
Poly(ADP-Ribose) Polymerase 1 Promotes Inflammation and Fibrosis in a Mouse Model of Chronic Pancreatitis

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. PARP Inhibition by Olaparib Reduces Tissue Injury in CP
2.2. Olaparib Suppresses Expression of Inflammatory Mediators in CP
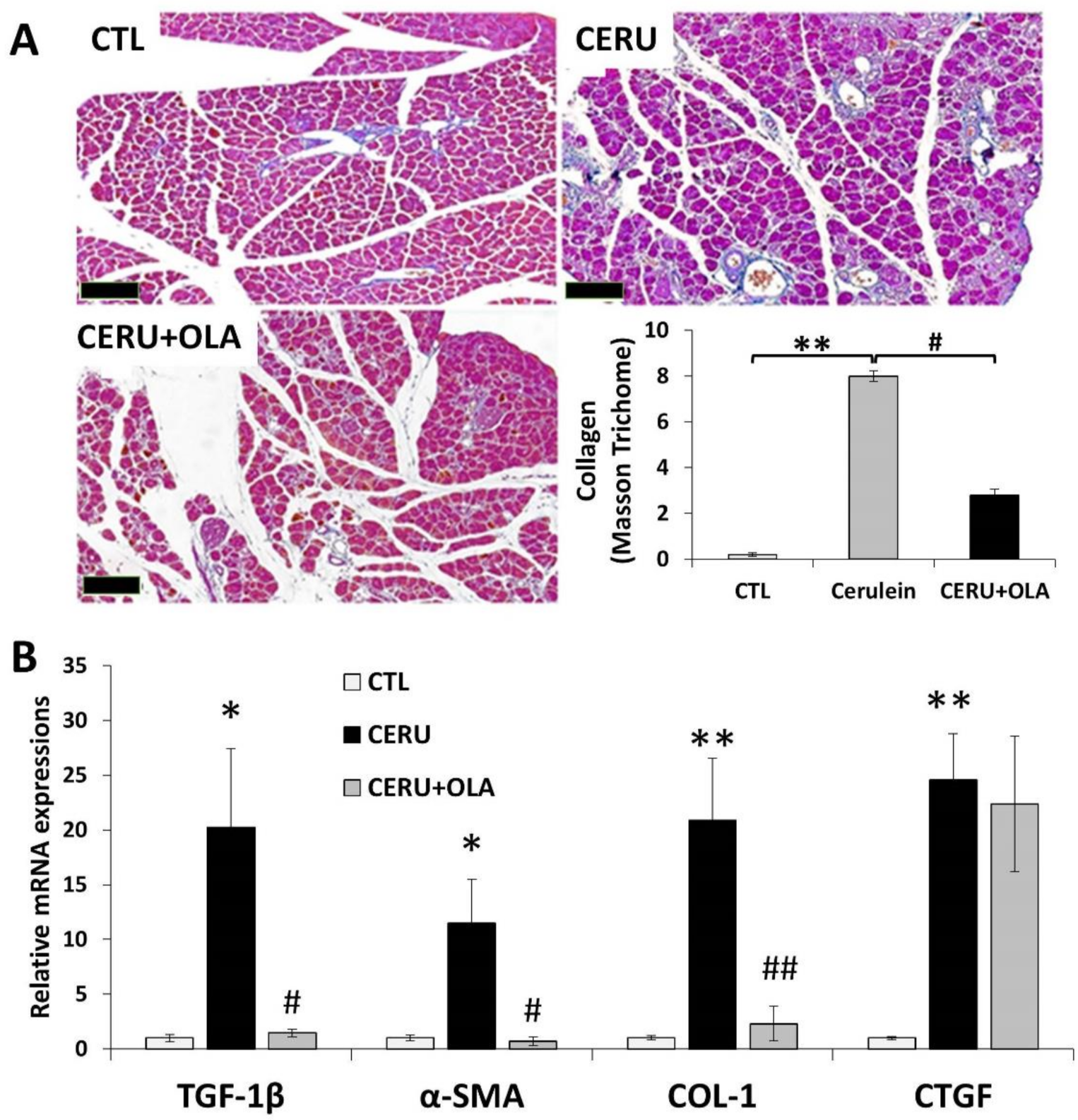
2.3. Olaparib Suppresses CP-Associated Pancreatic Fibrosis
2.4. Reduced Pancreatic Injury and Fibrosis in PARP1 Knockout Animals
2.5. Reduced Acinar Cell Damage/Death and Granulocyte Infiltration in PARP1 Knockout Animals
3. Discussion
4. Materials and Methods
4.1. Chronic Pancreatitis Model
4.2. Lactate Dehydrogenase (LDH), Amylase and Lipase Activity Measurements
4.3. H&E and Masson’s Trichrome
4.4. Measurement of Cell Viability (MTT Assay) and High-Content Analysis (HCA) Cytotoxicity
4.5. RNA Extraction and Real-Time Quantitative PCR Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP | acute pancreatitis |
| BSA | bovine serum albumin |
| CERU | Cerulein |
| CP | chronic pancreatitis |
| HCA | high-content analysis |
| LDH | lactate dehydrogenase |
| MPO | myeloperoxidase |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| OLA | olaparib |
| PAR | poly(ADP-ribose) |
| PARP | poly(ADP-ribose) polymerase |
| PARPi | PARP inhibitor |
| PARylation | poly(ADP-ribosyl)ation |
| PI | propidium iodide |
| SMA | smooth muscle actin |
References
- Czako, L.; Hegyi, P.; Rakonczay, Z., Jr.; Wittmann, T.; Otsuki, M. Interactions between the endocrine and exocrine pancreas and their clinical relevance. Pancreatology 2009, 9, 351–359. [Google Scholar] [CrossRef]
- Hegyi, P.J.; Soos, A.; Toth, E.; Ebert, A.; Venglovecz, V.; Marta, K.; Matrai, P.; Miko, A.; Bajor, J.; Sarlos, P.; et al. Evidence for diagnosis of early chronic pancreatitis after three episodes of acute pancreatitis: A cross-sectional multicentre international study with experimental animal model. Sci. Rep. 2021, 11, 1367. [Google Scholar] [CrossRef]
- Whitcomb, D.C.; Shimosegawa, T.; Chari, S.T.; Forsmark, C.E.; Frulloni, L.; Garg, P.; Hegyi, P.; Hirooka, Y.; Irisawa, A.; Ishikawa, T.; et al. International consensus statements on early chronic Pancreatitis. Recommendations from the working group for the international consensus guidelines for chronic pancreatitis in collaboration with The International Association of Pancreatology, American Pancreatic Association, Japan Pancreas Society, PancreasFest Working Group and European Pancreatic Club. Pancreatology 2018, 18, 516–527. [Google Scholar] [CrossRef]
- Hegyi, P.; Parniczky, A.; Lerch, M.M.; Sheel, A.R.G.; Rebours, V.; Forsmark, C.E.; Del Chiaro, M.; Rosendahl, J.; de-Madaria, E.; Szucs, A.; et al. International Consensus Guidelines for Risk Factors in Chronic Pancreatitis. Recommendations from the working group for the international consensus guidelines for chronic pancreatitis in collaboration with the International Association of Pancreatology, the American Pancreatic Association, the Japan Pancreas Society, and European Pancreatic Club. Pancreatology 2020, 20, 579–585. [Google Scholar] [CrossRef]
- Hegyi, E.; Sahin-Toth, M. Genetic Risk in Chronic Pancreatitis: The Trypsin-Dependent Pathway. Dig. Dis. Sci. 2017, 62, 1692–1701. [Google Scholar] [CrossRef]
- Sahin-Toth, M. Partial and complete SPINK1 deficiency cause distinct pancreatic phenotypes. Hum. Mutat. 2017, 38, 1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balazs, A.; Balla, Z.; Kui, B.; Maleth, J.; Rakonczay, Z., Jr.; Duerr, J.; Zhou-Suckow, Z.; Schatterny, J.; Sendler, M.; Mayerle, J.; et al. Ductal Mucus Obstruction and Reduced Fluid Secretion Are Early Defects in Chronic Pancreatitis. Front. Physiol. 2018, 9, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venglovecz, V.; Pallagi, P.; Kemeny, L.V.; Balazs, A.; Balla, Z.; Becskehazi, E.; Gal, E.; Toth, E.; Zvara, A.; Puskas, L.G.; et al. The Importance of Aquaporin 1 in Pancreatitis and Its Relation to the CFTR Cl(-) Channel. Front. Physiol. 2018, 9, 854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleeff, J.; Whitcomb, D.C.; Shimosegawa, T.; Esposito, I.; Lerch, M.M.; Gress, T.; Mayerle, J.; Drewes, A.M.; Rebours, V.; Akisik, F.; et al. Chronic pancreatitis. Nat. Rev. Dis. Primers 2017, 3, 17060. [Google Scholar] [CrossRef]
- Peng, C.; Li, Z.; Yu, X. The Role of Pancreatic Infiltrating Innate Immune Cells in Acute Pancreatitis. Int. J. Med. Sci. 2021, 18, 534–545. [Google Scholar] [CrossRef]
- Yang, Z.W.; Meng, X.X.; Xu, P. Central role of neutrophil in the pathogenesis of severe acute pancreatitis. J. Cell Mol. Med. 2015, 19, 2513–2520. [Google Scholar] [CrossRef] [PubMed]
- Apte, M.V.; Pirola, R.C.; Wilson, J.S. Pancreatic stellate cells: A starring role in normal and diseased pancreas. Front. Physiol. 2012, 3, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Haas De Mello, A.; Szczesny, B.; Toro, G.; Marcatti, M.; Druzhyna, N.; Liaudet, L.; Tarantini, S.; Salomao, R.; Garcia Soriano, F.; et al. Effects of the Poly(ADP-Ribose) Polymerase Inhibitor Olaparib in Cerulein-Induced Pancreatitis. Shock 2020, 53, 653–665. [Google Scholar] [CrossRef]
- Martinez-Bosch, N.; Fernandez-Zapico, M.E.; Navarro, P.; Yelamos, J. Poly(ADP-Ribose) Polymerases: New Players in the Pathogenesis of Exocrine Pancreatic Diseases. Am. J. Pathol. 2016, 186, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Mota, R.A.; Sanchez-Bueno, F.; Saenz, L.; Hernandez-Espinosa, D.; Jimeno, J.; Tornel, P.L.; Martinez-Torrano, A.; Ramirez, P.; Parrilla, P.; Yelamos, J. Inhibition of poly(ADP-ribose) polymerase attenuates the severity of acute pancreatitis and associated lung injury. Lab. Invest. 2005, 85, 1250–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdelyi, K.; Bakondi, E.; Gergely, P.; Szabo, C.; Virag, L. Pathophysiologic role of oxidative stress-induced poly(ADP-ribose) polymerase-1 activation: Focus on cell death and transcriptional regulation. Cell. Mol. Life Sci. 2005, 62, 751–759. [Google Scholar] [CrossRef]
- Virag, L.; Robaszkiewicz, A.; Rodriguez-Vargas, J.M.; Oliver, F.J. Poly(ADP-ribose) signaling in cell death. Mol. Asp. Med. 2013, 34, 1153–1167. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Quesada, R.; Munoz-Gamez, J.A.; Martin-Oliva, D.; Peralta-Leal, A.; Quiles-Perez, R.; Rodriguez-Vargas, J.M.; Ruiz de Almodovar, M.; Conde, C.; Ruiz-Extremera, A.; Oliver, F.J. Modulation of transcription by PARP-1: Consequences in carcinogenesis and inflammation. Curr. Med. Chem. 2007, 14, 1179–1187. [Google Scholar] [CrossRef] [Green Version]
- Berger, N.A.; Besson, V.C.; Boulares, A.H.; Burkle, A.; Chiarugi, A.; Clark, R.S.; Curtin, N.J.; Cuzzocrea, S.; Dawson, T.M.; Dawson, V.L.; et al. Opportunities for the repurposing of PARP inhibitors for the therapy of non-oncological diseases. Br. J. Pharmacol. 2018, 175, 192–222. [Google Scholar] [CrossRef]
- Ni, S.Y.; Zhong, X.L.; Li, Z.H.; Huang, D.J.; Xu, W.T.; Zhou, Y.; Ou, C.W.; Chen, M.S. Puerarin Alleviates Lipopolysaccharide-Induced Myocardial Fibrosis by Inhibiting PARP-1 to Prevent HMGB1-Mediated TLR4-NF-kappaB Signaling Pathway. Cardiovasc. Toxicol. 2020, 20, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Siavashpour, A.; Khalvati, B.; Azarpira, N.; Mohammadi, H.; Niknahad, H.; Heidari, R. Poly (ADP-Ribose) polymerase-1 (PARP-1) overactivity plays a pathogenic role in bile acids-induced nephrotoxicity in cholestatic rats. Toxicol. Lett. 2020, 330, 144–158. [Google Scholar] [CrossRef]
- Mota, R.; Sanchez-Bueno, F.; Berenguer-Pina, J.J.; Hernandez-Espinosa, D.; Parrilla, P.; Yelamos, J. Therapeutic treatment with poly(ADP-ribose) polymerase inhibitors attenuates the severity of acute pancreatitis and associated liver and lung injury. Br. J. Pharmacol. 2007, 151, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, P.; Petersen, O.H. The exocrine pancreas: The acinar-ductal tango in physiology and pathophysiology. Rev. Physiol. Biochem. Pharmacol. 2013, 165, 1–30. [Google Scholar] [CrossRef]
- Abu-El-Haija, M.; Gukovskaya, A.S.; Andersen, D.K.; Gardner, T.B.; Hegyi, P.; Pandol, S.J.; Papachristou, G.I.; Saluja, A.K.; Singh, V.K.; Uc, A.; et al. Accelerating the Drug Delivery Pipeline for Acute and Chronic Pancreatitis: Summary of the Working Group on Drug Development and Trials in Acute Pancreatitis at the National Institute of Diabetes and Digestive and Kidney Diseases Workshop. Pancreas 2018, 47, 1185–1192. [Google Scholar] [CrossRef]
- Manohar, M.; Verma, A.K.; Venkateshaiah, S.U.; Sanders, N.L.; Mishra, A. Chronic Pancreatitis Associated Acute Respiratory Failure. MOJ Immunol. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Balazs, A.; Hegyi, P. Cystic fibrosis-style changes in the early phase of pancreatitis. Clin. Res. Hepatol. Gastroenterol. 2015, 39 (Suppl. 1), S12–S17. [Google Scholar] [CrossRef]
- Yang, H.; Hua, C.; Yang, X.; Fan, X.; Song, H.; Peng, L.; Ci, X. Pterostilbene prevents LPS-induced early pulmonary fibrosis by suppressing oxidative stress, inflammation and apoptosis in vivo. Food Funct. 2020, 11, 4471–4484. [Google Scholar] [CrossRef]
- Wang, S.; Shi, X.L.; Feng, M.; Wang, X.; Zhang, Z.H.; Zhao, X.; Han, B.; Ma, H.C.; Dai, B.; Ding, Y.T. Puerarin protects against CCl4-induced liver fibrosis in mice: Possible role of PARP-1 inhibition. Int. Immunopharmacol. 2016, 38, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Sgambellone, S.; Lanzi, C.; Nardini, P.; Pini, A.; Moroni, F.; Masini, E.; Lucarini, L. Effects of PARP-1 Deficiency and Histamine H4 Receptor Inhibition in an Inflammatory Model of Lung Fibrosis in Mice. Front. Pharmacol. 2019, 10, 525. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Zhang, F.; Zhang, C.; Deng, S.; Wang, R.; Zhang, Y.; Huang, D.; Huang, K. Inhibition of PARP prevents angiotensin II-induced aortic fibrosis in rats. Int. J. Cardiol. 2013, 167, 2285–2293. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, Y.; Wang, L.; Zhang, F.; Deng, S.; Wang, R.; Zhang, Y.; Huang, K. Poly(ADP-ribose) polymerase 1 is indispensable for transforming growth factor-beta Induced Smad3 activation in vascular smooth muscle cell. PLoS ONE 2011, 6, e27123. [Google Scholar] [CrossRef] [Green Version]
- Lucarini, L.; Durante, M.; Lanzi, C.; Pini, A.; Boccalini, G.; Calosi, L.; Moroni, F.; Masini, E.; Mannaioni, G. HYDAMTIQ, a selective PARP-1 inhibitor, improves bleomycin-induced lung fibrosis by dampening the TGF-beta/SMAD signalling pathway. J. Cell. Mol. Med. 2017, 21, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Wahlberg, E.; Karlberg, T.; Kouznetsova, E.; Markova, N.; Macchiarulo, A.; Thorsell, A.G.; Pol, E.; Frostell, A.; Ekblad, T.; Oncu, D.; et al. Family-wide chemical profiling and structural analysis of PARP and tankyrase inhibitors. Nat. Biotechnol. 2012, 30, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cao, Y.; Staloch, D.A.; Gonzales, M.A.; Aronson, J.F.; Chao, C.; Hellmich, M.R.; Ko, T.C. Bone morphogenetic protein signaling protects against cerulein-induced pancreatic fibrosis. PLoS ONE 2014, 9, e89114. [Google Scholar] [CrossRef] [Green Version]
- Meldgaard, M.; Bollen, P.J.; Finsen, B. Non-invasive method for sampling and extraction of mouse DNA for PCR. Lab. Anim. 2004, 38, 413–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakondi, E.; Singh, S.B.; Hajnady, Z.; Nagy-Penzes, M.; Regdon, Z.; Kovacs, K.; Hegedus, C.; Madacsy, T.; Maleth, J.; Hegyi, P.; et al. Spilanthol Inhibits Inflammatory Transcription Factors and iNOS Expression in Macrophages and Exerts Anti-inflammatory Effects in Dermatitis and Pancreatitis. Int. J. Mol. Sci. 2019, 20, 4308. [Google Scholar] [CrossRef] [Green Version]
- Nathan, J.D.; Romac, J.; Peng, R.Y.; Peyton, M.; Rockey, D.C.; Liddle, R.A. Protection against chronic pancreatitis and pancreatic fibrosis in mice overexpressing pancreatic secretory trypsin inhibitor. Pancreas 2010, 39, e24–e30. [Google Scholar] [CrossRef] [Green Version]
- Regdon, Z.; Demeny, M.A.; Kovacs, K.; Hajnady, Z.; Nagy-Penzes, M.; Bakondi, E.; Kiss, A.; Hegedus, C.; Virag, L. High-content screening identifies inhibitors of oxidative stress-induced parthanatos: Cytoprotective and anti-inflammatory effects of ciclopirox. Br. J. Pharmacol. 2020. [Google Scholar] [CrossRef]
- Manohar, M.; Verma, A.K.; Venkateshaiah, S.U.; Mishra, A. Role of eosinophils in the initiation and progression of pancreatitis pathogenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 314, G211–G222. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | CERU | OLA + CERU | |
|---|---|---|---|
| Inflammatory infiltrate | 0.2 ± 0.2 | 2.6 ± 0.25 | 1.4 ± 0.25 |
| Acinar atrophy | 0.0 ± 0.0 | 2.4 ± 0.25 | 0.6 ± 0.25 |
| Fibrosis (intralobular) | 0.0 ± 0.0 | 2.6 ± 0.25 | 0.8 ± 0.37 |
| Fibrosis (interlobular) | 0.2 ± 0.2 | 2.8 ± 0.20 | 1.2 ± 0.20 |
| Fibrosis (perilobular) | 0.0 ± 0.0 | 2.6 ± 0.25 | 0.8 ± 0.20 |
| WT-CTL | WT-CERU | KO-CTL | KO-CERU | |
|---|---|---|---|---|
| Inflammatory infiltrate | 0.2 ± 0.2 | 2.6 ± 0.25 | 0 ± 0.0 | 0.6 ± 0.25 |
| Acinar atrophy | 0.0 ± 0.0 | 2.4 ± 0.25 | 0.0 ± 0.0 | 0.2 ± 0.2 |
| Fibrosis (intralobular) | 0.0 ± 0.0 | 2.6 ± 0.25 | 0.0 ± 0.0 | 0.2 ± 0.2 |
| Fibrosis (interlobular) | 0.2 ± 0.2 | 2.8 ± 0.20 | 0.2 ± 0.20 | 0.4 ± 0.2 |
| Fibrosis (perilobular) | 0.2 ± 0.2 | 2.6 ± 0.25 | 0.2 ± 0.2 | 0.2 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Hamoly, T.; Hajnády, Z.; Nagy-Pénzes, M.; Bakondi, E.; Regdon, Z.; Demény, M.A.; Kovács, K.; Hegedűs, C.; Abd El-Rahman, S.S.; Szabó, É.; et al. Poly(ADP-Ribose) Polymerase 1 Promotes Inflammation and Fibrosis in a Mouse Model of Chronic Pancreatitis. Int. J. Mol. Sci. 2021, 22, 3593. https://doi.org/10.3390/ijms22073593
El-Hamoly T, Hajnády Z, Nagy-Pénzes M, Bakondi E, Regdon Z, Demény MA, Kovács K, Hegedűs C, Abd El-Rahman SS, Szabó É, et al. Poly(ADP-Ribose) Polymerase 1 Promotes Inflammation and Fibrosis in a Mouse Model of Chronic Pancreatitis. International Journal of Molecular Sciences. 2021; 22(7):3593. https://doi.org/10.3390/ijms22073593
Chicago/Turabian StyleEl-Hamoly, Tarek, Zoltán Hajnády, Máté Nagy-Pénzes, Edina Bakondi, Zsolt Regdon, Máté A. Demény, Katalin Kovács, Csaba Hegedűs, Sahar S. Abd El-Rahman, Éva Szabó, and et al. 2021. "Poly(ADP-Ribose) Polymerase 1 Promotes Inflammation and Fibrosis in a Mouse Model of Chronic Pancreatitis" International Journal of Molecular Sciences 22, no. 7: 3593. https://doi.org/10.3390/ijms22073593