
Hypersensitivity Induced by Intrathecal Bradykinin Administration Is Enhanced by N-oleoyldopamine (OLDA) and Prevented by TRPV1 Antagonist


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
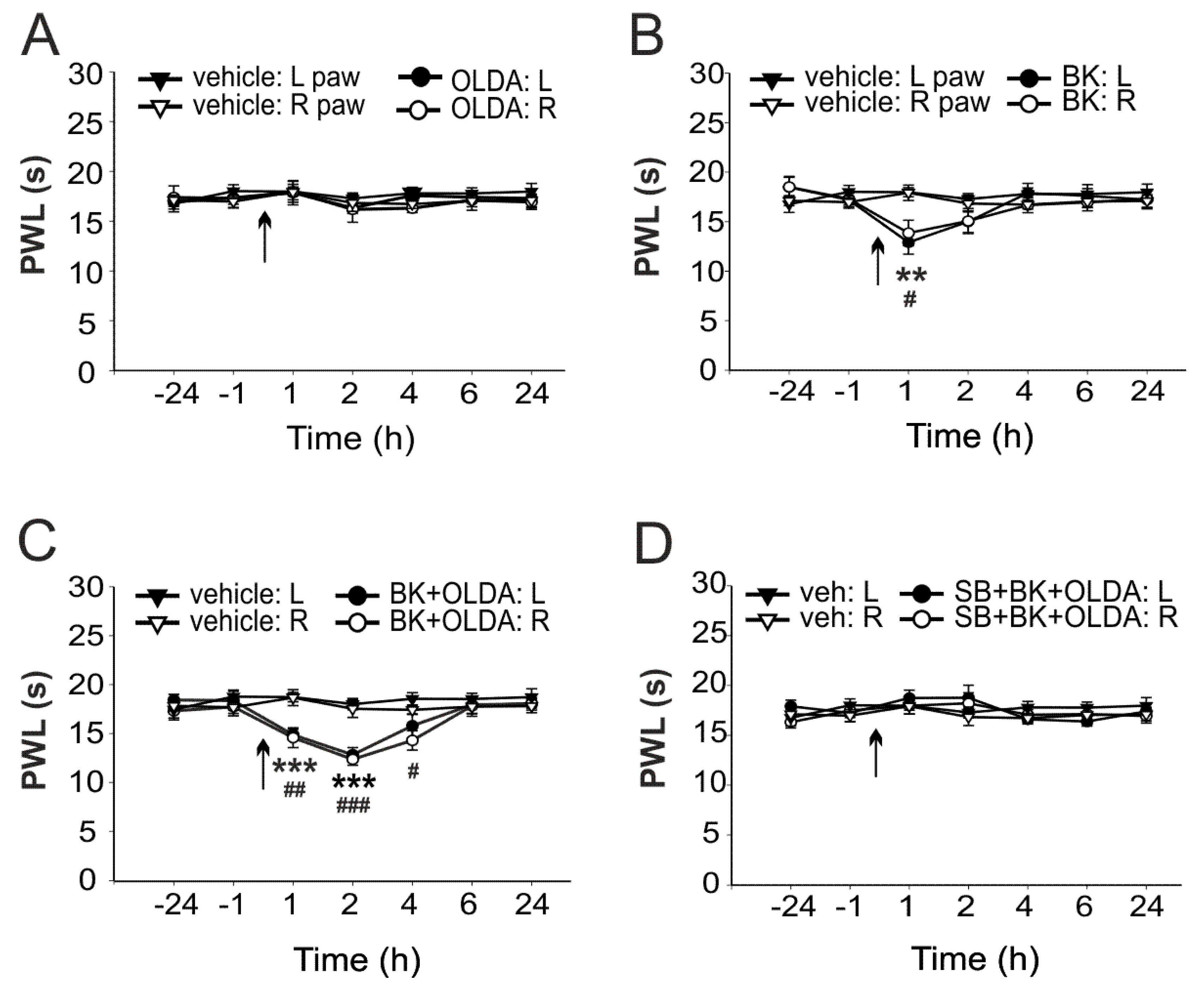
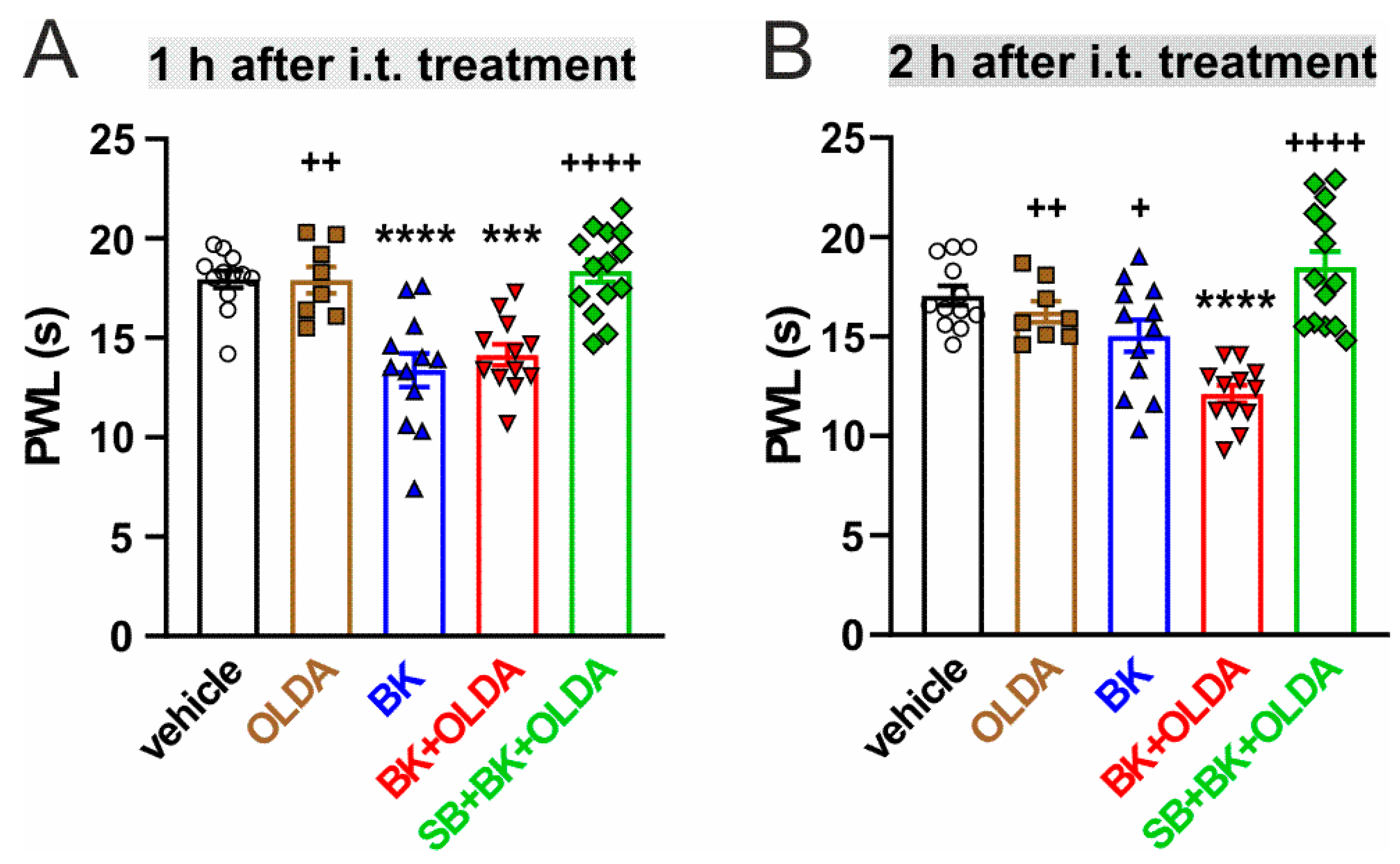
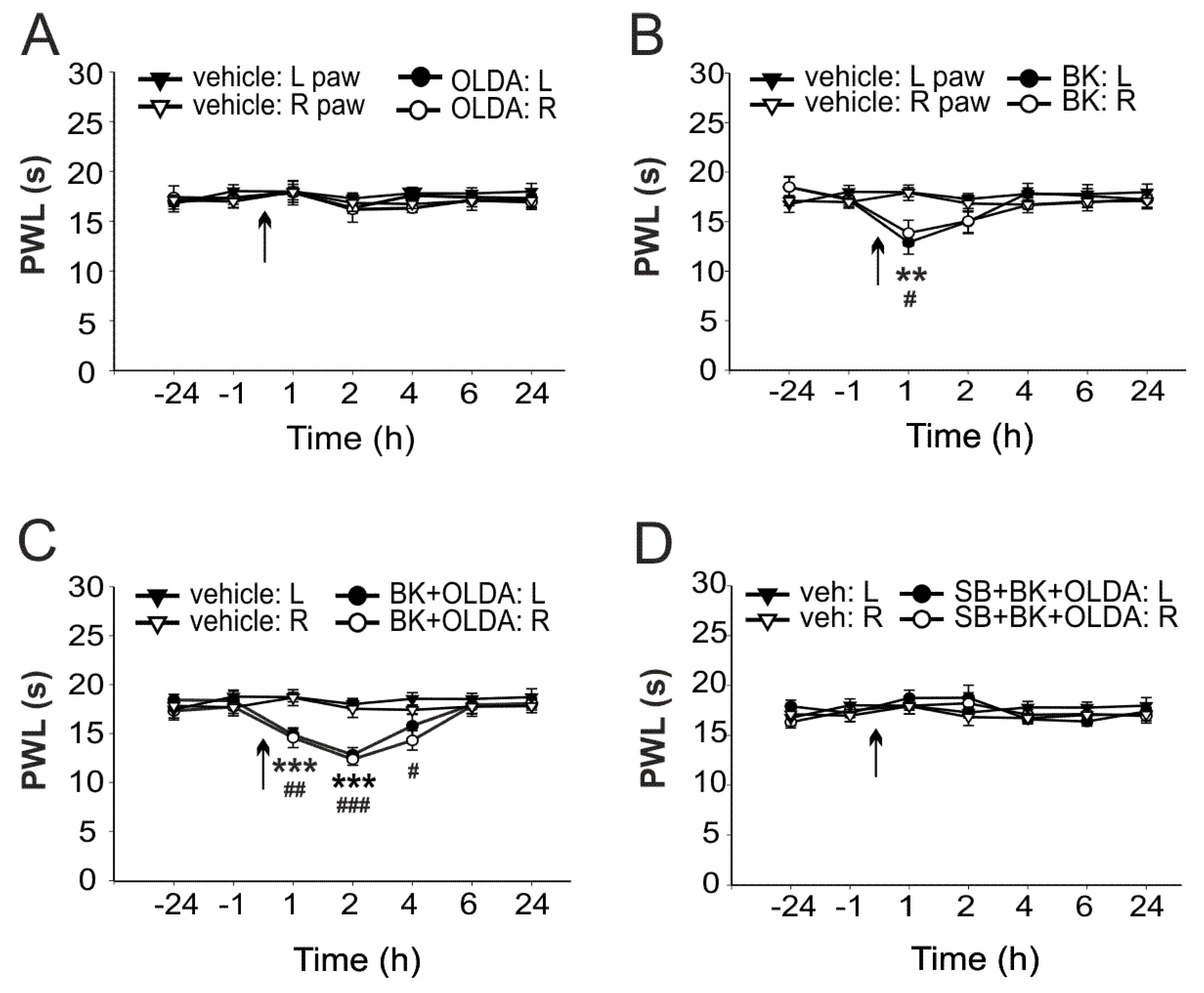
2.1. The Role of Spinal TRPV1 in the Thermal Hyperalgesia Induced by i.t. Bradykinin Treatment
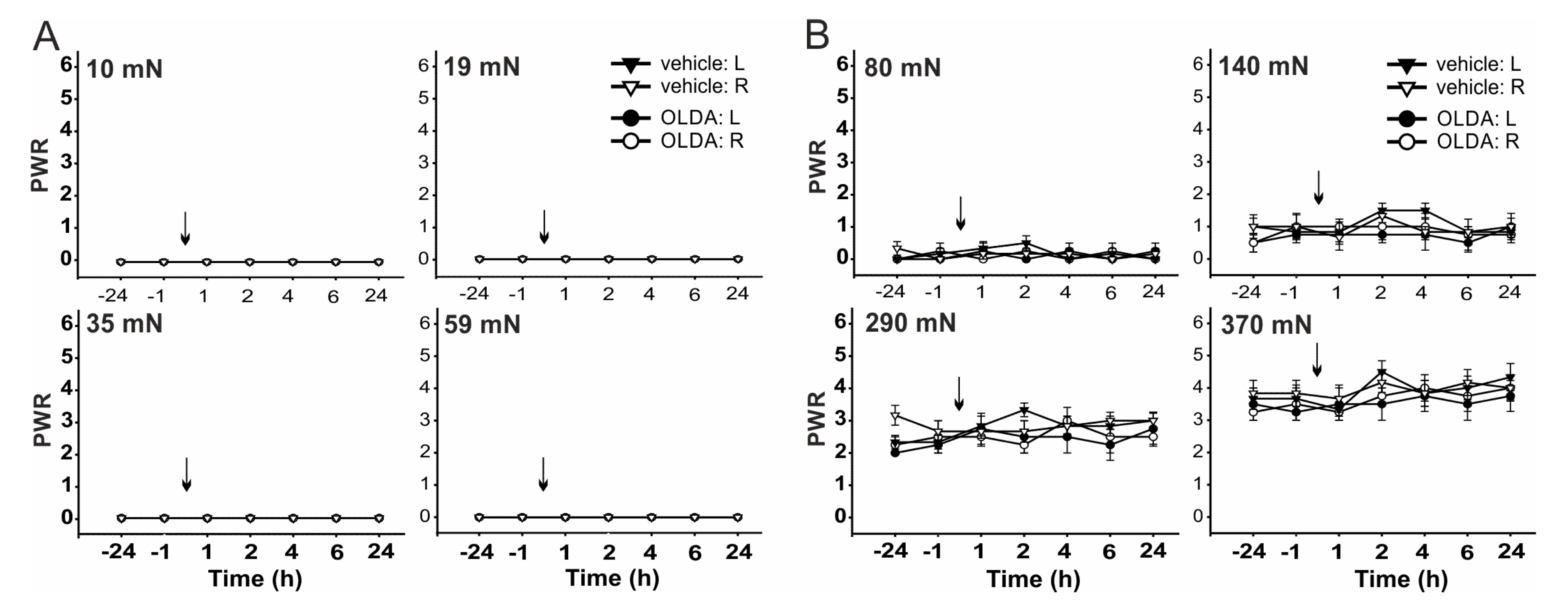
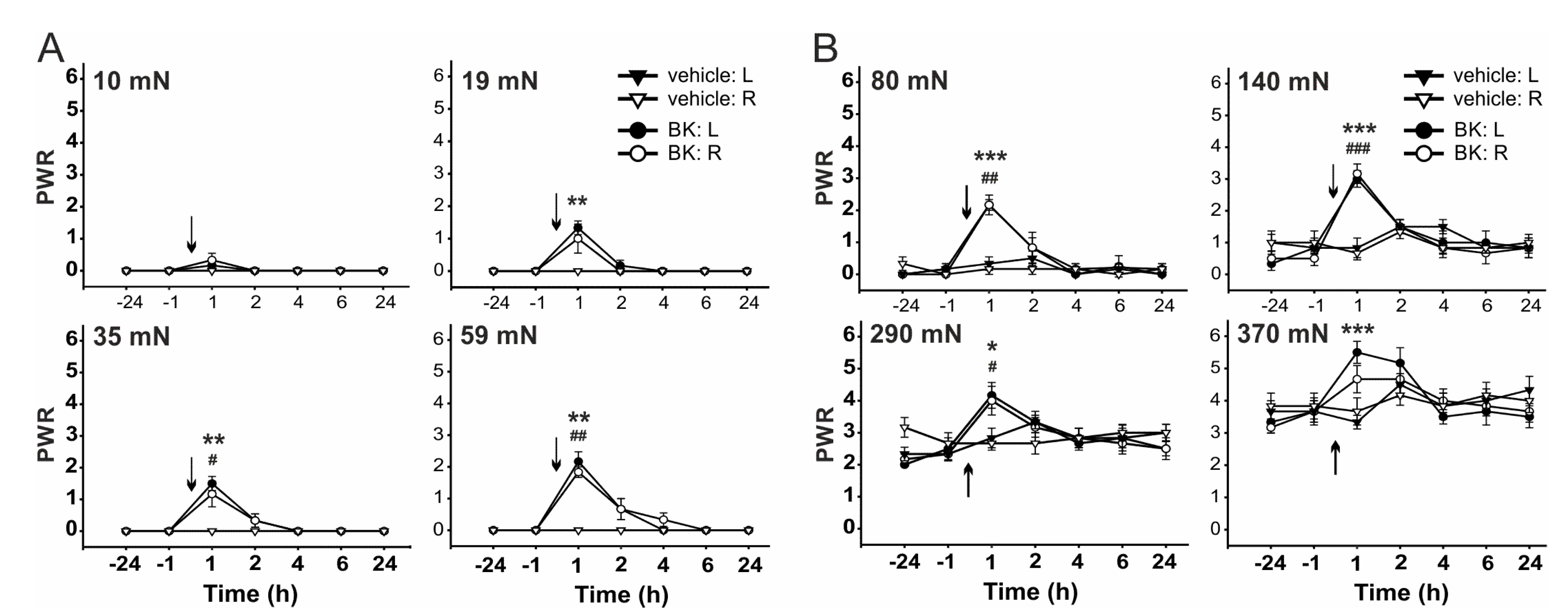
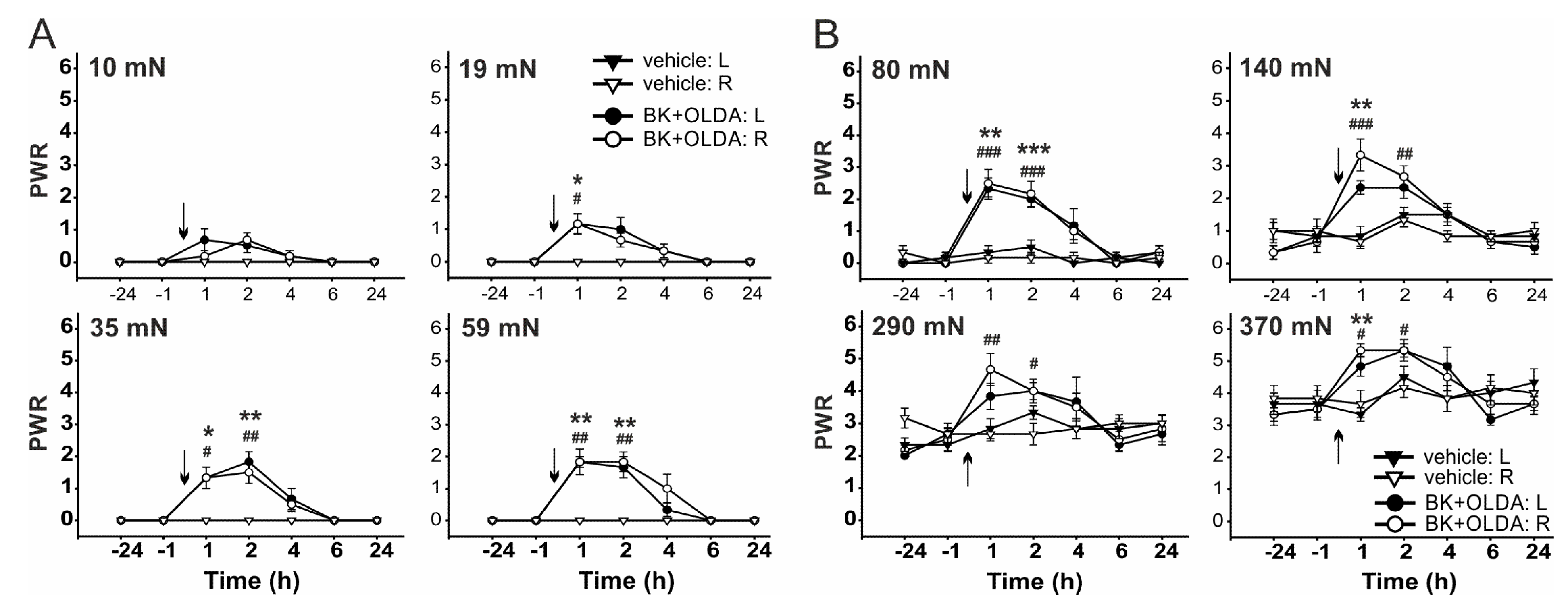
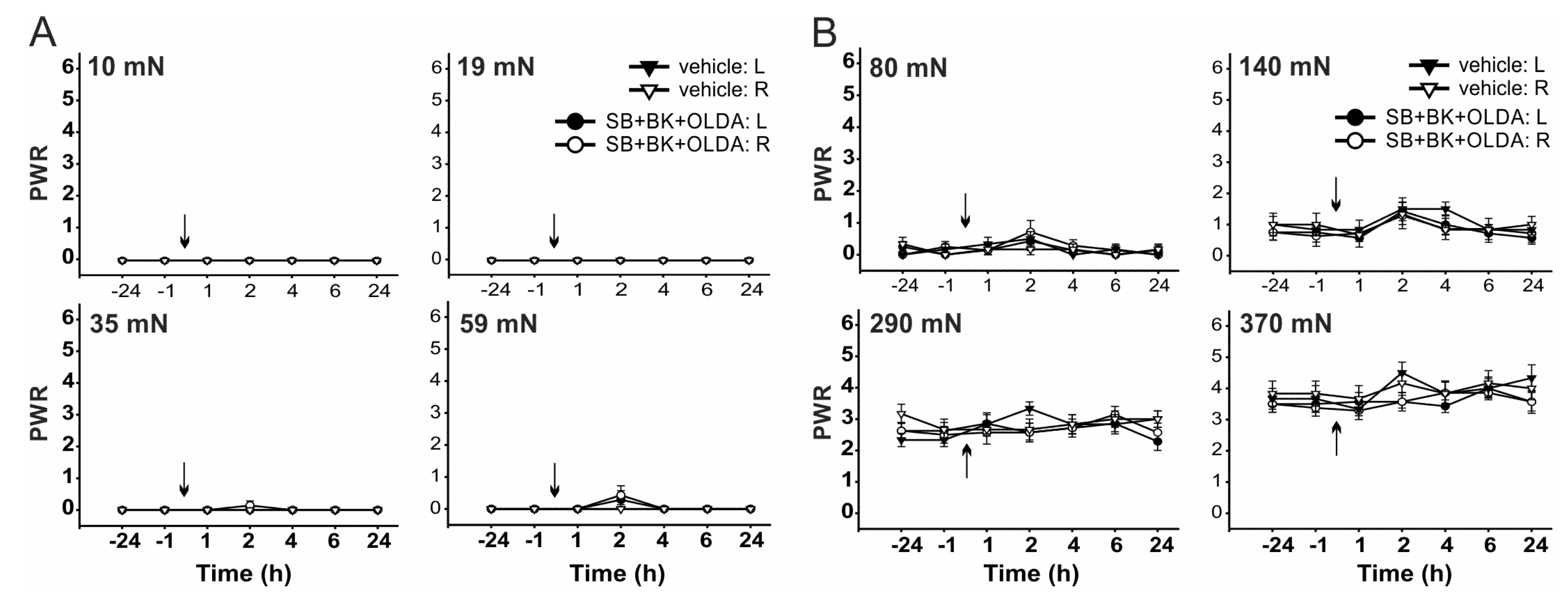
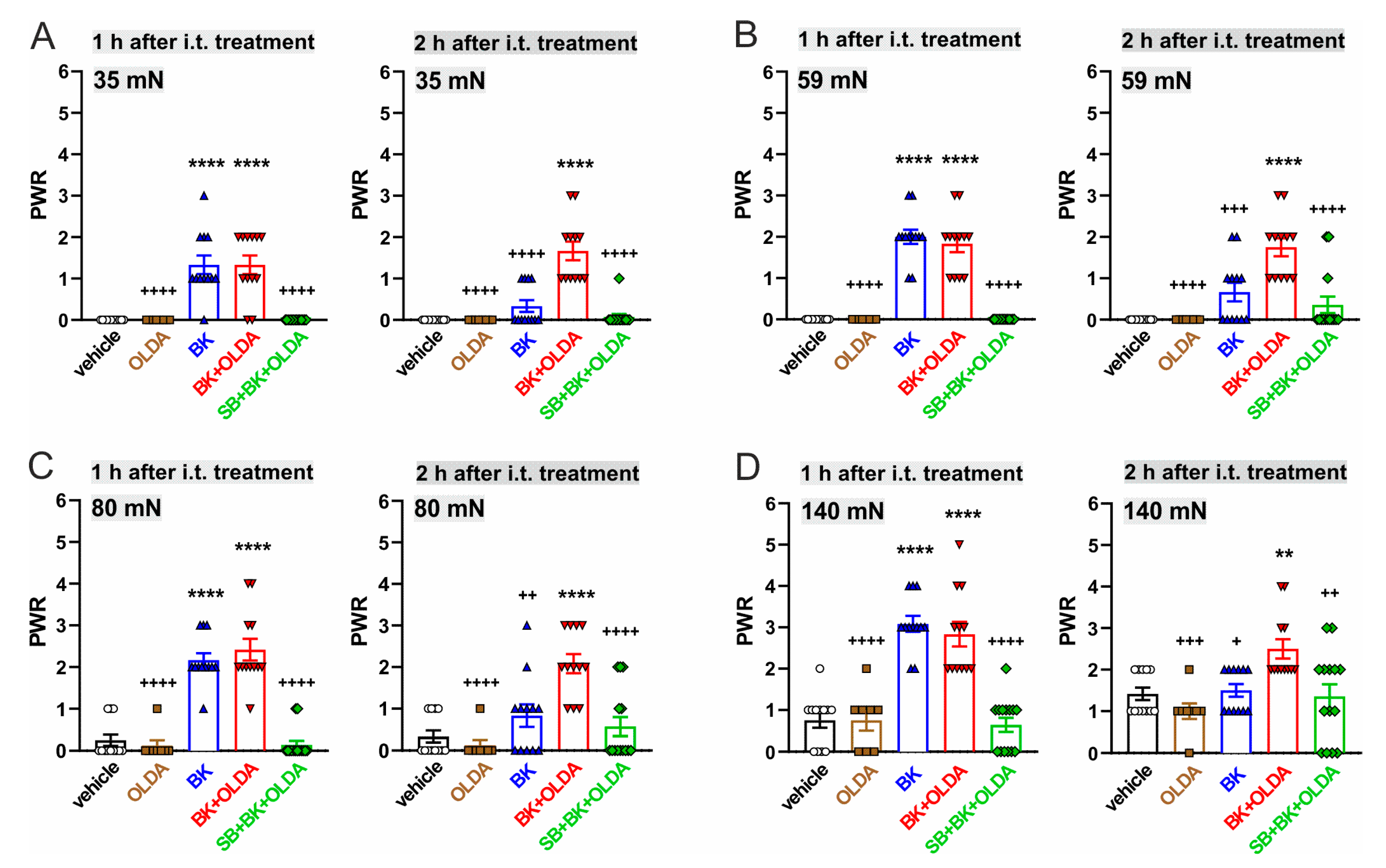
2.2. The Role of TRPV1 in the Mechanical Hypersensitivity Induced by i.t. Bradykinin Treatment
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals
4.3. Intrathecal Catheter Implantation
4.4. Behavioral Tests Procedures
4.5. Drugs
4.6. Experimental Groups
4.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mou, J.; Paillard, F.; Turnbull, B.; Trudeau, J.; Stoker, M.; Katz, N.P. Efficacy of Qutenza(R) (capsaicin) 8% patch for neuropathic pain: A meta-analysis of the Qutenza Clinical Trials Database. Pain 2013, 154, 1632–1639. [Google Scholar] [CrossRef]
- Schumacher, M.; Pasvankas, G. Topical capsaicin formulations in the management of neuropathic pain. Prog. Drug Res. 2014, 68, 105–128. [Google Scholar] [PubMed]
- Smith, H.; Brooks, J.R. Capsaicin-based therapies for pain control. Prog. Drug Res. 2014, 68, 129–146. [Google Scholar] [PubMed]
- Pospisilova, E.; Palecek, J. Post-operative pain behavior in rats is reduced after single high-concentration capsaicin application. Pain 2006, 125, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Uchytilova, E.; Spicarova, D.; Palecek, J. TRPV1 antagonist attenuates postoperative hypersensitivity by central and peripheral mechanisms. Mol. Pain 2014, 10, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, M.M.; Szallasi, A. Targeting nociceptive transient receptor potential channels to treat chronic pain: Current state of the field. Br. J. Pharmacol. 2018, 175, 2185–2203. [Google Scholar] [CrossRef]
- Treede, R.D.; Wagner, T.; Kern, K.U.; Husstedt, I.W.; Arendt, G.; Birklein, F.; Cegla, T.; Freynhagen, R.; Gockel, H.H.; Heskamp, M.L.; et al. Mechanism- and experience-based strategies to optimize treatment response to the capsaicin 8% cutaneous patch in patients with localized neuropathic pain. Curr. Med. Res. Opin. 2013, 29, 527–538. [Google Scholar] [CrossRef]
- Anand, P.; Bley, K. Topical capsaicin for pain management: Therapeutic potential and mechanisms of action of the new high-concentration capsaicin 8% patch. Br. J. Anaesth. 2011, 107, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Iadarola, M.J.; Mannes, A.J. The vanilloid agonist resiniferatoxin for interventional-based pain control. Curr. Top. Med. Chem. 2011, 11, 2171–2179. [Google Scholar] [CrossRef] [Green Version]
- Spicarova, D.; Nerandzic, V.; Palecek, J. Update on the role of spinal cord TRPV1 receptors in pain modulation. Physiol. Res. 2014, 63 (Suppl. S1), S225–S236. [Google Scholar] [CrossRef]
- Spicarova, D.; Nerandzic, V.; Palecek, J. Modulation of spinal cord synaptic activity by tumor necrosis factor alpha in a model of peripheral neuropathy. J. Neuroinflamm. 2011, 8, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spicarova, D.; Adamek, P.; Kalynovska, N.; Mrozkova, P.; Palecek, J. TRPV1 receptor inhibition decreases CCL2-induced hyperalgesia. Neuropharmacology 2014, 81, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Nerandzic, V.; Mrozkova, P.; Adamek, P.; Spicarova, D.; Nagy, I.; Palecek, J. Peripheral inflammation affects modulation of nociceptive synaptic transmission in the spinal cord induced by N-arachidonoylphosphatidylethanolamine. Br. J. Pharmacol. 2018, 175, 2322–2336. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Adamek, P.; Zhang, H.; Tatsui, C.E.; Rhines, L.D.; Mrozkova, P.; Li, Q.; Kosturakis, A.K.; Cassidy, R.M.; Harrison, D.S.; et al. The Cancer Chemotherapeutic Paclitaxel Increases Human and Rodent Sensory Neuron Responses to TRPV1 by Activation of TLR4. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 13487–13500. [Google Scholar] [CrossRef] [PubMed]
- Adamek, P.; Heles, M.; Palecek, J. Mechanical allodynia and enhanced responses to capsaicin are mediated by PI3K in a paclitaxel model of peripheral neuropathy. Neuropharmacology 2019, 146, 163–174. [Google Scholar] [CrossRef]
- Mrozkova, P.; Spicarova, D.; Palecek, J. Spinal PAR2 Activation Contributes to Hypersensitivity Induced by Peripheral Inflammation in Rats. Int. J. Mol. Sci. 2021, 22, 991. [Google Scholar] [CrossRef]
- Szolcsanyi, J.; Pinter, E. Transient receptor potential vanilloid 1 as a therapeutic target in analgesia. Expert Opin. Ther. Targets 2013, 17, 641–657. [Google Scholar] [CrossRef]
- Mrozkova, P.; Spicarova, D.; Palecek, J. Hypersensitivity Induced by Activation of Spinal Cord PAR2 Receptors Is Partially Mediated by TRPV1 Receptors. PLoS ONE 2016, 11, e0163991. [Google Scholar] [CrossRef] [Green Version]
- Spicarova, D.; Palecek, J. Tumor necrosis factor alpha sensitizes spinal cord TRPV1 receptors to the endogenous agonist N-oleoyldopamine. J. Neuroinflamm. 2010, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Spicarova, D.; Palecek, J. The role of the TRPV1 endogenous agonist N-Oleoyldopamine in modulation of nociceptive signaling at the spinal cord level. J. Neurophysiol. 2009, 102, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Couture, R.; Harrisson, M.; Vianna, R.M.; Cloutier, F. Kinin receptors in pain and inflammation. Eur. J. Pharmacol. 2001, 429, 161–176. [Google Scholar] [CrossRef]
- Lopes, P.; Kar, S.; Chretien, L.; Regoli, D.; Quirion, R.; Couture, R. Quantitative autoradiographic localization of [125I-Tyr8]bradykinin receptor binding sites in the rat spinal cord: Effects of neonatal capsaicin, noradrenergic deafferentation, dorsal rhizotomy and peripheral axotomy. Neuroscience 1995, 68, 867–881. [Google Scholar] [CrossRef]
- Wang, H.; Kohno, T.; Amaya, F.; Brenner, G.J.; Ito, N.; Allchorne, A.; Ji, R.R.; Woolf, C.J. Bradykinin produces pain hypersensitivity by potentiating spinal cord glutamatergic synaptic transmission. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 7986–7992. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.; Wotherspoon, G.; McNair, K.; Hudson, L.; Patel, S.; Gentry, C.; Winter, J. Regulation and function of spinal and peripheral neuronal B1 bradykinin receptors in inflammatory mechanical hyperalgesia. Pain 2003, 104, 683–691. [Google Scholar] [CrossRef]
- Kohno, T.; Wang, H.; Amaya, F.; Brenner, G.J.; Cheng, J.K.; Ji, R.R.; Woolf, C.J. Bradykinin enhances AMPA and NMDA receptor activity in spinal cord dorsal horn neurons by activating multiple kinases to produce pain hypersensitivity. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 4533–4540. [Google Scholar] [CrossRef]
- Chuang, H.H.; Prescott, E.D.; Kong, H.; Shields, S.; Jordt, S.E.; Basbaum, A.I.; Chao, M.V.; Julius, D. Bradykinin and nerve growth factor release the capsaicin receptor from PtdIns(4,5)P2-mediated inhibition. Nature 2001, 411, 957–962. [Google Scholar] [CrossRef]
- Cao, E.; Cordero-Morales, J.F.; Liu, B.; Qin, F.; Julius, D. TRPV1 channels are intrinsically heat sensitive and negatively regulated by phosphoinositide lipids. Neuron 2013, 77, 667–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukacs, V.; Yudin, Y.; Hammond, G.R.; Sharma, E.; Fukami, K.; Rohacs, T. Distinctive changes in plasma membrane phosphoinositides underlie differential regulation of TRPV1 in nociceptive neurons. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 11451–11463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbot, S.; Dias, J.P.; Lahjouji, K.; Bogo, M.R.; Campos, M.M.; Gaudreau, P.; Couture, R. Activation of TRPV1 by capsaicin induces functional kinin B(1) receptor in rat spinal cord microglia. J. Neuroinflamm. 2012, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Cernit, V.; Senecal, J.; Othman, R.; Couture, R. Reciprocal Regulatory Interaction between TRPV1 and Kinin B1 Receptor in a Rat Neuropathic Pain Model. Int. J. Mol. Sci. 2020, 21, 821. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.J.; Huang, S.M.; De Petrocellis, L.; Bisogno, T.; Ewing, S.A.; Miller, J.D.; Zipkin, R.E.; Daddario, N.; Appendino, G.; Di Marzo, V.; et al. N-oleoyldopamine, a novel endogenous capsaicin-like lipid that produces hyperalgesia. J. Biol. Chem. 2003, 278, 13633–13639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laneuville, O.; Reader, T.A.; Couture, R. Intrathecal bradykinin acts presynaptically on spinal noradrenergic terminals to produce antinociception in the rat. Eur. J. Pharmacol. 1989, 159, 273–283. [Google Scholar] [CrossRef]
- Kariya, K.; Yamauchi, A.; Hattori, S.; Tsuda, Y.; Okada, Y. The disappearance rate of intraventricular bradykinin in the brain of the conscious rat. Biochem. Biophys. Res. Commun. 1982, 107, 1461–1466. [Google Scholar] [CrossRef]
- Retamal, J.S.; Ramirez-Garcia, P.D.; Shenoy, P.A.; Poole, D.P.; Veldhuis, N.A. Internalized GPCRs as Potential Therapeutic Targets for the Management of Pain. Front. Mol. Neurosci. 2019, 12, 273. [Google Scholar] [CrossRef]
- Zimmerman, B.; Simaan, M.; Akoume, M.Y.; Houri, N.; Chevallier, S.; Seguela, P.; Laporte, S.A. Role of ssarrestins in bradykinin B2 receptor-mediated signalling. Cell. Signal. 2011, 23, 648–659. [Google Scholar] [CrossRef]
- Shin, J.; Cho, H.; Hwang, S.W.; Jung, J.; Shin, C.Y.; Lee, S.Y.; Kim, S.H.; Lee, M.G.; Choi, Y.H.; Kim, J.; et al. Bradykinin-12-lipoxygenase-VR1 signaling pathway for inflammatory hyperalgesia. Proc. Natl. Acad. Sci. USA 2002, 99, 10150–10155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.; da Silva, G.L.; Calixto, J.B. Contribution of vanilloid receptors to the overt nociception induced by B2 kinin receptor activation in mice. Br. J. Pharmacol. 2004, 141, 787–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katanosaka, K.; Banik, R.K.; Giron, R.; Higashi, T.; Tominaga, M.; Mizumura, K. Contribution of TRPV1 to the bradykinin-evoked nociceptive behavior and excitation of cutaneous sensory neurons. Neurosci. Res. 2008, 62, 168–175. [Google Scholar] [CrossRef]
- Kollarik, M.; Undem, B.J. Activation of bronchopulmonary vagal afferent nerves with bradykinin, acid and vanilloid receptor agonists in wild-type and TRPV1-/- mice. J. Physiol. 2004, 555, 115–123. [Google Scholar] [CrossRef]
- Rong, W.; Hillsley, K.; Davis, J.B.; Hicks, G.; Winchester, W.J.; Grundy, D. Jejunal afferent nerve sensitivity in wild-type and TRPV1 knockout mice. J. Physiol. 2004, 560, 867–881. [Google Scholar] [CrossRef]
- Dickenson, A.H.; Dray, A. Selective antagonism of capsaicin by capsazepine: Evidence for a spinal receptor site in capsaicin-induced antinociception. Br. J. Pharmacol. 1991, 104, 1045–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahner, M.R.; Li, D.P.; Chen, S.R.; Pan, H.L. Cardiac vanilloid receptor 1-expressing afferent nerves and their role in the cardiogenic sympathetic reflex in rats. J. Physiol. 2003, 551, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Petho, G.; Reeh, P.W. Sensory and signaling mechanisms of bradykinin, eicosanoids, platelet-activating factor, and nitric oxide in peripheral nociceptors. Physiol. Rev. 2012, 92, 1699–1775. [Google Scholar] [CrossRef] [PubMed]
- Mizumura, K.; Sugiura, T.; Katanosaka, K.; Banik, R.K.; Kozaki, Y. Excitation and sensitization of nociceptors by bradykinin: What do we know? Exp. Brain Res. 2009, 196, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, X.; McNaughton, P.A. Inflammatory pain: The cellular basis of heat hyperalgesia. Curr. Neuropharmacol. 2006, 4, 197–206. [Google Scholar] [CrossRef]
- Sugiura, T.; Tominaga, M.; Katsuya, H.; Mizumura, K. Bradykinin lowers the threshold temperature for heat activation of vanilloid receptor 1. J. Neurophysiol. 2002, 88, 544–548. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Geron, M.; Hazan, A.; Priel, A. Endogenous and Exogenous Vanilloids Evoke Disparate TRPV1 Activation to Produce Distinct Neuronal Responses. Front. Pharmacol. 2020, 11, 903. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uchytilova, E.; Spicarova, D.; Palecek, J. Hypersensitivity Induced by Intrathecal Bradykinin Administration Is Enhanced by N-oleoyldopamine (OLDA) and Prevented by TRPV1 Antagonist. Int. J. Mol. Sci. 2021, 22, 3712. https://doi.org/10.3390/ijms22073712
Uchytilova E, Spicarova D, Palecek J. Hypersensitivity Induced by Intrathecal Bradykinin Administration Is Enhanced by N-oleoyldopamine (OLDA) and Prevented by TRPV1 Antagonist. International Journal of Molecular Sciences. 2021; 22(7):3712. https://doi.org/10.3390/ijms22073712
Chicago/Turabian StyleUchytilova, Eva, Diana Spicarova, and Jiri Palecek. 2021. "Hypersensitivity Induced by Intrathecal Bradykinin Administration Is Enhanced by N-oleoyldopamine (OLDA) and Prevented by TRPV1 Antagonist" International Journal of Molecular Sciences 22, no. 7: 3712. https://doi.org/10.3390/ijms22073712
APA StyleUchytilova, E., Spicarova, D., & Palecek, J. (2021). Hypersensitivity Induced by Intrathecal Bradykinin Administration Is Enhanced by N-oleoyldopamine (OLDA) and Prevented by TRPV1 Antagonist. International Journal of Molecular Sciences, 22(7), 3712. https://doi.org/10.3390/ijms22073712