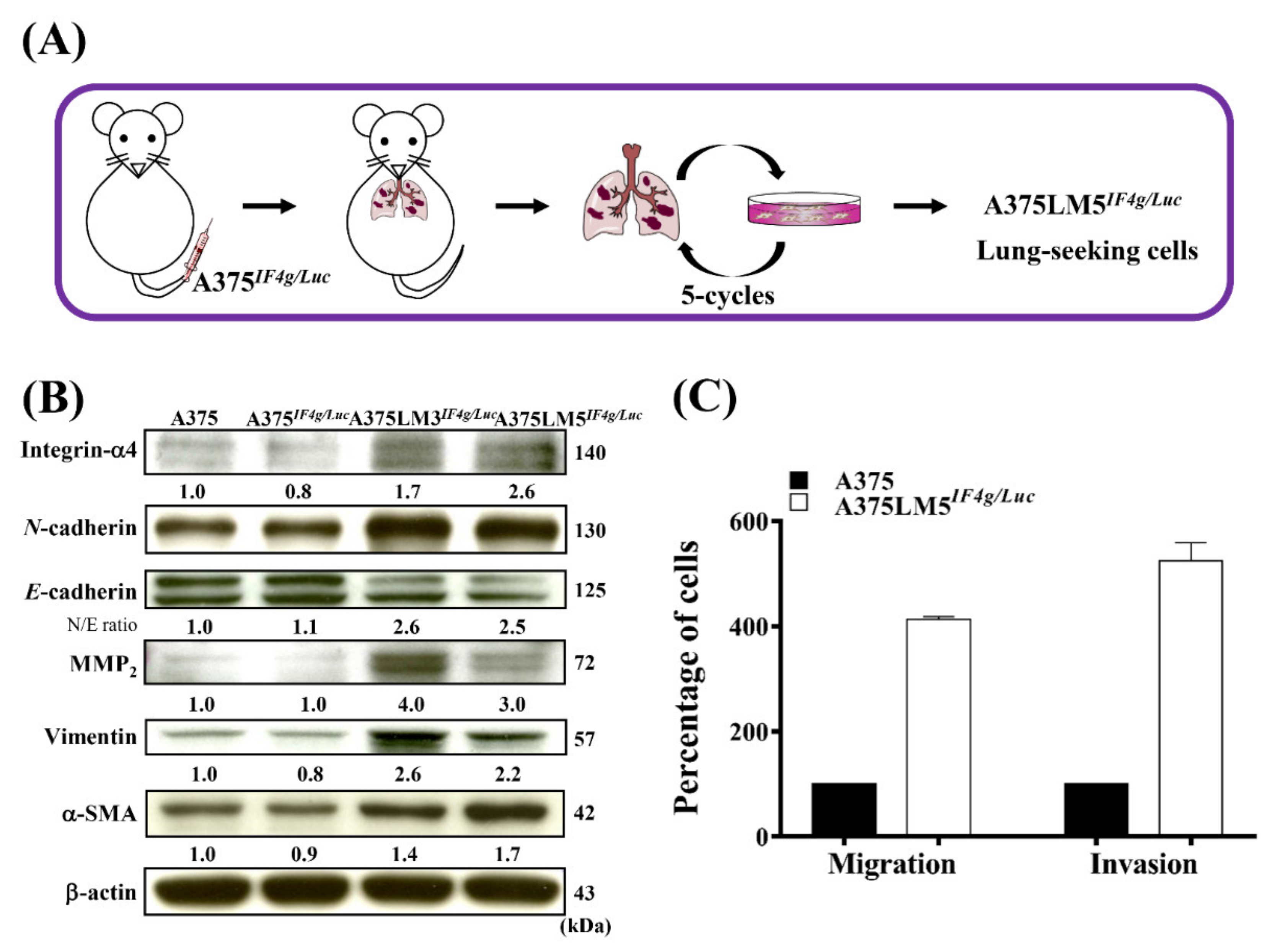{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
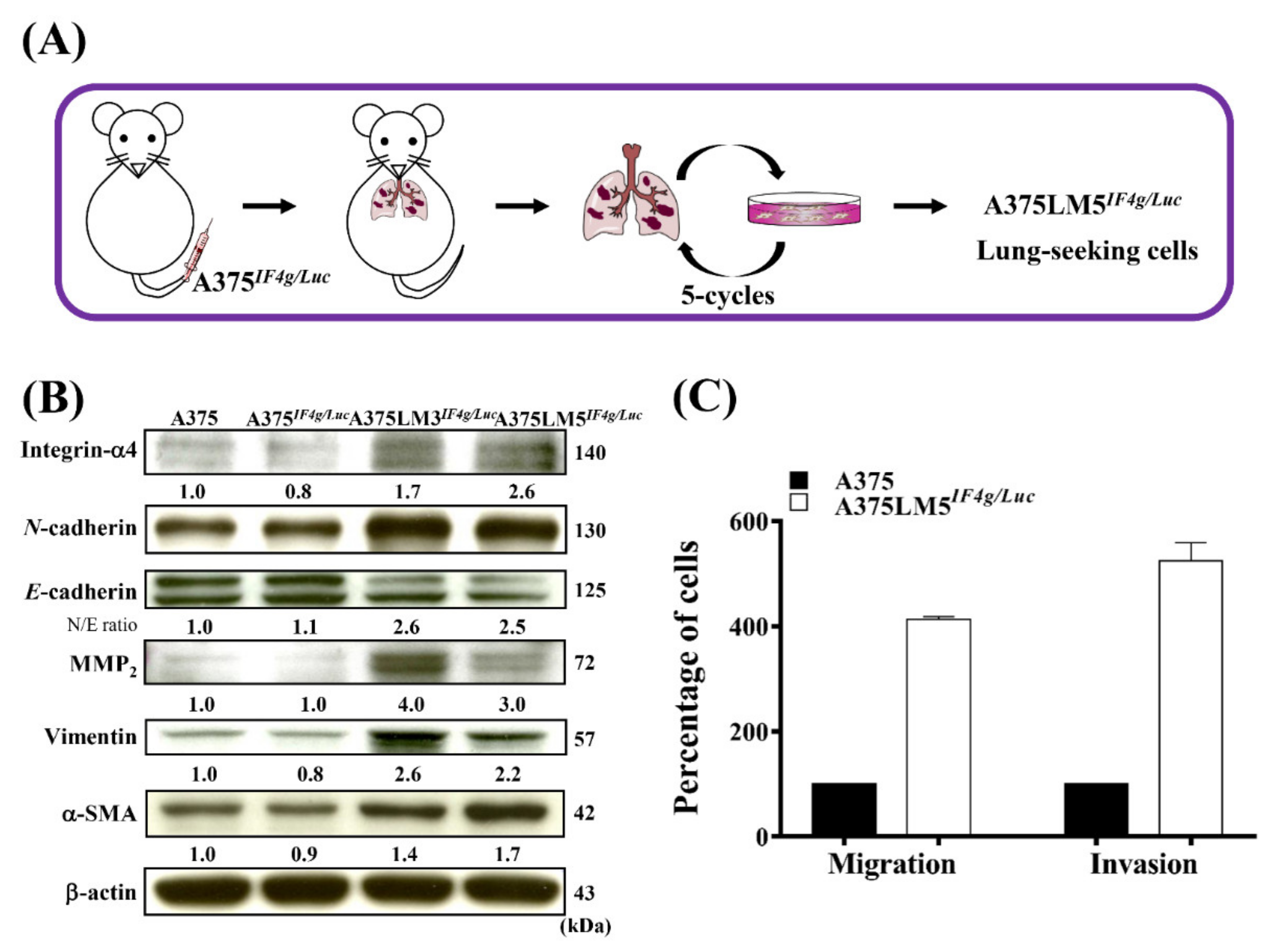
2.1. Establishment of A375LM5IF4g/Luc Lung-Seeking Melanoma Cells
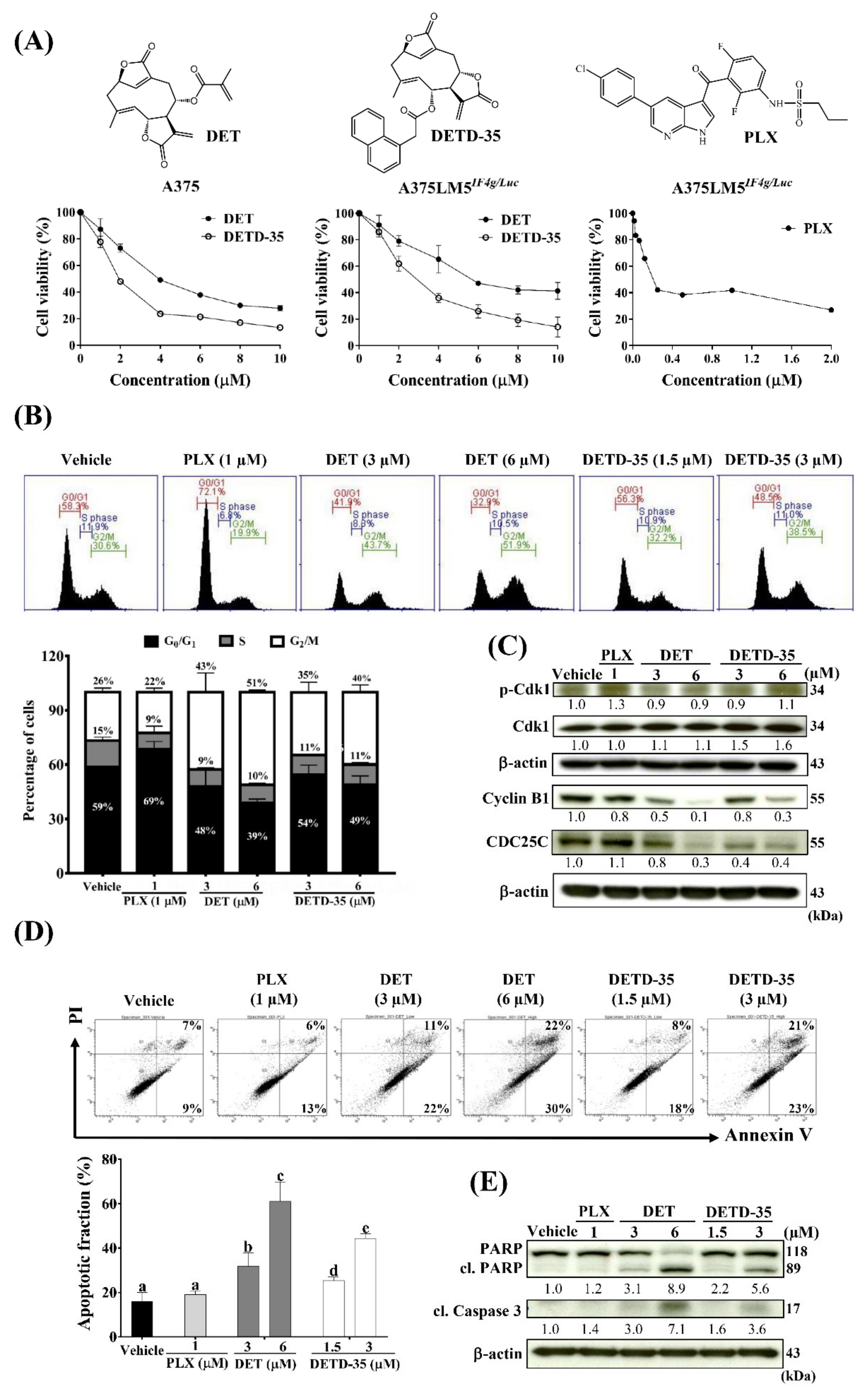
2.2. DET and DETD-35 Inhibited Proliferation and Induced G2/M-Phase Arrest in A375LM5IF4g/Luc Melanoma Cells
2.3. DET and DETD-35 Induced Apoptosis in A375LM5IF4g/Luc Melanoma Cells
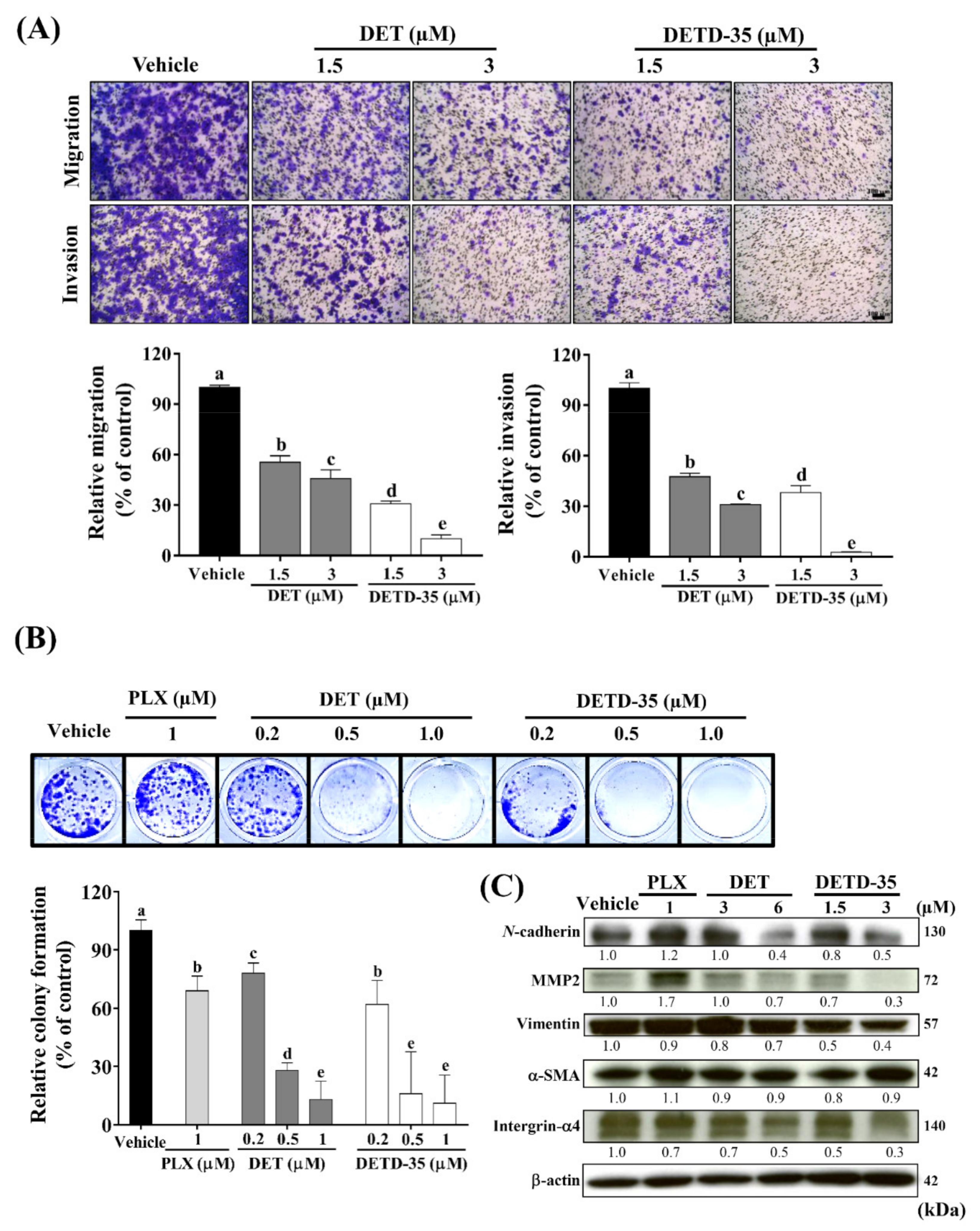
2.4. DET and DETD-35 Suppressed the Metastatic Properties of A375LM5IF4g/Luc Cells
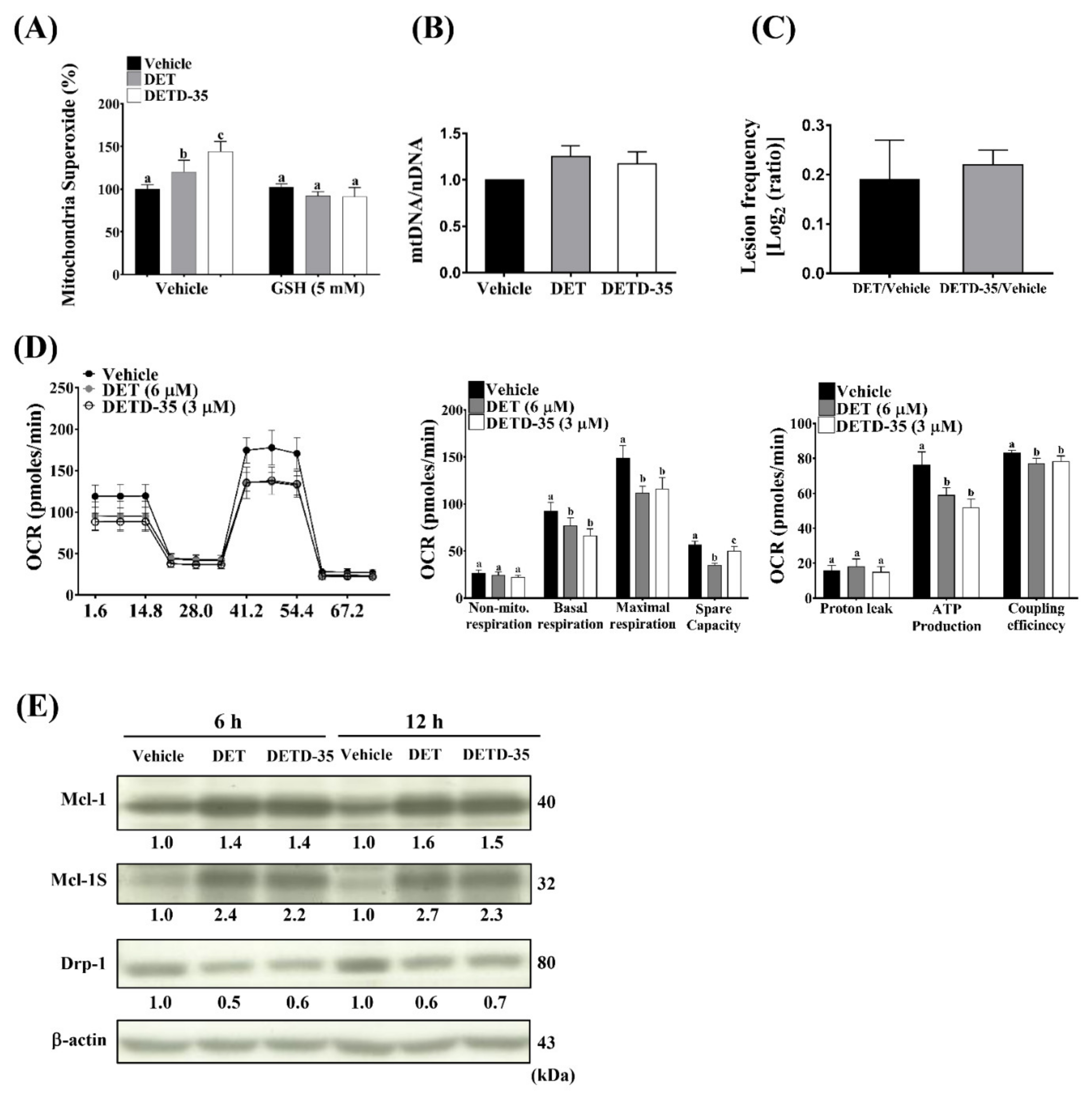
2.5. ROS Scavenger GSH Reversed DET- and DETD-35-Induced Oxidative Stress-Mediated Anti-Proliferative, Apoptotic and Migratory/Invasive Effects in A375LM5IF4g/Luc Cells
2.6. DET and DETD-35 Induced mtDNA Damage and Impaired Mitochondrial Function in A375LM5IF4g/Luc Cells
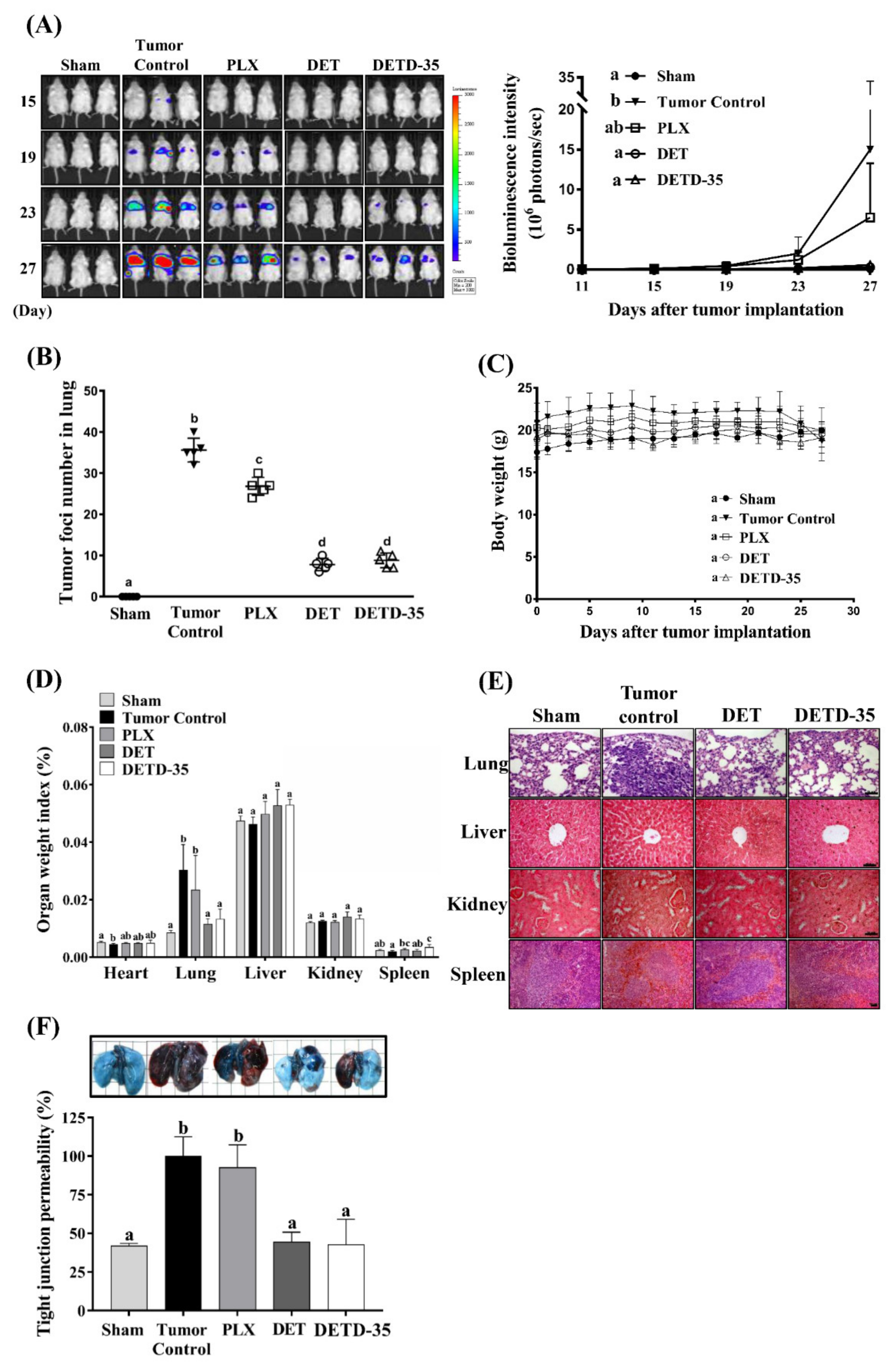
2.7. DET and DETD-35 Inhibited A375LM5IF4g/Luc Lung Metastasis in Xenograft Mice
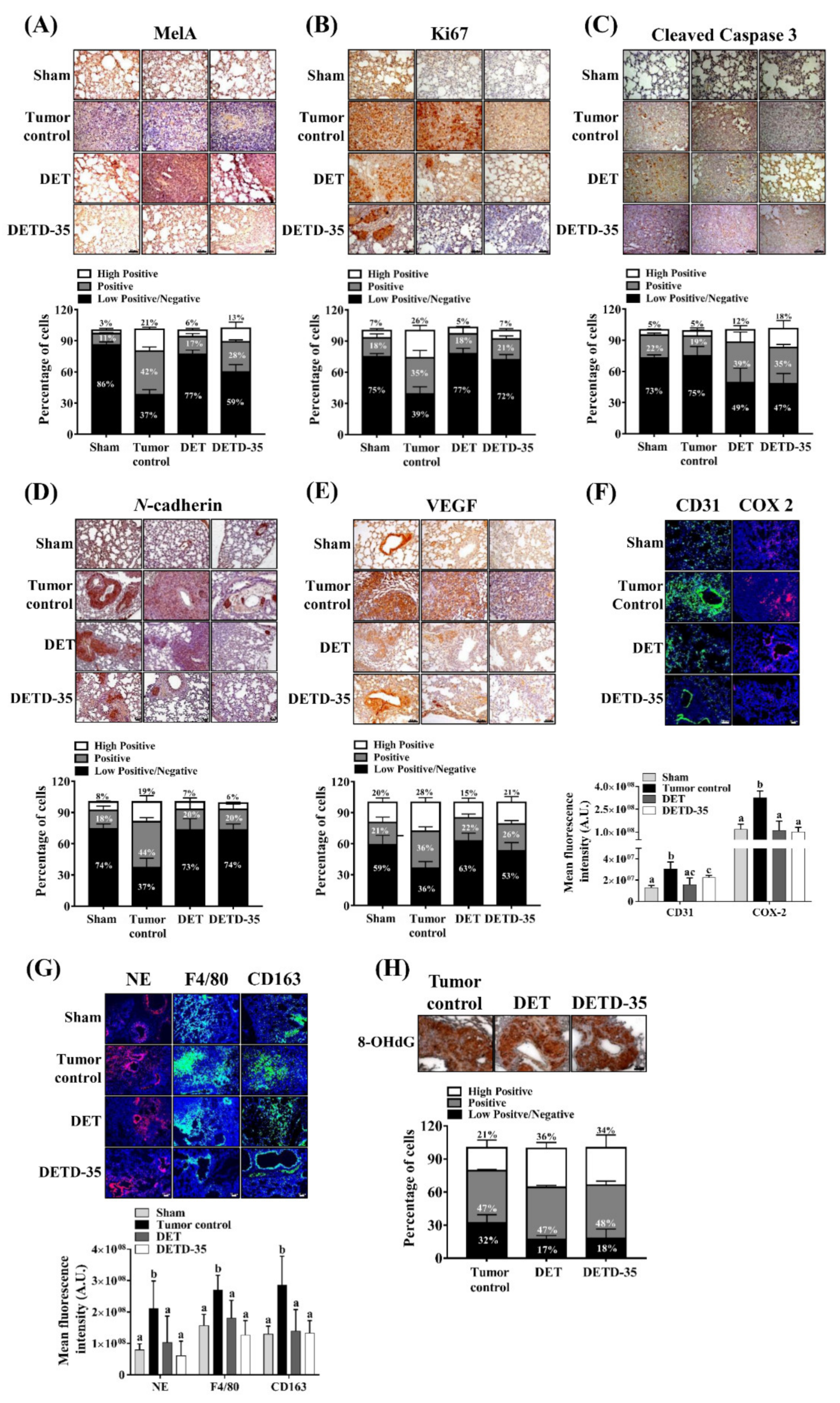
2.8. DET and DETD-35 Inhibited Various Biomarkers in the Lung Microenvironment of A375LM5IF4g/Luc Xenograft Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Compounds DET and DETD-35
4.3. Cell lines and Culture Conditions
4.4. Cell Viability Assay
4.5. Colony Formation Assay
4.6. Cell Cycle Analysis
4.7. Apoptosis Assay
4.8. Western Blot Analysis
4.9. Cell Migration/Invasion Assay
4.10. ROS Measurement
4.11. GSH Measurement
4.12. Mitochondrial DNA Damage Assay
4.13. Cell Mitochondria Stress Test
4.14. Animals
4.15. Experimental Lung Metastasis Mouse Model
4.16. Tight Junction Permeability Analysis
4.17. Immunofluorescence and Immunohistochemistry of Lung Tissues from A375LM5IF4g/Luc Metastatic Mouse Model
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Shain, A.H.; Bastian, B.C. From melanocytes to melanomas. Nat. Rev. Cancer 2016, 16, 345–358. [Google Scholar] [CrossRef]
- Tripp, M.K.; Watson, M.; Balk, S.J.; Swetter, S.M.; Gershenwald, J.E. State of the science on prevention and screening to reduce melanoma incidence and mortality: The time is now. CA A Cancer J. Clin. 2016, 66, 460–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, H.W.; Weinstock, M.A.; Feldman, S.R.; Coldiron, B.M. Incidence estimate of nonmelanoma skin cancer (Keratinocyte carcinomas) in the US population, 2012. JAMA Dermatol. 2015, 151, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Conic, R.Z.; Cabrera, C.I.; Khorana, A.A.; Gastman, B.R. Determination of the impact of melanoma surgical timing on survival using the National Cancer Database. J. Am. Acad. Dermatol. 2018, 78, 40–46.e7. [Google Scholar] [CrossRef] [PubMed]
- Ascierto, P.A.; Kirkwood, J.M.; Grob, J.-J.; Simeone, E.; Grimaldi, A.M.; Maio, M.; Palmieri, G.; Testori, A.; Marincola, F.M.; Mozzillo, N. The role of BRAF V600 mutation in melanoma. J. Transl. Med. 2012, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Cohen, M.S. The discovery of vemurafenib for the treatment of BRAF-mutated metastatic melanoma. Expert Opin. Drug Discov. 2016, 11, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Queirolo, P.; Picasso, V.; Spagnolo, F. Combined BRAF and MEK inhibition for the treatment of BRAF-mutated metastatic melanoma. Cancer Treat. Rev. 2015, 41, 519–526. [Google Scholar] [CrossRef]
- Maio, M. Melanoma as a model tumour for immuno-oncology. Ann. Oncol. 2012, 23, viii10–viii14. [Google Scholar] [CrossRef]
- Cochran, A.J.; Huang, R.-R.; Lee, J.; Itakura, E.; Leong, S.P.L.; Essner, R. Tumour–induced immune modulation of sentinel lymph nodes. Nat. Rev. Immunol. 2006, 6, 659–670. [Google Scholar] [CrossRef]
- Kim, J.; Bae, J.-S. Tumor-associated macrophages and neutrophils in tumor microenvironment. Mediat. Inflamm. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, J.A.; Fisher, D.E. The melanoma revolution: From UV carcinogenesis to a new era in therapeutics. Science 2014, 346, 945–949. [Google Scholar] [CrossRef] [Green Version]
- Gide, T.N.; Wilmott, J.S.; Scolyer, R.A.; Long, G.V. Primary and acquired resistance to immune checkpoint inhibitors in metastatic melanoma. Clin. Cancer Res. 2018, 24, 1260–1270. [Google Scholar] [CrossRef] [Green Version]
- Ashton, T.M.; McKenna, W.G.; Kunz-Schughart, L.A.; Higgins, G.S. Oxidative phosphorylation as an emerging target in cancer therapy. Clin. Cancer Res. 2018, 24, 2482–2490. [Google Scholar] [CrossRef] [Green Version]
- Fischer, G.M.; Jalali, A.; Yang, C.; Srivastava, A.; Ambati, C.R.; Sreekumar, A.; Hudgens, C.W.; Knighton, B.; Deng, W.; Ferguson, S.D.; et al. Molecular profiliang reveals unique immune and metabolic features of melanoma brain metastases. Cancer Discov. 2019, 9, 628–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, W.; Kumar, N.; Roginskaya, V.; Fouquerel, E.; Opresko, P.L.; Shiva, S.; Watkins, S.C.; Kolodieznyi, D.; Bruchez, M.P.; Van Houten, B. Chemoptogenetic damage to mitochondria causes rapid telomere dysfunction. Proc. Natl. Acad. Sci. USA 2019, 116, 18435–18444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.-T.; Yang, C.-C.; Shyur, L.-F. Phytomedicine—Modulating oxidative stress and the tumor microenvironment for cancer therapy. Pharmacol. Res. 2016, 114, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Piskounova, E.; Agathocleous, M.; Murphy, M.M.; Hu, Z.; Huddlestun, S.E.; Zhao, Z.; Leitch, A.M.; Johnson, T.M.; DeBerardinis, R.J.; Morrison, S.J. Oxidative stress inhibits distant metastasis by human melanoma cells. Nat. Cell Biol. 2015, 527, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Schumacker, P.T. Reactive oxygen species in cancer cells: Live by the sword, die by the sword. Cancer Cell 2006, 10, 175–176. [Google Scholar] [CrossRef] [Green Version]
- Gach, K.; Długosz, A.; Janecka, A. The role of oxidative stress in anticancer activity of sesquiterpene lactones. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.I. A List of Plants in Taiwan; Natural Publishing, Co.: Taipei, Taiwan, 1982; p. 1251. [Google Scholar]
- Huang, C.-C.; Lo, C.-P.; Chiu, C.-Y.; Shyur, L.-F. Deoxyelephantopin, a novel multifunctional agent, suppresses mammary tumour growth and lung metastasis and doubles survival time in mice. Br. J. Pharmacol. 2010, 159, 856–871. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-C.; Lin, K.-J.; Cheng, Y.-W.; Hsu, C.-A.; Yang, S.-S.; Shyur, L.-F. Hepatoprotective effect and mechanistic insights of deoxyelephantopin, a phyto-sesquiterpene lactone, against fulminant hepatitis. J. Nutr. Biochem. 2013, 24, 516–530. [Google Scholar] [CrossRef]
- Chao, W.-W.; Cheng, Y.-W.; Chen, Y.-R.; Lee, S.-H.; Chiou, C.-Y.; Shyur, L.-F. Phyto-sesquiterpene lactone deoxyelephantopin and cisplatin synergistically suppress lung metastasis of B16 melanoma in mice with reduced nephrotoxicity. Phytomedicine 2019, 56, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa-Goto, K.; Chen, J.-Y.; Cheng, Y.-T.; Lee, W.-L.; Takeya, M.; Saito, Y.; Lee, K.-H.; Shyur, L.-F. Novel sesquiterpene lactone analogues as potent anti-breast cancer agents. Mol. Oncol. 2016, 10, 921–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.-H.; Nakagawa-Goto, K.; Lee, K.-H.; Shyur, L.-F. A novel plant sesquiterpene lactone derivative, DETD-35, suppresses BRAFV600E mutant melanoma growth and overcomes acquired vemurafenib resistance in mice. Mol. Cancer Ther. 2016, 15, 1163–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodorogea, A.; Calinescu, A.; Popp, C.; Cioplea, M.; Nichita, L.; Hulea, I.; Brinzea, A.; Antohe, M.; Balaban, M.; Nedelcu, R.I.; et al. Epithelial-mesenchymal transition in skin cancers: A review. Anal. Cell. Pathol. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolillo, M.; Schinelli, S. Extracellular matrix alterations in metastatic processes. Int. J. Mol. Sci. 2019, 20, 4947. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Park, Y.M.; Kim, H.H.; Joo, K.M.; Lee, S.J.; Cho, H.J.; Kim, D.-H.; Lee, J.-I.; Kang, M.-S.; Seol, H.J.; et al. Alpha-smooth muscle actin (ACTA2) is required for metastatic potential of human lung adenocarcinoma. Clin. Cancer Res. 2013, 19, 5879–5889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, S.A.; McCabe, K.E.; Bartakova, A.; Delaney, J.; Pizzo, D.P.; Newbury, R.O.; Varner, J.A.; Schlaepfer, D.D.; Stupack, D.G. Integrin α4 enhances metastasis and may be associated with poor prognosis in mycnlow neuroblastoma. PLoS ONE 2015, 10, e0120815. [Google Scholar] [CrossRef] [Green Version]
- Paulitschke, V.; Berger, W.; Bileck, A.; Pirker, C.; Pehamberger, H.; Gerner, C.; Kunstfeld, R.; Paulitschke, P.; Hofstätter, E.; Knapp, B.; et al. Vemurafenib resistance signature by proteome analysis offers new strategies and rational therapeutic concepts. Mol. Cancer Ther. 2015, 14, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Dratkiewicz, E.; Simiczyjew, A.; Pietraszek-Gremplewicz, K.; Mazurkiewicz, J.; Nowak, D. Characterization of melanoma cell lines resistant to vemurafenib and evaluation of their responsiveness to EGFR-and met-inhibitor treatment. Int. J. Mol. Sci. 2019, 21, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribas, V.; Garcãa-Ruiz, C.; Fernã¡ndez-Checa, J.C. Glutathione and mitochondria. Front. Pharmacol. 2014, 5, 151. [Google Scholar] [CrossRef] [Green Version]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Yakes, F.M.; Van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc. Natl. Acad. Sci. USA 1997, 94, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Hunt, C.P.; Rooney, J.P.; Ryde, I.T.; Anbalagan, C.; Joglekar, R.; Meyer, J.N. PCR-based analysis of mitochondrial DNA copy number, mitochondrial DNA damage, and nuclear DNA damage. Curr. Protoc. Toxicol. 2016, 67, 20.11.1–20.11.25. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, V.K.; Lebrecht, D.; Nicholson, A.G.; Wells, A.; Bhayani, H.; Gazdhar, A.; Tamm, M.; Venhoff, N.; Geiser, T.; Walker, U.A. Mitochondrial DNA mutations and respiratory chain dysfunction in idiopathic and connective tissue disease-related lung fibrosis. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Gibson, S.B. MCL-1 is a gate keeper regulating cell death in cancer cells. J. Clin. Exp. Oncol. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Denisenko, T.V.; Gorbunova, A.S.; Zhivotovsky, B. Mitochondrial involvement in migration, invasion and metastasis. Front. Cell Dev. Biol. 2019, 7, 355. [Google Scholar] [CrossRef]
- Jiang, M.; Qin, C.; Han, M. Primary breast cancer induces pulmonary vascular hyperpermeability and promotes metastasis via the VEGF-PKC pathway. Mol. Carcinog. 2015, 55, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Xu, J.; Bi, Y.; Jiang, M.; Xu, X.; Liu, Q.; Jia, J. Primary tumor regulates the pulmonary microenvironment in melanoma carcinoma model and facilitates lung metastasis. Z. für Krebsforsch. 2012, 139, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free. Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef]
- Jafarian, A.H.; Roshan, N.M.; Gharib, M.; Moshirahmadi, V.; Tasbandi, A.; Ayatollahi, A.A.; Ayatollahi, H. Evaluation of cyclooxygenase-2 expression in association with clinical-pathological factors in malignant melanoma. Iran. J. Pathol. 2019, 14, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.R.; Siegelin, M.D.; Rompel, R.; Enk, A.H.; Gaiser, T. COX-2 expression in malignant melanoma: A novel prognostic marker? Melanoma Res. 2009, 19, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Im, A.; Kim, S.H.; Hyun, J.W.; Chae, S. Timosaponin AIII inhibits melanoma cell migration by suppressing COX -2 and in vivo tumor metastasis. Cancer Sci. 2016, 107, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Qin, J.; Li, Y.; Li, G.; Wang, Y.; Zhang, N.; Chen, P.; Li, C. Combination therapy of PKCζ and COX-2 inhibitors synergistically suppress melanoma metastasis. J. Exp. Clin. Cancer Res. 2017, 36, 115. [Google Scholar] [CrossRef] [Green Version]
- Sadhu, S.S.; Wang, S.; Averineni, R.K.; Seefeldt, T.; Yang, Y.; Guan, X. In-vitro and in-vivo inhibition of melanoma growth and metastasis by the drug combination of celecoxib and dacarbazine. Melanoma Res. 2016, 26, 572–579. [Google Scholar] [CrossRef]
- Fu, L.-Q.; Du, W.-L.; Cai, M.-H.; Yao, J.-Y.; Zhao, Y.-Y.; Mou, X.-Z. The roles of tumor-associated macrophages in tumor angiogenesis and metastasis. Cell. Immunol. 2020, 353, 104119. [Google Scholar] [CrossRef] [PubMed]
- Ghantous, A.; Gali-Muhtasib, H.; Vuorela, H.; Saliba, N.A.; Darwiche, N. What made sesquiterpene lactones reach cancer clinical trials? Drug Discov. Today 2010, 15, 668–678. [Google Scholar] [CrossRef]
- Kreuger, M.R.O.; Grootjans, S.; Biavatti, M.W.; Vandenabeele, P.; D’Herde, K. Sesquiterpene lactones as drugs with multiple targets in cancer treatment. Anti-Cancer Drugs 2012, 23, 883–896. [Google Scholar] [CrossRef]
- Cvetanova, B.; Shen, Y.-C.; Shyur, L.-F. Cumingianoside A, a phyto-triterpenoid saponin inhibits acquired braf inhibitor resistant melanoma growth via programmed cell death. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef]
- Yang, C.-C.; Chang, C.-K.; Chang, M.-T.; Shyur, L.-F. Plant galactolipid dLGG suppresses lung metastasis of melanoma through deregulating TNF-α-mediated pulmonary vascular permeability and circulating oxylipin dynamics in mice. Int. J. Cancer 2018, 143, 3248–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC profiler: An open source plugin for the quantitative evaluation and automated scoring of immunohistochemistry images of human tissue samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cvetanova, B.; Li, M.-Y.; Yang, C.-C.; Hsiao, P.-W.; Yang, Y.-C.; Feng, J.-H.; Shen, Y.-C.; Nakagawa-Goto, K.; Lee, K.-H.; Shyur, L.-F. Sesquiterpene Lactone Deoxyelephantopin Isolated from Elephantopus scaber and Its Derivative DETD-35 Suppress BRAFV600E Mutant Melanoma Lung Metastasis in Mice. Int. J. Mol. Sci. 2021, 22, 3226. https://doi.org/10.3390/ijms22063226
Cvetanova B, Li M-Y, Yang C-C, Hsiao P-W, Yang Y-C, Feng J-H, Shen Y-C, Nakagawa-Goto K, Lee K-H, Shyur L-F. Sesquiterpene Lactone Deoxyelephantopin Isolated from Elephantopus scaber and Its Derivative DETD-35 Suppress BRAFV600E Mutant Melanoma Lung Metastasis in Mice. International Journal of Molecular Sciences. 2021; 22(6):3226. https://doi.org/10.3390/ijms22063226
Chicago/Turabian StyleCvetanova, Biljana, Meng-Yi Li, Chung-Chih Yang, Pei-Wen Hsiao, Yu-Chih Yang, Jia-Hua Feng, Ya-Ching Shen, Kyoko Nakagawa-Goto, Kuo-Hsiung Lee, and Lie-Fen Shyur. 2021. "Sesquiterpene Lactone Deoxyelephantopin Isolated from Elephantopus scaber and Its Derivative DETD-35 Suppress BRAFV600E Mutant Melanoma Lung Metastasis in Mice" International Journal of Molecular Sciences 22, no. 6: 3226. https://doi.org/10.3390/ijms22063226