
Recent Advances Clarifying the Structure and Function of Plant Apyrases (Nucleoside Triphosphate Diphosphohydrolases)




Abstract
:
1. Introduction
2. Biochemical Properties of APY
3. Structural Organization of Plant Apyrases
4. Functional Studies on Plant APYs
4.1. Studies Linking APY Expression to Growth Control
Correlating eATP Treatments and APY Effects on Growth
4.2. APY Control of Stress Responses
5. Potential Value of Apyrase Inhibitors
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Zimmermann, H. Ectonucleoside triphosphate diphosphohydrolases and ecto-5′-nucleotidase in purinergic signaling: How the field developed and where we are now. Purinergic Signal. 2020, 1–9. [Google Scholar] [CrossRef]
- Herrero, A.B.; Uccelletti, D.; Hirschberg, C.B.; Dominguez, A.; Abeijon, C. The Golgi GDPase of the fungal pathogen Candida albicans affects morphogenesis, glycosylation, and cell wall properties. Eukaryot. Cell 2002, 1, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G.; Verkhratsky, A. Evolutionary origins of the purinergic signalling system. Acta Physiol. 2009, 195, 415–447. [Google Scholar] [CrossRef]
- Ho, J.; Theg, S.M. The Formation of Stromules In Vitro from Chloroplasts Isolated from Nicotiana benthamiana. PLoS ONE 2016, 11, e0146489. [Google Scholar] [CrossRef] [Green Version]
- Clark, G.; Roux, S.J. Role of Ca2+ in Mediating Plant Responses to Extracellular ATP and ADP. Int. J. Mol. Sci. 2018, 19, 3590. [Google Scholar] [CrossRef] [Green Version]
- Clark, G.B.; Morgan, R.O.; Fernandez, M.P.; Salmi, M.L.; Roux, S.J. Breakthroughs spotlighting roles for extracellular nucleotides and apyrases in stress responses and growth and development. Plant Sci. 2014, 225, 107–116. [Google Scholar] [CrossRef]
- Cannon, A.E.; Salmi, M.L.; Cantero, A.; Roux, S.J. Generation of Transgenic Spores of the Fern Ceratopteris Richardii to Analyze Ca2+ Transport Dynamics during Gravity-Directed Polarization; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Krishnan, P.S. The Preparation of Apyrase from Potato. Arch. Biochem. 1948, 16, 474–476. [Google Scholar] [PubMed]
- Wujak, M.; Banach, M.; Porowinska, D.; Piskulak, K.; Komoszynski, M. Isolation and bioinformatic analysis of seven genes encoding potato apyrase. Bacterial overexpresssion, refolding and initial kinetic studies on some recombinant potato apyrases. Phytochemistry 2013, 93, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Farani, P.S.G.; Marconato, D.G.; Emidio, N.B.; Pereira, V.R.D.; Alves, I.J.; da Silveira, L.S.; Couri, M.R.C.; Vasconcelos, E.G.; Castro-Borges, W.; Silva, A.A.; et al. Screening of plant derived chalcones on the inhibition of potato apyrase: Potential protein biotechnological applications in health. Int. J. Biol. Macromol. 2020, 164, 687–693. [Google Scholar] [CrossRef]
- Lomakina, G.Y.; Konik, P.A.; Ugarova, N.N. The Kinetics of Hydrolysis of ATP by Apyrase A from Solanum tuberosum. Mosc. Univ. Chem. Bull. 2020, 75, 374–381. [Google Scholar]
- Meng, R.; Zhu, L.Q.; Yang, Y.F.; Zhu, L.C.; Hou, Z.K.; Jin, L.; Wang, B.C. Apyrases in Arabidopsis thaliana. Biol. Plant. 2019, 63, 38–42. [Google Scholar] [CrossRef]
- Shibata, K.; Abe, S.; Yoneda, M.; Davies, E. Sub-cellular distribution and isotypes of a 49-kDa apyrase from Pisum sativum. Plant Physiol. Biochem. 2002, 40, 407–415. [Google Scholar] [CrossRef]
- Chen, Y.R.; Datta, N.; Roux, S.J. Purification and Partial Characterization of A Calmodulin-Stimulated Nucleoside Triphosphatase from Pea Nuclei. J. Biol. Chem. 1987, 262, 10689–10694. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Kim, S.Y.; Libault, M.; Berg, R.H.; Tanaka, K.; Stacey, G.; Taylor, C.G. GS52 Ecto-Apyrase Plays a Critical Role during Soybean Nodulation. Plant Physiol. 2009, 149, 994–1004. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Nguyen, C.T.; Libault, M.; Cheng, J.; Stacey, G. Enzymatic Activity of the Soybean Ecto-Apyrase GS52 Is Essential for Stimulation of Nodulation. Plant Physiol. 2011, 155, 1988–1998. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.B.; Ni, J.; Shah, F.A.; Ye, K.Q.; Hu, H.; Wang, Q.J.; Wang, D.D.; Yao, Y.Y.; Huang, S.W.; Hou, J.Y.; et al. Genome-wide identification, characterization and expression pattern analysis of APYRASE family members in response to abiotic and biotic stresses in wheat. PeerJ 2019, 7, e7622. [Google Scholar] [CrossRef]
- Matsumoto, H.; Yamaya, T.; Tanigawa, M. ACTIVATION OF ATPASE ACTIVITY IN THE CHROMATIN FRACTION OF PEA NUCLEI BY CALCIUM AND CALMODULIN. Plant Cell Physiol. 1984, 25, 191–195. [Google Scholar]
- Steinebrunner, I.; Jeter, C.; Song, C.; Roux, S.J. Molecular and biochemical comparison of two different apyrases from Arabidopsis thaliana. Plant Physiol. Biochem. 2000, 38, 913–922. [Google Scholar] [CrossRef]
- Massalski, C.; Bloch, J.; Zebisch, M.; Steinebrunner, I. The Biochemical Properties of the Arabidopsis Ecto-Nucleoside Triphosphate Diphosphohydrolase AtAPY1 Contradict a Direct Role in Purinergic Signaling. PLoS ONE 2015, 10, e0115832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, T.Y.; Christiansen, K.; Moreno, I.; Lao, J.M.; Loque, D.; Orellana, A.; Heazlewood, J.L.; Clark, G.; Roux, S.J. AtAPY1 and AtAPY2 Function as Golgi-Localized Nucleoside Diphosphatases in Arabidopsis thaliana. Plant Cell Physiol. 2012, 53, 1913–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, M.; Massalski, C.; Kurth, T.; Steinebrunner, I. The Arabidopsis apyrase AtAPY1 is localized in the Golgi instead of the extracellular space. BMC Plant Biol. 2012, 12, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, T.Y.; Lao, J.; Manalansan, B.; Loque, D.; Roux, S.J.; Heazlewood, J.L. Biochemical characterization of Arabidopsis APYRASE family reveals their roles in regulating endomembrane NDP/NMP homoeostasis. Biochem. J. 2015, 472, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Steinebrunner, I.; Sun, Y.; Butterfield, T.; Torres, J.; Arnold, D.; Gonzalez, A.; Jacob, F.; Reichler, S.; Roux, S.J. Apyrases (nucleoside triphosphate-diphosphohydrolases) play a key role in growth control in arabidopsis. Plant Physiol. 2007, 144, 961–975. [Google Scholar] [CrossRef] [Green Version]
- Clark, G.; Fraley, D.; Steinebrunner, I.; Cervantes, A.; Onyirimba, J.; Liu, A.; Torres, J.; Tang, W.Q.; Kim, J.; Roux, S.J. Extracellular Nucleotides and Apyrases Regulate Stomatal Aperture in Arabidopsis. Plant Physiol. 2011, 156, 1740–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, M.H.; Wu, J.; Yao, J.C.; Gallardo, I.F.; Dugger, J.W.; Webb, L.J.; Huang, J.; Salmi, M.L.; Song, J.W.; Clark, G.; et al. Apyrase Suppression Raises Extracellular ATP Levels and Induces Gene Expression and Cell Wall Changes Characteristic of Stress Responses1 C W OPEN. Plant Physiol. 2014, 164, 2054–2067. [Google Scholar] [CrossRef] [Green Version]
- Carson, M.; Johnson, D.H.; McDonald, H.; Brouillette, C.; DeLucas, L.J. His-tag impact on structure. Acta Crystallogr. Sect. D Biol. Crystallogr. 2007, 63, 295–301. [Google Scholar] [CrossRef]
- Ferenc-Mrozek, A.; Bojarska, E.; Stepinski, J.; Darzynkiewicz, E.; Lukaszewicz, M. Effect of the His-Tag Location on Decapping Scavenger Enzymes and Their Hydrolytic Activity toward Cap Analogs. ACS Omega 2020, 5, 10759–10766. [Google Scholar] [CrossRef]
- Abe, S.; Moustafa, M.F.M.; Shibata, K.; Yoneda, M.; Davies, E. Purification and characterization of the major isotypes of apyrase from the cytoskeleton fraction in Pisum sativum. Plant Physiol. Biochem. 2002, 40, 1019–1023. [Google Scholar] [CrossRef]
- Weeraratne, G.; Clark, G.; Roux, S.J. Biochemical Characterization of Nuclear Apyrases in Arabidopsis thaliana. In Proceedings of the Plant Biology Meeting, San Jose, CA, USA, 3–7 August 2019. [Google Scholar]
- Weeraratne, G. Genetic and Biochemical Studies of the Function of Apyrase 1 and Apyrase 2 in Etiolated Seedlings of Arabidopsis thaliana. Ph.D. Thesis, University of Texas at Austin, Austin, TX, USA, 2019. [Google Scholar]
- Li, H.; Hu, J.B.; Pang, J.; Zhao, L.T.; Yang, B.; Kang, X.L.; Wang, A.M.; Xu, T.D.; Yang, Z.B. Rho GTPase ROP1 Interactome Analysis Reveals Novel ROP1-Associated Pathways for Pollen Tube Polar Growth in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 7033. [Google Scholar] [CrossRef]
- Jiang, H.-W.; Clark, G.; Roux, S.J. PATL4 is an interacting partner of AtAPY1 and may be required for its growth-promoting functions. In Proceedings of the Plant Biology Meeting, San Jose, CA, USA, 3–7 August 2019. [Google Scholar]
- Tejos, R.; Rodriguez-Furlan, C.; Adamowski, M.; Sauer, M.; Norambuena, L.; Friml, J. PATELLINS are regulators of auxin-mediated PIN1 relocation and plant development in Arabidopsis thaliana. J. Cell Sci. 2018, 131, jcs204198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.R.; Sun, J.; Zhao, R.; Ding, M.Q.; Zhang, Y.N.; Sun, Y.L.; Wang, W.; Tan, Y.Q.; Liu, D.D.; Ma, X.J.; et al. Populus euphratica APYRASE2 Enhances Cold Tolerance by Modulating Vesicular Trafficking and Extracellular ATP in Arabidopsis Plants. Plant Physiol. 2015, 169, 530–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windsor, B.; Roux, S.J.; Lloyd, A. Multiherbicide tolerance conferred by AtPgp1 and apyrase overexpression in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Song, C.J.; Steinebrunner, I.; Wang, X.Z.; Stout, S.C.; Roux, S.J. Extracellular ATP induces the accumulation of superoxide via NADPH oxidases in Arabidopsis. Plant Physiol. 2006, 140, 1222–1232. [Google Scholar] [CrossRef] [Green Version]
- Kukulski, F.; Komoszynski, M. Purification and characterization of NTPDase1 (ecto-apyrase) and NTPDase2 (ecto-ATPase) from porcine brain cortex synaptosomes. Eur. J. Biochem. 2003, 270, 3447–3454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yegutkin, G.G. Enzymes involved in metabolism of extracellular nucleotides and nucleosides: Functional implications and measurement of activities. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 473–497. [Google Scholar] [CrossRef]
- Kozakiewicz, A.; Neurnann, P.; Banach, M.; Komoszyiski, M.; Wojtczak, A. Modeling studies of potato nucleoside triphosphate diphosphohydrolase NTPDase1: An insight into the catalytic mechanism. Acta Biochim. Pol. 2008, 55, 141–150. [Google Scholar] [CrossRef]
- Summers, E.L.; Cumming, M.H.; Oulavallickal, T.; Roberts, N.J.; Arcus, V.L. Structures and kinetics for plant nucleoside triphosphate diphosphohydrolases support a domain motion catalytic mechanism. Protein Sci. 2017, 26, 1627–1638. [Google Scholar] [CrossRef] [Green Version]
- Zebisch, M.; Krauss, M.; Schafer, P.; Strater, N. Structures of Legionella pneumophila NTPDase1 in complex with polyoxometallates. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 1147–1154. [Google Scholar] [CrossRef]
- Zebisch, M.; Krauss, M.; Schafer, P.; Strater, N. Crystallographic Evidence for a Domain Motion in Rat Nucleoside Triphosphate Diphosphohydrolase (NTPDase) 1. J. Mol. Biol. 2012, 415, 288–306. [Google Scholar] [CrossRef]
- Zebisch, M.; Baqi, Y.; Schafer, P.; Muller, C.E.; Strater, N. Crystal structure of NTPDase2 in complex with the sulfoanthraquinone inhibitor PSB-071. J. Struct. Biol. 2014, 185, 336–341. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER: Fully automated protein structure prediction in CASP8. Proteins Struct. Funct. Bioinform. 2009, 77, 100–113. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Yang, J.Y.; Zhang, Y. COFACTOR: An accurate comparative algorithm for structure-based protein function annotation. Nucleic Acids Res. 2012, 40, W471–W477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.Y.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.M.; Kirley, T.L. Site-directed mutagenesis of a human brain ecto-apyrase: Evidence that the E-type ATPases are related to the actin/heat shock 70/sugar kinase superfamily. Biochemistry 1999, 38, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H. The sugar kinase heat shock protein 70 actin super family: Implications of conserved structure for mechanism. Annu. Rev. Biophys. Biomol. Struct. 1996, 25, 137–162. [Google Scholar] [CrossRef]
- Zebisch, M.; Strater, N. Structural insight into signal conversion and inactivation by NTPDase2 in purinergic signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 6882–6887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zebisch, M.; Krauss, M.; Schafer, P.; Lauble, P.; Strater, N. Crystallographic Snapshots along the Reaction Pathway of Nucleoside Triphosphate Diphosphohydrolases. Structure 2013, 21, 1460–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, A.S.; Bradley, A.R.; Valasatava, Y.; Duarte, J.M.; Prlic, A.; Rose, P.W. NGL viewer: Web-based molecular graphics for large complexes. Bioinformatics 2018, 34, 755–3758. [Google Scholar] [CrossRef] [Green Version]
- Day, R.B.; McAlvin, C.B.; Loh, J.T.; Denny, R.L.; Wood, T.C.; Young, N.D.; Stacey, G. Differential expression of two soybean apyrases, one of which is an early nodulin. Mol. Plant-Microbe Interact. 2000, 13, 1053–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsirigos, K.D.; Peters, C.; Shu, N.; Kall, L.; Elofsson, A. The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides. Nucleic Acids Res. 2015, 43, W401–W407. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, S.J.; Steinebrunner, I. Extracellular ATP: An unexpected role as a signaler in plants. Trends Plant Sci. 2007, 12, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.W.; Liu, B.; Li, J.X.; Yu, N.N.; Zou, X.X.; Chen, L.P. Light- and temperature-regulated BjAPY2 may have a role in stem expansion of Brassica juncea. Funct. Integr. Genom. 2015, 15, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Reichler, S.A.; Torres, J.; Rivera, A.L.; Cintolesi, V.A.; Clark, G.; Roux, S.J. Intersection of two signalling pathways: Extracellular nucleotides regulate pollen germination and pollen tube growth via nitric oxide. J. Exp. Bot. 2009, 60, 2129–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, G.; Wu, M.; Wat, N.; Onyirimba, J.; Pham, T.; Herz, N.; Ogoti, J.; Gomez, D.; Canales, A.A.; Aranda, G.; et al. Both the stimulation and inhibition of root hair growth induced by extracellular nucleotides in Arabidopsis are mediated by nitric oxide and reactive oxygen species. Plant Mol. Biol. 2010, 74, 423–435. [Google Scholar] [CrossRef]
- Clark, G.; Torres, J.; Finlayson, S.; Guan, X.Y.; Handley, C.; Lee, J.; Kays, J.E.; Chen, Z.J.; Roux, S.J. Apyrase (Nucleoside Triphosphate-Diphosphohydrolase) and Extracellular Nucleotides Regulate Cotton Fiber Elongation in Cultured Ovules. Plant Physiol. 2010, 152, 1073–1083. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.S.; Qin, B.Z.; Feng, K.L.; Yan, R.L.; Kang, E.F.; Liu, T.; Shang, Z.L. Extracellular ATP promoted pollen germination and tube growth of Nicotiana tabacum through promoting K+ and Ca2+ absorption. Plant Reprod. 2018, 31, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.J.; Dong, X.X.; Xue, Y.Y.; Xu, J.W.; Zhang, A.Q.; Feng, M.; Zhao, Q.; Xia, S.Y.; Yin, Y.H.; He, S.H.; et al. Redox-Responsive Transcription Factor 1 (PRFT1) Is Involved in Extracellular ATP-Regulated Arabidopsis thaliana Seedling Growth. Plant Cell Physiol. 2020, 61, 685–698. [Google Scholar] [CrossRef]
- Roux, S.J.; Song, C.; Jeter, C. Regulation of plant growth and development by extracellular nucleotides. In Commun. Plants Neuronal Asp. Plant Life; Baluška, F., Mancuso, S., Volkmann, D., Eds.; Springer: Berlin/Heidelberg, Germany. [CrossRef]
- Liu, X.; Wu, J.; Clark, G.; Lundy, S.; Lim, M.; Arnold, D.; Chan, J.; Tang, W.Q.; Muday, G.K.; Gardner, G.; et al. Role for Apyrases in Polar Auxin Transport in Arabidopsis. Plant Physiol. 2012, 160, 1985–1995. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.X.; Zhu, R.J.; Kang, E.F.; Shang, Z.L. RRFT1 (Redox Responsive Transcription Factor 1) is involved in extracellular ATP-regulated gene expression in Arabidopsis thaliana seedlings. Plant Signal. Behav. 2020, 15, 1748282. [Google Scholar] [CrossRef]
- Vandenbussche, F.; Vaseva, I.; Vissenberg, K.; Van Der Straeten, D. Ethylene in vegetative development: A tale with a riddle. New Phytol. 2012, 194, 895–909. [Google Scholar] [CrossRef]
- Vaseva, I.I.; Qudeimat, E.; Potuschak, T.; Du, Y.L.; Genschik, P.; Vandenbussche, F.; Van Der Straeten, D. The plant hormone ethylene restricts Arabidopsis growth via the epidermis. Proc. Natl. Acad. Sci. USA 2018, 115, E4130–E4139. [Google Scholar] [CrossRef] [Green Version]
- Chivasa, S. Insights into Plant Extracellular ATP Signaling Revealed by the Discovery of an ATP-Regulated Transcription Factor. Plant Cell Physiol. 2020, 61, 673–674. [Google Scholar] [CrossRef]
- Pham, A.Q.; Cho, S.H.; Nguyen, C.T.; Stacey, G. Arabidopsis Lectin Receptor Kinase P2K2 Is a Second Plant Receptor for Extracellular ATP and Contributes to Innate Immunity(1)( OPEN ). Plant Physiol. 2020, 183, 1364–1375. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Wang, B.C.; Farris, B.; Clark, G.; Roux, S.J. Modulation of Root Skewing in Arabidopsis by Apyrases and Extracellular ATP. Plant Cell Physiol. 2015, 56, 2197–2206. [Google Scholar] [CrossRef] [Green Version]
- Clark, G.; Vanegas, D.; Cannon, A.; Hage, W.; Jankovic, M.; McLamore, E.; Roux, S. Extracellular ATP levels are higher in the growth zones of primary roots of wild-type Arabidopsis seedlings and are changed by altered expression of apyrase enzymes and auxin transporters. In Proceedings of the Plant Biology Meeting, Honolulu, HI, USA, 24–28 June 2017. [Google Scholar]
- Vanegas, D.C.; Clark, G.; Cannon, A.E.; Roux, S.; Chaturvedi, P.; McLamore, E.S. A self-referencing biosensor for real-time monitoring of physiological ATP transport in plant systems. Biosens. Bioelectron. 2015, 74, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Krogman, W.; Sparks, J.A.; Blancaflor, E.B. Cell Type-Specific Imaging of Calcium Signaling inArabidopsis thalianaSeedling Roots Using GCaMP3. Int. J. Mol. Sci. 2020, 21, 6385. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Stacey, G.; Leblanc-Fournier, N.; Legue, V.; Moulia, B.; Davies, J.M. Early Extracellular ATP Signaling in Arabidopsis Root Epidermis: A Multi-Conductance Process. Front. Plant Sci. 2019, 10, 1064. [Google Scholar] [CrossRef]
- Mohammad-Sidik, A.; Sun, J.; Shin, R.; Song, Z.Z.; Ning, Y.Z.; Matthus, E.; Wilkins, K.A.; Davies, J.M. Annexin 1 Is a Component of eATP-Induced Cytosolic Calcium Elevation in Arabidopsis thaliana Roots. Int. J. Mol. Sci. 2021, 22, 494. [Google Scholar] [CrossRef]
- Matthus, E.; Wilkins, K.A.; Swarbreck, S.M.; Doddrell, N.H.; Doccula, F.G.; Costa, A.; Davies, J.M. Phosphate Starvation Alters Abiotic-Stress-Induced Cytosolic Free Calcium Increases in Roots. Plant Physiol. 2019, 179, 1754–1767. [Google Scholar] [CrossRef] [Green Version]
- Matthus, E.; Wilkins, K.A.; Davies, J.M. Iron availability modulates the Arabidopsis thaliana root calcium signature evoked by exogenous ATP. Plant Signal. Behav. 2019, 14, 1640563. [Google Scholar] [CrossRef] [PubMed]
- Scheerer, U.; Trube, N.; Netzer, F.; Rennenberg, H.; Herschbach, C. ATP as Phosphorus and Nitrogen Source for Nutrient Uptake by Fagus sylvatica and Populus x canescens Roots. Front. Plant Sci. 2019, 10, 378. [Google Scholar] [CrossRef] [PubMed]
- Matthus, E.; Sun, J.; Wang, L.M.; Bhat, M.G.; Mohammad-Sidik, A.B.; Wilkins, K.A.; Leblanc-Fournier, N.; Legue, V.; Moulia, B.; Stacey, G.; et al. DORN1/P2K1 and purino-calcium signalling in plants: Making waves with extracellular ATP. Ann. Bot. 2019, 124, 1227–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veerappa, R.; Slocum, R.D.; Siegenthaler, A.; Wang, J.; Clark, G.; Roux, S.J. Ectopic expression of a pea apyrase enhances root system architecture and drought survival in Arabidopsis and soybean. Plant Cell Environ. 2019, 42, 337–353. [Google Scholar] [CrossRef]
- Sharma, T.; Morita, E.H.; Abe, S. Expression pattern of PsAPY1 during apical hook development in pea. Biologia 2014, 69, 293–299. [Google Scholar] [CrossRef]
- Lang, T.; Deng, C.; Yao, J.; Zhang, H.L.; Wang, Y.; Deng, S.R. A Salt-Signaling Network Involving Ethylene, Extracellular ATP, Hydrogen Peroxide, and Calcium Mediates K+/Na+ Homeostasis in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 8683. [Google Scholar] [CrossRef]
- Jewell, J.B.; Tanaka, K. Transcriptomic perspective on extracellular ATP signaling: A few curious trifles. Plant Signal. Behav. 2019, 14, 1659079. [Google Scholar] [CrossRef]
- Choi, J.; Tanaka, K.; Cao, Y.R.; Qi, Y.; Qiu, J.; Liang, Y.; Lee, S.Y.; Stacey, G. Identification of a Plant Receptor for Extracellular ATP. Science 2014, 343, 290–294. [Google Scholar] [CrossRef]
- Jewell, J.B.; Sowders, J.M.; He, R.F.; Willis, M.A.; Gang, D.R.; Tanaka, K. Extracellular ATP Shapes a Defense-Related Transcriptome Both Independently and along with Other Defense Signaling Pathways. Plant Physiol. 2019, 179, 1144–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Q.Z.; Wang, Y.P.; Fan, B.Q.; Sun, K.; Liang, J.Y.; Feng, H.Q.; Jia, L.Y. Extracellular ATP affects cell viability, respiratory O-2 uptake, and intracellular ATP production of tobacco cell suspension culture in response to hydrogen peroxide-induced oxidative stress. Biologia 2020, 75, 1437–1443. [Google Scholar] [CrossRef]
- Toyoda, K.; Yao, S.; Takagi, M.; Uchioki, M.; Miki, M.; Tanaka, K.; Suzuki, T.; Amano, M.; Kiba, A.; Kato, T.; et al. The plant cell wall as a site for molecular contacts in fungal pathogenesis. Physiol. Mol. Plant Pathol. 2016, 95, 44–49. [Google Scholar] [CrossRef]
- Toyoda, K.; Yasunaga, E.; Niwa, M.; Ohwatari, Y.; Nakashima, A.; Inagaki, Y.; Ichinose, Y.; Shiraishi, T. H2O2 production by copper amine oxidase, a component of the ecto-apyrase (ATPase)-containing protein complex(es) in the pea cell wall, is regulated by an elicitor and a suppressor from Mycosphaerella pinodes. J. Gen. Plant Pathol. 2012, 78, 311–315. [Google Scholar] [CrossRef]
- Wang, Q.W.; Jia, L.Y.; Shi, D.L.; Wang, R.F.; Lu, L.N.; Xie, J.J.; Sun, K.; Feng, H.Q.; Li, X. Effects of extracellular ATP on local and systemic responses of bean (Phaseolus vulgaris L) leaves to wounding. Biosci. Biotechnol. Biochem. 2019, 83, 417–428. [Google Scholar] [CrossRef]
- Tanaka, K.; Toth, K.; Stacey, G. Role of Ectoapyrases in Nodulation. Biol. Nitrogen Fixat. 2015, 517–524. [Google Scholar] [CrossRef]
- Guiguet, A.; Dubreuil, G.; Harris, M.O.; Appel, H.M.; Schultz, J.C.; Pereira, M.H.; Giron, D. Shared weapons of blood- and plant-feeding insects: Surprising commonalities for manipulating hosts. J. Insect Physiol. 2016, 84, 4–21. [Google Scholar] [CrossRef]
- Saraiva, N.B.; Emidio, N.B.; Vital, C.E.; Gusmao, M.A.N.; Marconato, D.G.; Coutinho, F.S.; Pereira, J.F.; Auad, A.M.; Faria-Pinto, P.; Ramos, H.J.O.; et al. Molecular profiling of the Mahanarva spectabilis salivary glands and phytohormonal response of elephant grass. Int. J. Trop. Insect Sci. 2021, 41, 849–860. [Google Scholar] [CrossRef]
- Meghji, P.; Burnstock, G. Inhibition of Extracellular ATP Degradation in Endothelial-Cells. Life Sci. 1995, 57, 763–771. [Google Scholar] [CrossRef]
- Long, B.; Li, G.H.; Feng, R.R.; Tao, L.; Liu, D.D.; Feng, H.Q.; Qin, Q.M. NTPDase Specific Inhibitors Suppress Rice Infection by Magnaporthe oryzae. J. Phytopathol. 2015, 163, 227–232. [Google Scholar] [CrossRef]
- Windsor, J.B.; Thomas, C.; Hurley, L.; Roux, S.J.; Lloyd, A.M. Automated colorimetric screen for apyrase inhibitors. Biotechniques 2002, 33, 1024–1030. [Google Scholar] [CrossRef]
- Tripathy, M.K.; Weeraratne, G.; Clark, G.; Roux, S.J. Apyrase inhibitors enhance the ability of diverse fungicides to inhibit the growth of different plant-pathogenic fungi. Mol. Plant Pathol. 2017, 18, 1012–1023. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowles, A.F. The GDA1_CD39 superfamily: NTPDases with diverse functions. Purinergic Signal. 2011, 7, 21–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Advances | References |
|---|---|
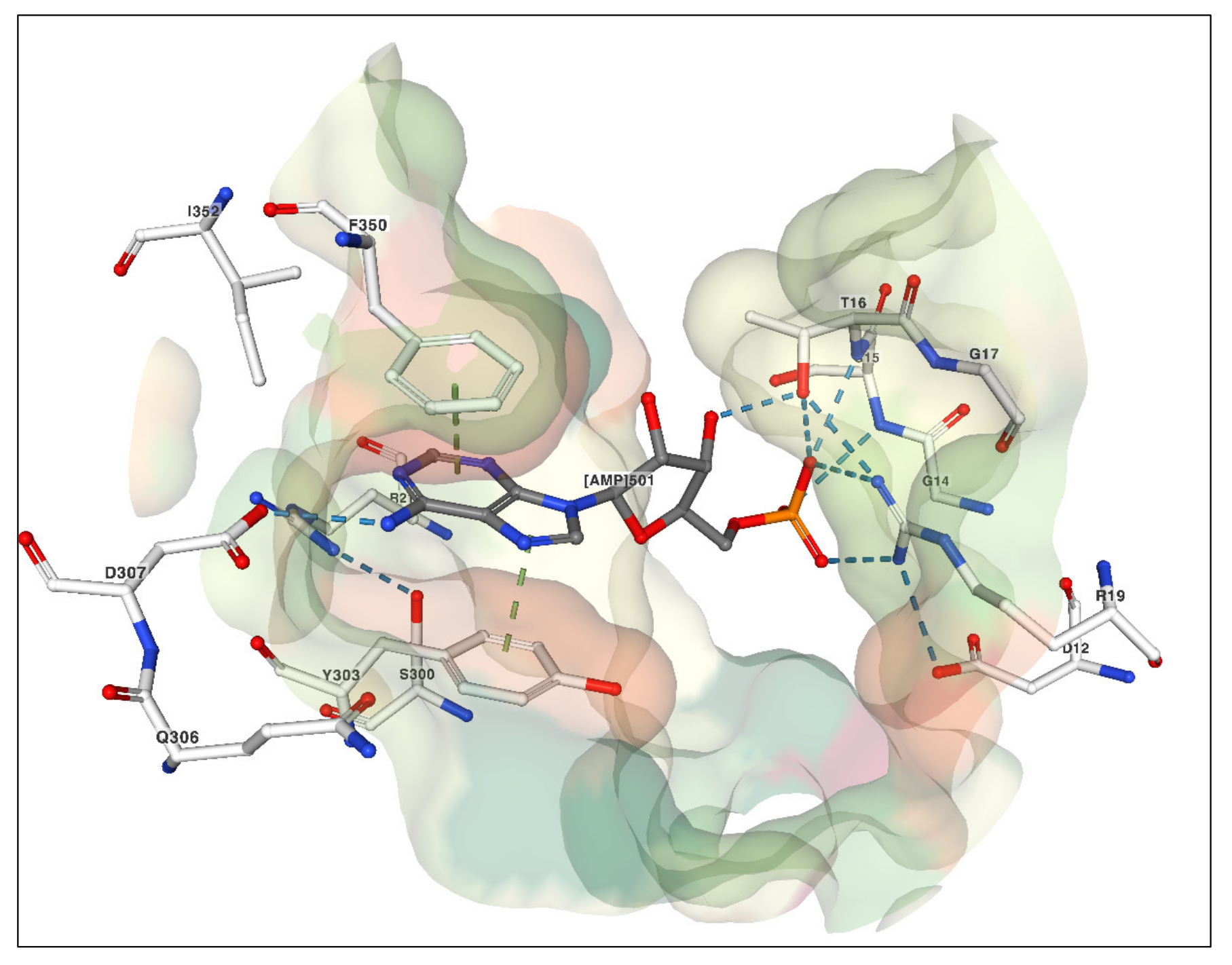
| New crystal structures, insights on NTP binding domain | [41] |
| New evidence that APYs can hydrolyze ATP in ECM and nuclei | [30,35,88,90] |
| Initial identification of potential binding partners for APYs | [32,33,88] |
| New evidence for APY protective role in abiotic stress responses | [17,35,80] |
| New evidence for APY protective role in defense against pathogens | [87,88,96] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clark, G.; Brown, K.A.; Tripathy, M.K.; Roux, S.J. Recent Advances Clarifying the Structure and Function of Plant Apyrases (Nucleoside Triphosphate Diphosphohydrolases). Int. J. Mol. Sci. 2021, 22, 3283. https://doi.org/10.3390/ijms22063283
Clark G, Brown KA, Tripathy MK, Roux SJ. Recent Advances Clarifying the Structure and Function of Plant Apyrases (Nucleoside Triphosphate Diphosphohydrolases). International Journal of Molecular Sciences. 2021; 22(6):3283. https://doi.org/10.3390/ijms22063283
Chicago/Turabian StyleClark, Greg, Katherine A. Brown, Manas K. Tripathy, and Stanley J. Roux. 2021. "Recent Advances Clarifying the Structure and Function of Plant Apyrases (Nucleoside Triphosphate Diphosphohydrolases)" International Journal of Molecular Sciences 22, no. 6: 3283. https://doi.org/10.3390/ijms22063283
APA StyleClark, G., Brown, K. A., Tripathy, M. K., & Roux, S. J. (2021). Recent Advances Clarifying the Structure and Function of Plant Apyrases (Nucleoside Triphosphate Diphosphohydrolases). International Journal of Molecular Sciences, 22(6), 3283. https://doi.org/10.3390/ijms22063283