
Effect of Glucocorticoid Receptor Antagonism on Alcohol Self-Administration in Genetically-Selected Marchigian Sardinian Alcohol-Preferring and Non-Preferring Wistar Rats

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Experiment 1.1: Effect of Mifepristone on Alcohol Self-Administration in Male and Female msP and Wistar Rats
2.2. Experiment 1.2: Effect of Mifepristone on Saccharin Self-Administration in Male and Female msP and Wistar Rats
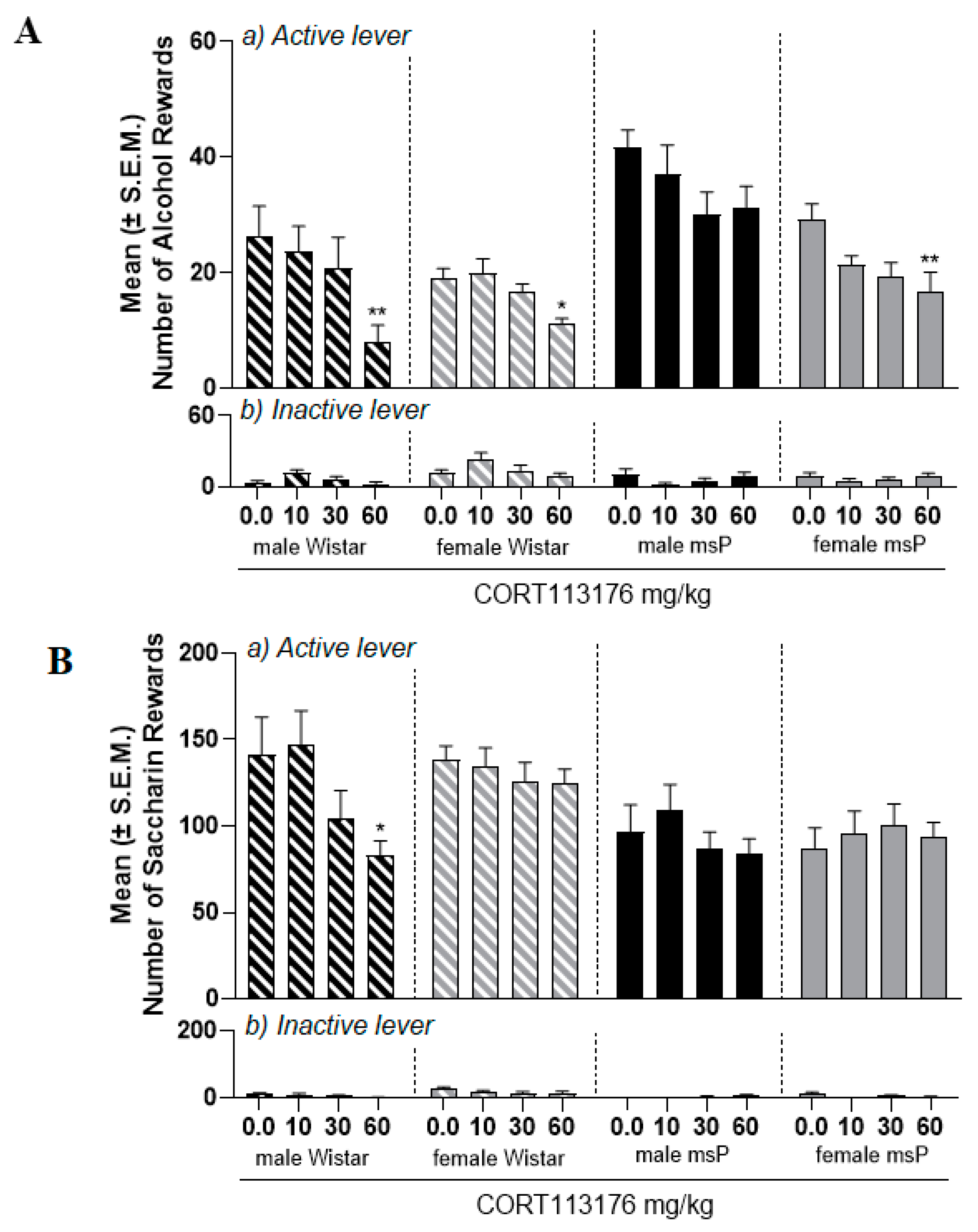
2.3. Experiment 2.1: Effect of CORT113176 on Alcohol Self-Administration in Male and Female msP and Wistar Rats
2.4. Experiment 2.2: Effect of CORT113176 on Saccharin Self-Administration in Male and Female msP and Wistar Rats
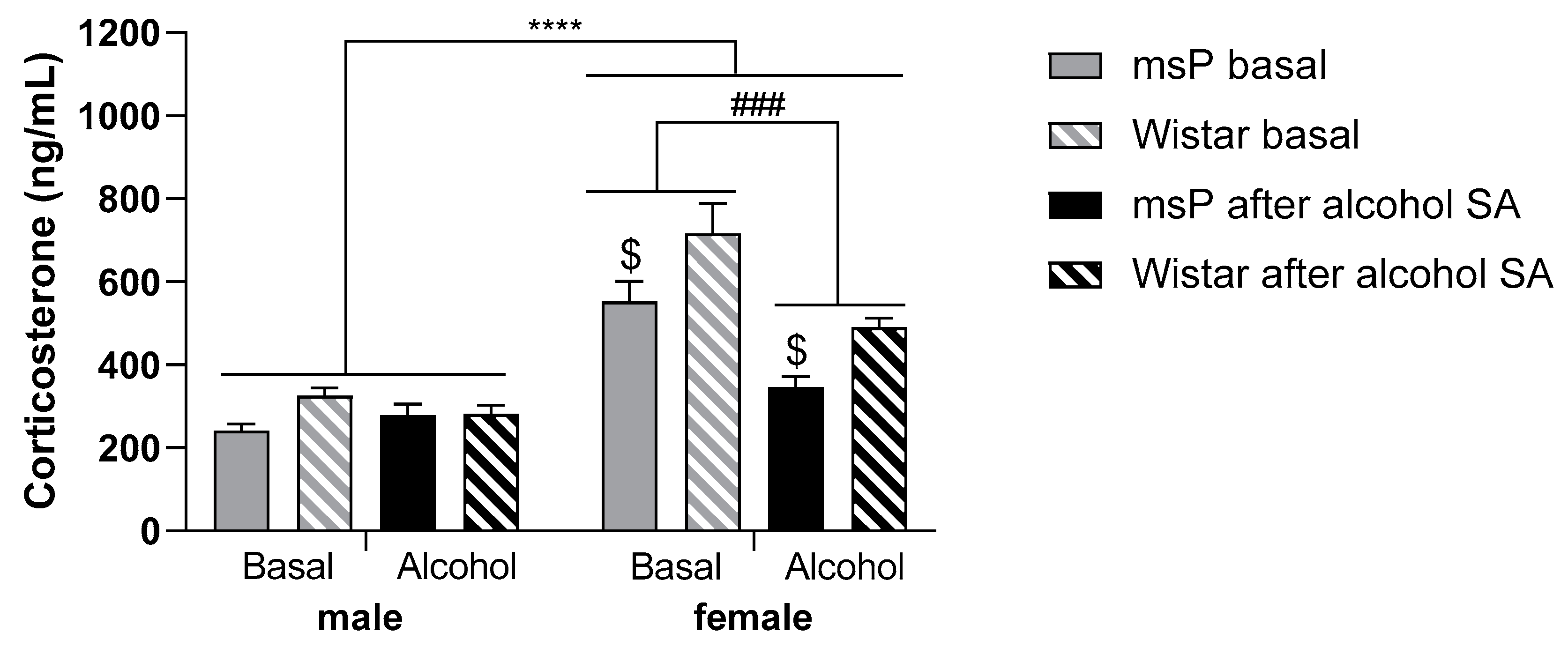
2.5. Experiment 3: Blood CORT Levels Following Alcohol Self-Administration in Male and Female msP and Wistar Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Self-Administration Apparatus
4.4. Self-Administration Training
4.5. Experiment 1.1: Effect of Mifepristone on Alcohol Self-Administration in Male and Female msP and Wistar Rats
4.6. Experiment 1.2: Effect of Mifepristone on Saccharin Self-Administration in Male and Female msP and Wistar Rats
4.7. Experiment 2.1 Effect of CORT113176 on Alcohol Self-Administration in Male and Female msP and Wistar Rats
4.8. Experiment 2.2: Effect of CORT113176 on Saccharin Self-Administration in Male and Female msP and Wistar Rats
4.9. Experiment 3: Blood Corticosterone Levels Following Alcohol Self-Administration in Male and Female msP and Wistar Rats
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koob, G.F. Theoretical Frameworks and Mechanistic Aspects of Alcohol Addiction as a Reward Deficit Disorder. Curr. Top. Behav. Neurosci. 2013, 13, 3–30. [Google Scholar]
- Witkiewitz, K.; Litten, R.Z.; Leggio, L. Advances in the Science and Treatment of Alcohol Use Disorder. Sci. Adv. 2019, 5, eaax4043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, M.A.C.; Wand, G. Stress and the HPA Axis: Role of Glucocorticoids in Alcohol Dependence. Alcohol Res. 2012, 34, 468–483. [Google Scholar]
- Edwards, S.; Little, H.J.; Richardson, H.N.; Vendruscolo, L.F. Divergent Regulation of Distinct Glucocorticoid Systems in Alcohol Dependence. Alcohol 2015, 49, 811–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.P.; Cullinan, W.E. Neurocircuitry of Stress: Central Control of the Hypothalamo–Pituitary–Adrenocortical Axis. Trends Neurosci. 1997, 20, 78–84. [Google Scholar] [CrossRef]
- Reul, J.M.H.M.; De Kloet, E.R. Two Receptor Systems for Corticosterone in Rat Brain: Microdistribution and Differential Occupation. Endocrinology 1985, 117, 2505–2511. [Google Scholar] [CrossRef] [PubMed]
- Fahlke, C.; Engel, J.A.; Eriksson, C.P.; Hȧrd, E.; Söderpalm, B. Involvement of Corticosterone in the Modulation of Ethanol Consumption in the Rat. Alcohol 1994, 11, 195–202. [Google Scholar] [CrossRef]
- Fahkle, C.; Hansen, S. Effect of Local Intracerebral Corticosterone Implants on Alcohol Intake in the Rat. Alcohol Alcohol. 1999, 34, 851–861. [Google Scholar]
- Fahkle, C.; Hard, E.; Thomasson, R.; Engel, J.A.; Hansen, S. Metyrapone-Induced Suppression of Corticosterone Synthesis re-Duces Ethanol Consumption in High-Preferring Rats. Pharm. Biochem. Behav. 1994, 48, 977–981. [Google Scholar]
- Fahkle, C.; Hard, E.; Eriksson, C.J.; Engel, J.A.; Hansen, S. Consequence of Long-Term Exposure to Corticosterone or Dexame-Thasone on Ethanol Consumption in the Adrenalectomized Rat, and the Effect of Type I and Type II Corticosteroid Receptor an-Tagonists. Psychopharmacology 1995, 117, 216–224. [Google Scholar]
- Tunstall, B.J.; A Carmack, S.; Koob, G.F.; Vendruscolo, L.F. Dysregulation of Brain Stress Systems Mediates Compulsive Alcohol Drinking. Curr. Opin. Behav. Sci. 2017, 13, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Richardson, H.N.; Lee, S.Y.; O’Dell, L.E.; Koob, G.F.; Rivier, C.L. Alcohol Self-Administration Acutely Stimulates the Hypotha-Lamic-Pituitary-Adrenal Axis, but Alcohol Dependence Leads to a Dampened Neuroendocrine State. Eur. J. Neurosci. 2008, 28, 1641–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vendruscolo, L.F.; Barbier, E.; Schlosburg, J.E.; Misra, K.K.; Whitfield, T.W., Jr.; Logrip, M.L.; Rivier, C.; Repunte-Canonigo, V.; Zorrilla, E.P.; Sanna, P.P.; et al. Corticosteroid-Dependent Plasticity Mediates Compulsive Alcohol Drinking in Rats. J. Neurosci. 2012, 30, 7563–7571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccocioppo, R.; Economidou, D.; Cippitelli, A.; Cucculelli, M.; Ubaldi, M.; Soverchia, L.; Lourdusamy, A.; Massi, M. Genetically Selected Marchigian Sardinian Alcohol-Preferring (msP) Rats: An Animal Model to Study the Neurobiology of Alcoholism. Addict. Biol. 2006, 11, 339–355. [Google Scholar] [CrossRef] [Green Version]
- Borruto, A.M.; Stopponi, S.; Li, H.; Weiss, F.; Roberto, M.; Ciccocioppo, R. Genetically Selected Alcohol-Preferring msP Rats to Study Alcohol Use Disorder: Anything Lost in Translation? Neuropharmacology 2021, 186, 108446. [Google Scholar] [CrossRef]
- Hansson, A.C.; Cippitelli, A.; Sommer, W.H.; Fedeli, A.; Bjork, K.; Soverchia, L.; Terasmaa, A.; Massi, M.; Heilig, M.; Ciccocioppo, R. Variation at the Rat Crhr1 Locus and Sensitivity to Relapse into Alcohol Seeking Induced by Environmental Stress. Proc. Natl. Acad. Sci. USA 2006, 103, 15236–15241. [Google Scholar] [CrossRef] [Green Version]
- Hansson, A.C.; Cippitelli, A.; Sommer, W.H.; Ciccocioppo, R.; Heilig, M. Region-Specific Down-Regulation of Crhr1 Gene Ex-Pression in Alcohol-Preferring msP Rats Following ad lib Access to Alcohol. Addict.Biol. 2007, 12, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Logrip, M.L.; Walker, J.R.; Ayanwuyi, L.O.; Sabino, V.; Ciccocioppo, R.; Koob, G.F.; Zorrilla, E.P. Evaluation of Alcohol Pref-erence and Drinking in msP Rats Bearing a Crhr1 Promoter Polymorphism. Front Psychiatry 2018, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Ayanwuyi, L.O.; Carvajal, F.; Lerma-Cabrera, J.M.; Domi, E.; Bjork, K.; Ubaldi, M.; Heilig, M.; Roberto, M.; Ciccocioppo, R.; Cippitelli, A. Role of a Genetic Polymorphism in the Corticotropin-Releasing Factor Receptor 1 Gene in Alcohol Drinking and Seeking Behaviors of Marchigian Sardinian Alcohol-Preferring Rats. Front. Psychiatry 2013, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Cippitelli, A.; Ayanwuyi, L.O.; Barbier, E.; Domi, E.; Lerma-Cabrera, J.M.; Carvajal, F.; Scuppa, G.; Li, H.; Ubaldi, M.; Heilig, M.; et al. Polymorphism in the Corticotropin-Releasing Factor Receptor 1 (CRF1-R) Gene Plays a Role in Shaping the High Anxious Phenotype of Marchigian Sardinian Alcohol-Preferring (msP) Rats. Psychopharmacology 2015, 232, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Blomeyer, D.; Treutlein, J.; Esser, G.; Schmidt, M.H.; Schumann, G.; Laucht, M. Interaction Between CRHR1 Gene and Stressful Life Events Predicts Adolescent Heavy Alcohol Use. Biol. Psychiatry 2008, 63, 146–151. [Google Scholar] [CrossRef]
- Treutlein, J.; Kissling, C.; Frank, J.; Wiemann, S.; Dong, L.; Depner, M.; Saam, C.; Lascorz, J.; Soyka, M.; Preuss, U.W.; et al. Genetic Association of the Human Corticotropin Releasing Hormone Receptor 1 (CRHR1) with Binge Drinking and Alcohol Intake Patterns in Two Inde-Pendent Samples. Mol. Psychiatry 2006, 11, 594–602. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.; Kallupi, M.; Luu, G.; Oleata, C.S.; Heilig, M.; Koob, G.F.; Ciccocioppo, R.; Roberto, M. Enhanced GABAergic Transmission in the Central Nucleus of the Amygdala of Genetically Selected Marchigian Sardinian Rats: Alcohol and CRF Effects. Neuropharmacology 2013, 67, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.; Varodayan, F.P.; Oleata, C.S.; Luu, G.; Kirson, D.; Heilig, M.; Ciccocioppo, R.; Roberto, M. Glutamatergic Transmission in the Central Nucleus of the Amygdala is Selectively Altered in Marchigian Sardinian Alcohol-Preferring Rats: Al-Cohol and CRF Effects. Neuropharmacology 2016, 102, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Natividad, L.A.; Buczynski, M.W.; Herman, M.A.; Kirson, D.; Oleata, C.S.; Irimia, C.; Polis, I.; Ciccocioppo, R.; Roberto, M.; Parsons, L.H. Constitutive Increases in Amygdalar Corticotropin-Releasing Factor and Fatty Acid Amide Hydrolase Drive an Anxious Phenotype. Biol. Psychiatry 2017, 82, 500–510. [Google Scholar] [CrossRef]
- Natividad, L.A.; Steinman, M.Q.; McGinn, M.A.; Sureshchandra, S.; Kerr, T.M.; Ciccocioppo, R.; Messaoudi, I.; Edwards, S.; Roberto, M. Impaired Hypothalamic Feedback Dyregulates Brain Glucocorticoid Signaling in Genetically-Selected Marchigian Sardinian Alcohol-Preferring Rats. Addict. Biol. 2021, 26, e12978. [Google Scholar] [CrossRef] [PubMed]
- Vendruscolo, L.F.; Estey, D.; Goodell, V.; Macshane, L.G.; Logrip, M.L.; Schlosburg, J.E.; McGinn, M.A.; Zamora-Martinez, E.R.; Belanoff, J.K.; Hunt, H.J.; et al. Glucocorticoid Receptor Antagonism Decreases Alcohol Seeking in Alcohol-Dependent Individuals. J. Clin. Investig. 2015, 125, 3193–3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simms, A.J.; Haass-Koffler, C.L.; Bito-Onon, J.; Li, R.; Bartlett, S.E. Mifepristone in the Central Nucleus of the Amygdala Reduces Yohimbine Stress-Induced Reinstatement of Ethanol-Seeking. Neuropsychopharmacology 2012, 37, 906–918. [Google Scholar] [CrossRef] [PubMed]
- Borruto, A.M.; Fotio, Y.; Stopponi, S.; Brunori, G.; Petrella, M.; Caputi, F.F.; Romualdi, P.; Candeletti, S.; Narendran, R.; Rorick-Kehn, L.M.; et al. NOP Receptor Antagonism Reduces Alcohol Drinking in Male and Female Rats through Mechanisms Involving the Central Amygdala and Ventral Tegmental Area. Br. J. Pharmacol. 2020, 177, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Vozella, V.; Cruz, B.; Natividad, L.; Benvenuti, F.; Cannella, N.; Edwards, S.; Zorrilla, E.; Ciccocioppo, R.; Roberto, M. Glucocorticoid Receptor Antagonist Mifepristone Does Not Alter Innate Anxiety-Like Behavior in Genetically-Selected Marchigian Sardinian (msP) Rats. Int. J. Mol. Sci. 2021, 22, 3095. [Google Scholar] [CrossRef] [PubMed]
- Peltier, M.R.; Verplaetse, T.L.; Mineur, Y.S.; Petrakis, I.L.; Cosgrove, K.P.; Picciotto, M.R.; McKee, S.A. Sex Differences in Stress-Related Alcohol Use. Neurobiol. Stress 2019, 10, 100149. [Google Scholar] [CrossRef]
- Hudson, A.; Stamp, J.A. Ovarian Hormones and Propensity to Drug Relapse: A Review. Neurosci. Biobehav. Rev. 2011, 35, 427–436. [Google Scholar] [CrossRef]
- Heck, A.L.; Handa, R.J. Sex Differences in the Hypothalamic-Pituitary-Adrenal Axis’response to Stress: An Important Role for Gonadal Hormones. Neuropsychopharmacology 2019, 44, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Koenig, H.N.; Olive, M. The Glucocorticoid Receptor Antagonist Mifepristone Reduces Ethanol Intake in Rats Under Limited Access Conditions. Psychoneuroendocrinology 2004, 29, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Savarese, A.M.; Ozburn, A.R.; Metten, P.; Schlumbohm, J.P.; Hack, W.R.; Lemoine, K.; Hunt, H.; Hausch, F.; Bauder, M.; Crabbe, J.C. Targeting the Glucocorticoid Receptor Reduces Binge-Like Drinking in High Drinking in the Dark (HDID-1) Mice. Alcohol. Clin. Exp. Res. 2020, 44, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Logrip, M.L.; Gainey, S.C. Sex Differences in the Long-Term Effects of Past Stress on Alcohol Self-Administration, Glucocorticoid Sensitivity and Phosphodiesterase 10A Expression. Neuropharmacology 2020, 164, 107857. [Google Scholar] [CrossRef] [PubMed]
- Oyola, M.G.; Handa, R.J. Hypothalamic-Pituitary-Adrenal and Hypothalamic-Pituitary-Gonadal Axes: Sex Differences in Regu-Lation of Stress Responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef] [PubMed]
- Critchlow, V.; Liebelt, R.A.; Bar-Sela, M.; Mountcastle, W.; Lipscomb, H.S. Sex Difference in Resting Pituitary-Adrenal Function in the Rat. Am. J. Physiol.-Leg. Content 1963, 205, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Rivier, C. Female Rats Release More Corticosterone than Males in Response to Alcohol: Influence of Circulating Sex Steroids and Possible Consequences for Blood Alcohol Levels. Alcohol. Clin. Exp. Res. 1993, 17, 854–859. [Google Scholar] [CrossRef]
- Ogilvie, K.M.; Rivier, C. Gender Difference in Hypothalamic-Pituitary-Adrenal Axis Response to Alcohol in the Rat: Activational Role of Gonadal Steroids. Brain Res. 1997, 766, 19–28. [Google Scholar] [CrossRef]
- Solomon, M.B.; Wulsin, A.C.; Rice, T.; Wick, D.; Myers, B.; McKlveen, J.; Flak, J.N.; Ulrich-Lai, Y.; Herman, J.P. The Selective Glucocorticoid Receptor Antagonist CORT 108297 Decreases Neuroendocrine Stress Responses and Immobility in the Forced Swim Test. Horm. Behav. 2014, 65, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulsin, A.C.; Herman, J.P.; Solomon, M.B. Mifepristone Decreases Depression-like Behavior and Modulates Neuroendocrine and Central Hypothalamic–Pituitary–Adrenocortical Axis Responsiveness to Stress. Psychoneuroendocrinology 2010, 35, 1100–1112. [Google Scholar] [CrossRef] [Green Version]
- Samson, H.H. Initiation of Ethanol Reinforcement Using a Sucrose-Substitution Procedure in Food- and Water-Sated Rats. Alcohol. Clin. Exp. Res. 1986, 10, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Vahl, T.P.; Ulrich-Lai, Y.M.; Ostrander, M.M.; Dolgas, C.M.; Elfers, E.E.; Seeley, R.J.; D’Alessio, D.A.; Herman, J.P. Comparative analysis of ACTH and Corticosterone Sampling Methods in Rats. Am. J. Physiol. Metab. 2005, 289, E823–E828. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benvenuti, F.; Cannella, N.; Stopponi, S.; Soverchia, L.; Ubaldi, M.; Lunerti, V.; Vozella, V.; Cruz, B.; Roberto, M.; Ciccocioppo, R. Effect of Glucocorticoid Receptor Antagonism on Alcohol Self-Administration in Genetically-Selected Marchigian Sardinian Alcohol-Preferring and Non-Preferring Wistar Rats. Int. J. Mol. Sci. 2021, 22, 4184. https://doi.org/10.3390/ijms22084184
Benvenuti F, Cannella N, Stopponi S, Soverchia L, Ubaldi M, Lunerti V, Vozella V, Cruz B, Roberto M, Ciccocioppo R. Effect of Glucocorticoid Receptor Antagonism on Alcohol Self-Administration in Genetically-Selected Marchigian Sardinian Alcohol-Preferring and Non-Preferring Wistar Rats. International Journal of Molecular Sciences. 2021; 22(8):4184. https://doi.org/10.3390/ijms22084184
Chicago/Turabian StyleBenvenuti, Federica, Nazzareno Cannella, Serena Stopponi, Laura Soverchia, Massimo Ubaldi, Veronica Lunerti, Valentina Vozella, Bryan Cruz, Marisa Roberto, and Roberto Ciccocioppo. 2021. "Effect of Glucocorticoid Receptor Antagonism on Alcohol Self-Administration in Genetically-Selected Marchigian Sardinian Alcohol-Preferring and Non-Preferring Wistar Rats" International Journal of Molecular Sciences 22, no. 8: 4184. https://doi.org/10.3390/ijms22084184