
Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders

Abstract
:1. Introduction
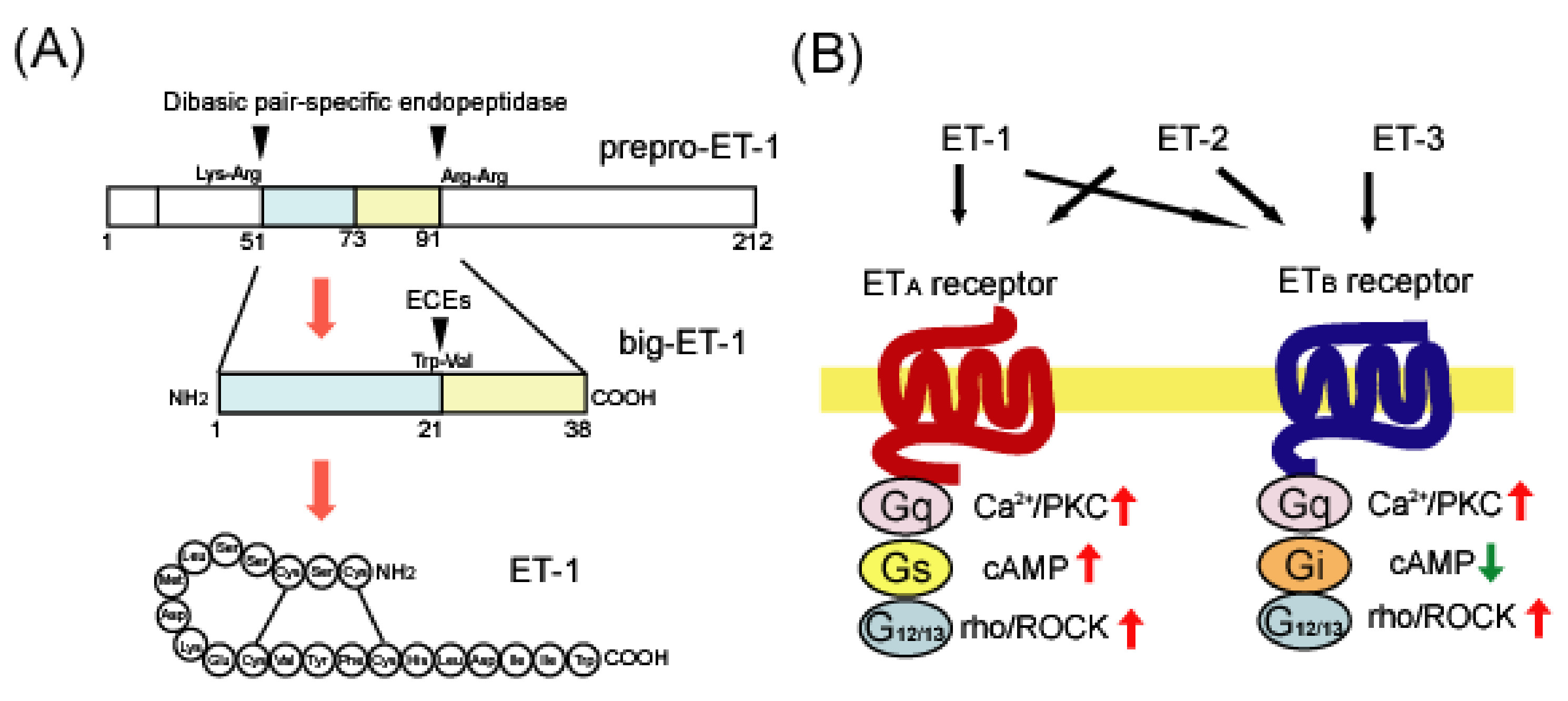
2. Overview of Endothelin
2.1. Endothelin Ligands
2.2. Endothelin Receptors
2.3. Endothelin Agonists and Antagonists
3. ET System in the Brain
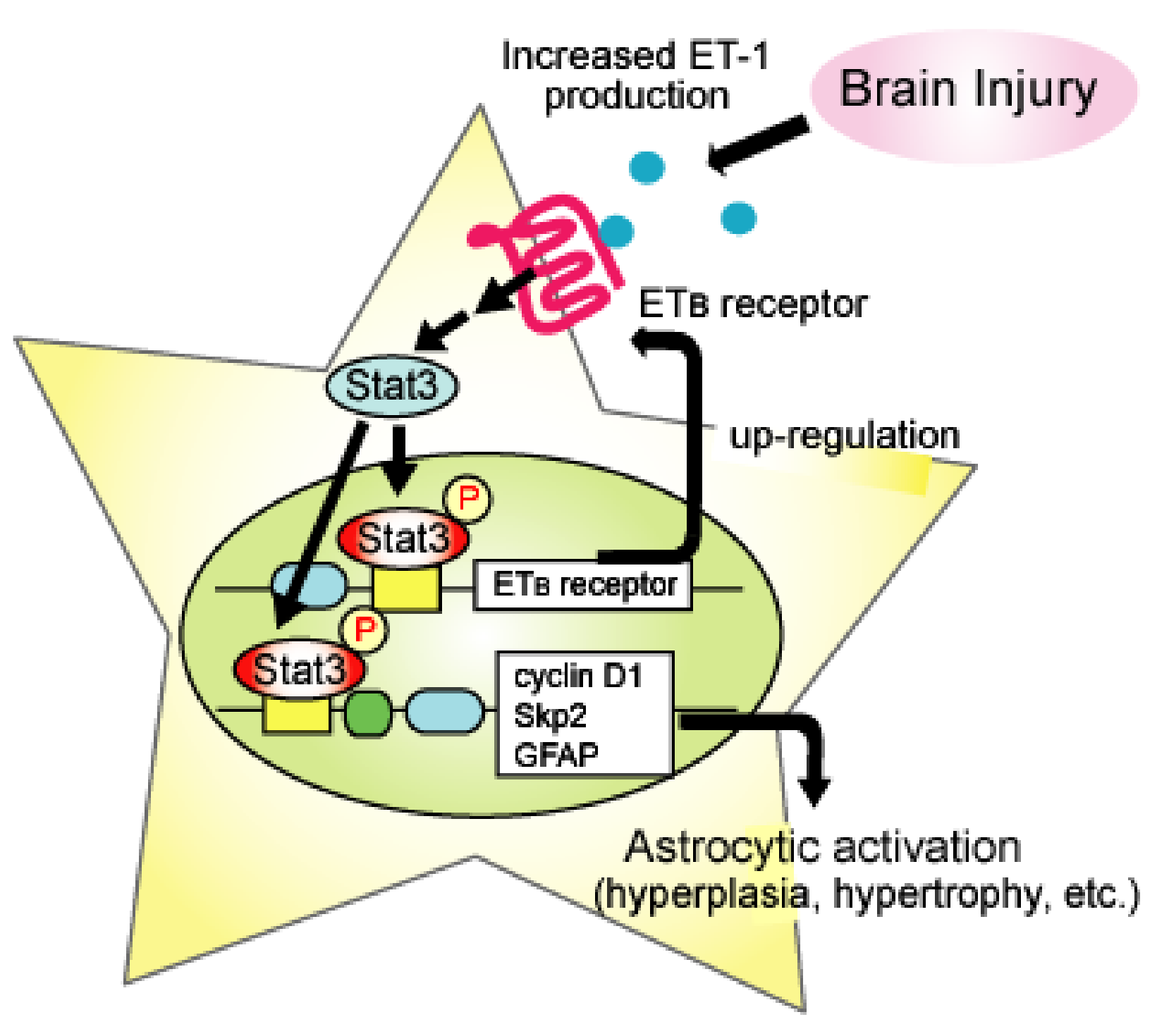
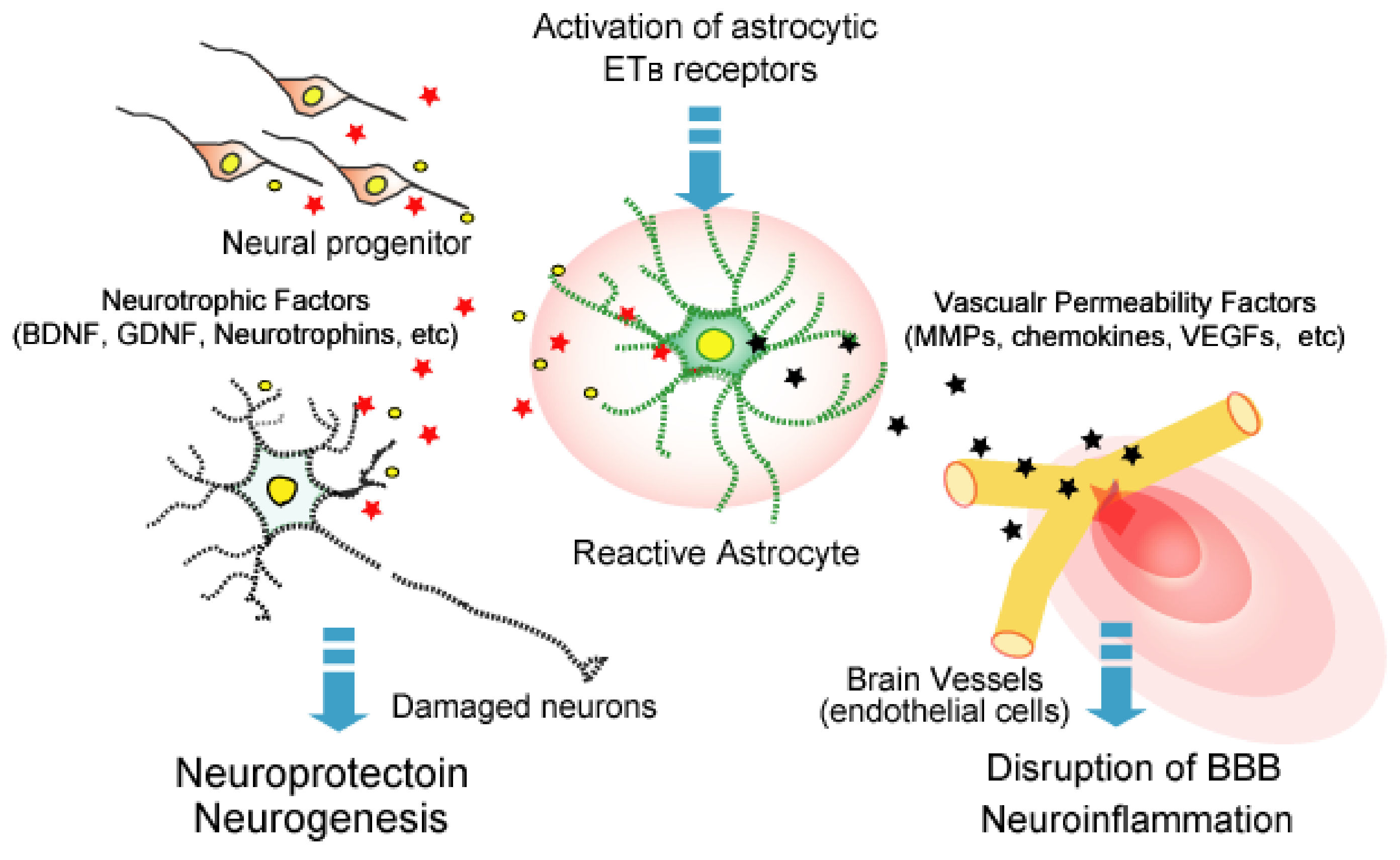
4. ETB-receptors-mediated Astrocytic Activation
5. Roles of Astrocytic ETB Receptors in Brain Disorders
5.1. Alzheimer’s Disease
5.2. Brain Ischemia
5.3. Neuropathic Pain
5.4. Traumatic Brain Injury (TBI)
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Verkhratsky, A.; Nedergaard, M. Physiology of Astroglia. Physiol. Rev. 2018, 98, 239–389. [Google Scholar] [CrossRef]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 2, 638–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddelow, S.A.; Barres, B.A. Reactive astrocytes: Production, function, and therapeutic potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostenbach, S.; Cambron, M.; D’haeseleer, M.; Kooijman, R.; De Keyser, J. Astrocyte loss and astrogliosis in neuroinflammatory disorders. Neurosci. Lett. 2014, 565, 39–41. [Google Scholar] [CrossRef]
- Colombo, E.; Farina, C. Astrocytes: Key regulators of neuroinflammation. Trends Immunol. 2016, 37, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Cassé, F.; Richetin, K.; Toni, N. Astrocytes’ contribution to adult neurogenesis in physiology and Alzheimer’s disease. Front. Cell Neurosci. 2018, 12, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lium, Z.; Chopp, M. Astrocytes, therapeutic targets for neuroprotection and neurorestoration in ischemic stroke. Prog. Neurobiol. 2016, 144, 103–120. [Google Scholar]
- Magnusson, J.P.; Göritz, C.; Tatarishvili, J.; Dias, D.O.; Smith, E.M.; Lindvall, O.; Kokaia, Z.; Frisén, J. A latent neurogenic program in astrocytes regulated by Notch signaling in the mouse. Science 2014, 346, 237–241. [Google Scholar] [CrossRef]
- Lo, F.S.; Zhao, S.; Erzurumlu, R.S. Astrocytes promote peripheral nerve injury-induced reactive synaptogenesis in the neonatal CNS. J. Neurophysiol. 2011, 106, 2876–2887. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Li, J.; Zheng, J.; Qin, S. Reactive astrocytes in neurodegenerative diseases. Aging Dis. 2019, 10, 664–675. [Google Scholar] [CrossRef] [Green Version]
- Izrael, M.; Slutsky, S.G.; Revel, M. Rising stars: Astrocytes as a therapeutic target for ALS disease. Front. Neurosci. 2020, 14, 824. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shao, A.; Yao, Y.; Tu, S.; Deng, Y.; Zhang, J. Dual roles of astrocytes in plasticity and reconstruction after traumatic brain injury. Cell Commun. Signal. 2020, 18, 62. [Google Scholar] [CrossRef] [Green Version]
- Acosta, C.; Anderson, H.D.; Anderson, C.M. Astrocyte dysfunction in Alzheimer disease. J. Neurosci. Res. 2017, 95, 2430–2447. [Google Scholar] [CrossRef] [PubMed]
- Pekny, M.; Wilhelmsson, U.; Tatlisumak, T.; Pekna, M. Astrocyte activation and reactive gliosis-A new target in stroke? Neurosci. Lett. 2019, 689, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, M.; Kurihara, H.; Kimura, S.; Tomobe, Y.; Kobayashi, M.; Mitsui, Y.; Yazaki, Y.; Goto, K.; Masaki, T. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988, 332, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Davenport, A.P.; Hyndman, K.A.; Dhaun, N.; Southan, C.; Kohan, D.E.; Pollock, J.S.; Pollock, D.M.; Webb, D.J.; Maguire, J.J. Endothelin. Pharmacol. Rev. 2016, 68, 357–418. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Mihara, S.; Ninomiya, M.; Fujimoto, M. Protective effect of endothelin type A receptor antagonist on brain edema and injury after transient middle cerebral artery occlusion in rats. Stroke 2001, 32, 2143–2148. [Google Scholar] [CrossRef] [PubMed]
- Barone, F.C.; Globus, M.Y.; Price, W.J.; White, R.F.; Storer, B.L.; Feuerstein, G.Z.; Busto, R.; Ohlstein, E.H. Endothelin levels increase in rat focal and global ischemia. J. Cereb. Blood Flow Metab. 1994, 14, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Lampl, Y.; Fleminger, G.; Gilad, R.; Galron, R.; Sarova-Pinhas, I.; Sokolovsky, M. Endothelin in cerebrospinal fluid and plasma of patients in the early stage of ischemic stroke. Stroke 1997, 28, 1951–1955. [Google Scholar] [CrossRef] [PubMed]
- Ziv, I.; Fleminger, G.; Djaldetti, R.; Achiron, A.; Melamed, E.; Sokolovsky, M. Increased plasma endothelin-1 in acute ischemic stroke. Stroke 1992, 23, 1014–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, H.; Ide, H.; Ishii, H.; Kabuto, M.; Handa, Y.; Kubota, T. Endothelin-1 levels in plasma and cerebrospinal fluid following subarachnoid haemorrhage. J. Clin. Neurosci. 1995, 2, 252–256. [Google Scholar] [CrossRef]
- Suzuki, R.; Masaoka, H.; Hirata, Y.; Marumo, F.; Isotani, E.; Hirakawa, K. The role of endotheln-1 in the origin of cerebral vasospasm in patients with aneurysmal subarachnoid hemorrhage. J. Neurosurg. 1992, 77, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Michinaga, S.; Inoue, A.; Yamamoto, H.; Ryu, R.; Inoue, A.; Mizuguchi, H.; Koyama, Y. Endothelin receptor antagonists al-leviate blood-brain barrier disruption and cerebral edema in a mouse model of traumatic brain injury: A comparison between bosentan and ambrisentan. Neuropharmacology 2020, 175, 108182. [Google Scholar] [CrossRef] [PubMed]
- Maier, B.; Lehnert, M.; Laurer, H.L.; Marzi, I. Biphasic elevation in cerebrospinal fluid and plasma concentrations of endothelin 1 after traumatic brain injury in human patients. Shock 2007, 27, 610–614. [Google Scholar] [CrossRef]
- Palmer, J.C.; Barker, R.; Kehoe, P.G.; Love, S. Endothelin-1 is elevated in Alzheimer’s disease and upregulated by amyloid-beta. J. Alzheimers Dis. 2012, 29, 853–861. [Google Scholar] [CrossRef] [Green Version]
- Ranno, E.; D’Antoni, S.; Spatuzza, M.; Berretta, A.; Laureanti, F.; Bonaccorso, C.M.; Pellitteri, R.; Longone, P.; Spalloni, A.; Iyer, A.M.; et al. Endothelin-1 is over-expressed in amyotrophic lateral sclerosis and induces motor neuron cell death. Neurobiol. Dis. 2014, 65, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Haufschild, T.; Shaw, S.G.; Kesselring, J.; Flammer, J. Increased endothelin-1 plasma levels in patients with multiple sclerosis. J. Neuroophthalmol. 2001, 21, 37–38. [Google Scholar] [CrossRef] [Green Version]
- Rolinski, B.; Heigermoser, A.; Lederer, E.; Bogner, J.R.; Loch, O.; Goebel, F.D. Endothelin-1 is elevated in the cerebrospinal fluid of HIV-infected patients with encephalopathy. Infection 1999, 27, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Hahn, S.; Gartner, S.; Pardo, C.A.; Netesan, S.K.; McArthur, J.; Nath, A. Molecular programming of endothelin-1 in HIV-infected brain: Role of Tat in up-regulation of ET-1 and its inhibition by statins. FASEB J. 2007, 21, 777–789. [Google Scholar] [CrossRef] [Green Version]
- D’Orléans-Juste, P.; Akide Ndunge, O.B.; Desbiens, L.; Tanowitz, H.B.; Desruisseaux, M.S. Endothelins in inflammatory neurological diseases. Pharmacol. Ther. 2019, 94, 145–160. [Google Scholar] [CrossRef] [PubMed]
- Armstead, W.M. Endothelins and the role of endothelin antagonists in the management of posttraumatic vasospasm. Curr. Pharm. Des. 2004, 10, 2185–2192. [Google Scholar] [CrossRef] [PubMed]
- Milan, J.; Charalambous, C.; Elhag, R.; Chen, T.C.; Li, W.; Guan, S.; Hofman, F.M.; Zidovetzki, R. Multiple signaling pathways are involved in endothelin-1-induced brain endothelial cell migration. Am. J. Physiol. Cell Physiol. 2006, 291, C155–C164. [Google Scholar] [CrossRef] [Green Version]
- Briyal, S.; Nguyen, C.; Leonard, M.; Gulati, A. Stimulation of endothelin B receptors by IRL-1620 decreases the progression of Alzheimer’s disease. Neuroscience 2015, 301, 1–11. [Google Scholar] [CrossRef]
- Adams, K.L.; Riparini, G.; Banerjee, P.; Breur, M.; Bugiani, M.; Gallo, V. Endothelin-1 signaling maintains glial progenitor proliferation in the postnatal subventricular zone. Nat. Commun. 2020, 11, 2138. [Google Scholar] [CrossRef]
- Nishikawa, K.; Ayukawa, K.; Hara, Y.; Wada, K.; Aoki, S. Endothelin/endothelin-B receptor signals regulate ventricle-directed interkinetic nuclear migration of cerebral cortical neural progenitors. Neurochem. Int. 2011, 58, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Ukita, A.; Abe, K.; Iwamae, K.; Tokuyama, S.; Tanaka, K.; Kotake, Y. Dexamethasone downregulates endothelin receptors and reduces endothelin-induced production of matrix metalloproteinases in cultured rat astrocytes. Mol. Pharmacol. 2017, 92, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Lazarini, F.; Strosberg, A.D.; Couraud, P.O.; Cazaubon, S.M. Coupling of ETB endothelin receptor to mitogen-activated protein kinase stimulation and DNA synthesis in primary cultures of rat astrocytes. J. Neurochem. 1996, 66, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, N.; Takemura, M.; Koyama, Y.; Shigenaga, Y.; Okada, T.; Baba, A. Endothelins promote the activation of astrocytes in rat neostriatum through ETB receptors. Eur. J. Neurosci. 1997, 9, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Takemura, M.; Fujiki, K.; Ishikawa, N.; Shigenaga, Y.; Baba, A. BQ788, an endothelin ETB receptor antagonist attenuates stab wound injury-induced reactive astrocytes in rat brain. Glia 1999, 26, 268–271. [Google Scholar] [CrossRef]
- Gadea, A.; Schinelli, S.; Gallo, V. Endothelin-1 regulates astrocyte proliferation and reactive gliosis via a JNK/c-Jun signaling pathway. J. Neurosci. 2008, 28, 2394–2408. [Google Scholar] [CrossRef]
- Michinaga, S.; Kimura, A.; Hatanaka, S.; Minami, S.; Asano, A.; Ikushima, Y.; Matsui, S.; Toriyama, Y.; Fujii, M.; Koyama, Y. Delayed administration of BQ788, an ETB antagonist, after experimental traumatic brain Injury promotes recovery of blood-brain barrier function and a reduction of cerebral edema in mice. J. Neurotrauma. 2018, 35, 1481–1494. [Google Scholar] [CrossRef]
- Saida, K.; Mitsui, Y.; Ishida, N. A novel peptide, vasoactive intestinal contractor, of a new (endothelin) peptide family. Molecular cloning, expression, and biological activity. J. Biol. Chem. 1989, 264, 14613–14616. [Google Scholar] [CrossRef]
- Masuo, Y.; Ishikawa, Y.; Kozakai, T.; Uchide, T.; Komatsu, Y.; Saida, K. Vasoactive intestinal contractor/endothelin-2 gene expression in the murine central nervous system. Biochem. Biophys. Res. Commun. 2003, 300, 661–668. [Google Scholar] [CrossRef]
- Uchide, T.; Masuda, H.; Lee, Y.S.; Makiyama, Y.; Mitsui, Y.; Saida, K. Fluctuating gene expression and localized cellular distribution of vasoactive intestinal contractor (VIC) in mouse uterus. J. Histochem. Cytochem. 2000, 48, 699–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, I.; Bramall, A.N.; Baynash, A.G.; Rattner, A.; Rakheja, D.; Post, M.; Joza, S.; McKerlie, C.; Stewart, D.J.; McInnes, R.R.; et al. Endothelin-2 deficiency causes growth retardation, hypothermia, and emphysema in mice. J. Clin. Investig. 2013, 123, 2643–2653. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Suzuki, N.; Onda, H.; Fujino, M. Abundance of endothelin-3 in rat intestine, pituitary gland and brain. Biochem. Biophys. Res. Commun. 1989, 164, 74–80. [Google Scholar] [CrossRef]
- Baynash, A.G.; Hosoda, K.; Giaid, A.; Richardson, J.A.; Emoto, N.; Hammer, R.E.; Yanagisawa, M. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell 1994, 79, 1277–1285. [Google Scholar] [CrossRef]
- Hofstra, R.M.; Osinga, J.; Tan-Sindhunata, G.; Wu, Y.; Kamsteeg, E.J.; Stulp, R.P.; van Ravenswaaij-Arts, C.; Majoor-Krakauer, D.; Angrist, M.; Chakravarti, A.; et al. A homozygous mutation in the endothelin-3 gene associated with a combined Waardenburg type 2 and Hirschsprung phenotype (Shah-Waardenburg syndrome). Nat. Genet. 1996, 12, 445–447. [Google Scholar] [CrossRef]
- Stow, L.R.; Jacobs, M.E.; Wingo, C.S.; Cain, B.D. Endothelin-1 gene regulation. FASEB J. 2011, 25, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Discher, D.J.; Bishopric, N.H.; Webster, K.A. Hypoxia regulates expression of the endothelin-1 gene through a proximal hypoxia-inducible factor-1 binding site on the antisense strand. Biochem. Biophys. Res. Commun. 1998, 245, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Eckman, E.A.; Reed, D.K.; Eckman, C.B. Degradation of the Alzheimer’s amyloid beta peptide by endothelin-converting enzyme. J. Biol. Chem. 2001, 276, 24540–24548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, J.C.; Kehoe, P.G.; Love, S. Endothelin-converting enzyme-1 in Alzheimer’s disease and vascular dementia. Neuropathol. Appl. Neurobiol. 2010, 36, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.; Love, S. Endothelin receptor antagonists: Potential in Alzheimer’s disease. Pharmacol. Res. 2011, 63, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Kawanabe, Y.; Okamoto, Y.; Miwa, S.; Hashimoto, N.; Masaki, T. Molecular mechanisms for the activation of voltage-independent Ca2+ channels by endothelin-1 in chinese hamster ovary cells stably expressing human endothelin-A receptors. Mol. Pharmacol. 2002, 62, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Kawanabe, Y.; Nozaki, K.; Hashimoto, N.; Masaki, T. Characterization of Ca2+ channels and G proteins involved in arachidonic acid release by endothelin-1/endothelin-A receptor. Mol. Pharmacol. 2003, 64, 689–695. [Google Scholar] [CrossRef] [Green Version]
- Jouneaux, C.; Mallat, A.; Serradeil-Le Gal, C.; Goldsmith, P.; Hanoune, J.; Lotersztajn, S. Coupling of endothelin B receptors to the calcium pump and phospholipase C via Gs and Gq in rat liver. J. Biol. Chem. 1994, 269, 1845–1851. [Google Scholar] [CrossRef]
- Eguchi, S.; Hirata, Y.; Imai, T.; Marumo, F. Endothelin receptor subtypes are coupled to adenylate cyclase via different guanyl nucleotide-binding proteins in vasculature. Endocrinology 1993, 132, 524–529. [Google Scholar] [CrossRef]
- Rauh, A.; Windischhofer, W.; Kovacevic, A.; DeVaney, T.; Huber, E.; Semlitsch, M.; Leis, H.J.; Sattler, W.; Malle, E. Endothelin (ET)-1 and ET-3 promote expression of c-fos and c-jun in human choriocarcinoma via ETB receptor-mediated Gi- and Gq-pathways and MAP kinase activation. Br. J. Pharmacol. 2008, 154, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Horinouchi, T.; Asano, H.; Higa, T.; Nishimoto, A.; Nishiya, T.; Muramatsu, I.; Miwa, S. Differential coupling of human endothelin type A receptor to Gq/11 and G12 proteins: The functional significance of receptor expression level in generating multiple receptor signaling. J. Pharmacol. Sci. 2009, 111, 338–351. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, S.; Ridley, A.J.; Lutz, S. The function of Rho-associated kinases ROCK1 and ROCK2 in the pathogenesis of cardiovascular disease. Front. Pharmacol. 2015, 6, 276. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, H.; Rashid, M. Development of Rho-kinase inhibitors for cardiovascular medicine. Trends Pharmacol. Sci. 2007, 28, 296–302. [Google Scholar] [CrossRef]
- Koyama, Y.; Baba, A. Endothelin-induced cytoskeletal actin re-organization in cultured astrocytes: Inhibition by C3 ADP-ribosyltransferase. Glia 1996, 16, 342–350. [Google Scholar] [CrossRef]
- Koyama, Y.; Yoshioka, Y.; Shinde, M.; Matsuda, T.; Baba, A. Focal adhesion kinase mediates endothelin-induced cyclin D3 expression in rat cultured astrocytes. J. Neurochem. 2004, 90, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.J.; Walker, J.W. Endothelin receptor dimers evaluated by FRET, ligand binding, and calcium mobilization. Biophys. J. 2008, 95, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, N.J.; Walker, J.W. Sustained Ca2+ signaling and delayed internalization associated with endothelin receptor heterodimers linked through a PDZ finger. Can. J. Physiol. Pharmacol. 2008, 86, 526–535. [Google Scholar] [CrossRef]
- Stanimirovic, D.B.; Ball, R.; Mealing, G.; Morley, P.; Durkin, J.P. The role of intracellular calcium and protein kinase C in endothelin-stimulated proliferation of rat type I astrocytes. Glia 1995, 15, 119–130. [Google Scholar] [CrossRef]
- Holzwarth, J.A.; Glaum, S.R.; Miller, R.J. Activation of endothelin receptors by sarafotoxin regulates Ca2+ homeostasis in cerebellar astrocytes. Glia 1992, 5, 239–250. [Google Scholar] [CrossRef]
- Koyama, Y.; Sumie, S.; Nakano, Y.; Nagao, T.; Tokumaru, S.; Michinaga, S. Endothelin-1 stimulates expression of cyclin D1 and S-phase kinase-associated protein 2 by activating the transcription factor STAT3 in cultured rat astrocytes. J. Biol. Chem. 2019, 294, 3920–3933. [Google Scholar] [CrossRef]
- Tabernero, A.; Sánchez-Alvarez, R.; Medina, J.M. Increased levels of cyclins D1 and D3 after inhibition of gap junctional communication in astrocytes. J. Neurochem. 2006, 96, 973–982. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Prasanna, G.; Yorio, T. Endothelin-1-mediated signaling in the expression of matrix metalloproteinases and tissue inhibitors of metalloproteinases in astrocytes. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3737–3745. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Tsujikawa, K.; Matsuda, T.; Baba, A. Endothelin-1 stimulates glial cell line-derived neurotrophic factor expression in cultured rat astrocytes. Biochem. Biophys. Res. Commun. 2003, 303, 1101–1105. [Google Scholar] [CrossRef]
- Koyama, Y.; Tanaka, K. Endothelins stimulate the production of stromelysin-1 in cultured rat astrocytes. Biochem. Biophys. Res. Commun. 2008, 371, 659–663. [Google Scholar] [CrossRef]
- Tencé, M.; Ezan, P.; Amigou, E.; Giaume, C. Increased interaction of connexin43 with zonula occludens-1 during inhibition of gap junctions by G protein-coupled receptor agonists. Cell Signal. 2012, 24, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Hama, H.; Sakurai, T.; Kasuya, Y.; Fujiki, M.; Masaki, T.; Goto, K. Action of endothelin-1 on rat astrocytes through the ETB receptor. Biochem. Biophys. Res. Commun. 1992, 186, 355–362. [Google Scholar] [CrossRef]
- Cazaubon, S.; Chaverot, N.; Romero, I.A.; Girault, J.A.; Adamson, P.; Strosberg, A.D.; Couraud, P.O. Growth factor activity of endothelin-1 in primary astrocytes mediated by adhesion-dependent and -independent pathways. J. Neurosci. 1997, 17, 6203–6212. [Google Scholar] [CrossRef] [Green Version]
- Ihara, M.; Noguchi, K.; Saeki, T.; Fukuroda, T.; Tsuchida, S.; Kimura, S.; Fukami, T.; Ishikawa, K.; Nishikibe, M.; Yano, M. Biological profiles of highly potent novel endothelin antagonists selective for the ETA receptor. Life Sci. 1992, 50, 247–255. [Google Scholar] [CrossRef]
- Sogabe, K.; Nirei, H.; Shoubo, M.; Nomoto, A.; Ao, S.; Notsu, Y.; Ono, T. Pharmacological profile of FR139317, a novel, potent endothelin ETA receptor antagonist. J. Pharmacol. Exp. Ther. 1993, 264, 1040–1046. [Google Scholar]
- Clozel, M.; Breu, V.; Gray, G.A.; Kalina, B.; Löffler, B.M.; Burri, K.; Cassal, J.M.; Hirth, G.; Müller, M.; Neidhart, W. Pharmacological characterization of bosentan, a new potent orally active nonpeptide endothelin receptor antagonist. J. Pharmacol. Exp. Ther. 1994, 270, 228–235. [Google Scholar]
- Vatter, H.; Seifert, V. Ambrisentan, a non-peptide endothelin receptor antagonist. Cardiovasc. Drug Rev. 2006, 24, 63–76. [Google Scholar] [CrossRef]
- Bolli, M.H.; Boss, C.; Binkert, C.; Buchmann, S.; Bur, D.; Hess, P.; Iglarz, M.; Meyer, S.; Rein, J.; Rey, M.; et al. The discovery of N-[5-(4-bromophenyl)-6-[2-[(5-bromo-2-pyrimidinyl)oxy]ethoxy]-4-pyrimidinyl]-N′-p ropylsulfamide (Macitentan), an orally active, potent dual endothelin receptor antagonist. J. Med. Chem. 2012, 55, 7849–7861. [Google Scholar] [CrossRef]
- Takai, M.; Umemura, I.; Yamasaki, K.; Watakabe, T.; Fujitani, Y.; Oda, K.; Urade, Y.; Inui, T.; Yamamura, T.; Okada, T. A potent and specific agonist, Suc-[Glu9,Ala11,15]-endothelin-1(8-21), IRL 1620, for the ETB receptor. Biochem. Biophys. Res. Commun. 1992, 184, 953–959. [Google Scholar] [CrossRef]
- Leonard, M.G.; Gulati, A. Endothelin B receptor agonist, IRL-1620, enhances angiogenesis and neurogenesis following cerebral ischemia in rats. Brain Res. 2013, 1528, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Leonard, M.G.; Briyal, S.; Gulati, A. Endothelin B receptor agonist, IRL-1620, provides long-term neuroprotection in cerebral ischemia in rats. Brain Res. 2012, 1464, 14–23. [Google Scholar] [CrossRef]
- Ishikawa, K.; Ihara, M.; Noguchi, K.; Mase, T.; Mino, N.; Saeki, T.; Fukuroda, T.; Fukami, T.; Ozaki, S.; Nagase, T. Biochemical and pharmacological profile of a potent and selective endothelin B-receptor antagonist, BQ-788. Proc. Natl. Acad. Sci. USA 1994, 91, 4892–4896. [Google Scholar] [CrossRef] [Green Version]
- Von Geldern, T.W.; Tasker, A.S.; Sorensen, B.K.; Winn, M.; Szczepankiewicz, B.G.; Dixon, D.B.; Chiou, W.J.; Wang, L.; Wessale, J.L.; Adler, A.; et al. Pyrrolidine-3-carboxylic acids as endothelin antagonists. 4. Side chain conformational restriction leads to ETB selectivity. J. Med. Chem. 1999, 42, 3668–3678. [Google Scholar] [CrossRef] [PubMed]
- Balwierczak, J.L.; Bruseo, C.W.; DelGrande, D.; Jeng, A.Y.; Savage, P.; Shetty, S.S. Characterization of a potent and selective endothelin-B receptor antagonist, IRL 2500. J. Cardiovasc. Pharmacol. 1995, 26 (Suppl. 3), S393–S396. [Google Scholar] [CrossRef]
- Nagiri, C.; Shihoya, W.; Inoue, A.; Kadji, F.M.N.; Aoki, J.; Nureki, O. Crystal structure of human endothelin ETB receptor in complex with peptide inverse agonist IRL2500. Commun. Biol. 2019, 2, 236. [Google Scholar] [CrossRef]
- Skopál, J.; Turbucz, P.; Vastag, M.; Bori, Z.; Pék, M.; deChâtel, R.; Nagy, Z.; Tóth, M.; Karádi, I. Regulation of endothelin release from human brain microvessel endothelial cells. J. Cardiovasc. Pharmacol. 1998, 31 (Suppl. 1), S370–S372. [Google Scholar] [CrossRef]
- Desai, D.; He, S.; Yorio, T.; Krishnamoorthy, R.R.; Prasanna, G. Hypoxia augments TNF-alpha-mediated endothelin-1 release and cell proliferation in human optic nerve head astrocytes. Biochem. Biophys. Res. Commun. 2004, 318, 642–648. [Google Scholar] [CrossRef]
- Ehrenreich, H.; Costa, T.; Clouse, K.A.; Pluta, R.M.; Ogino, Y.; Coligan, J.E.; Burd, P.R. Thrombin is a regulator of astrocytic endothelin-1. Brain Res. 1993, 600, 201–207. [Google Scholar] [CrossRef]
- Didier, N.; Romero, I.A.; Créminon, C.; Wijkhuisen, A.; Grassi, J.; Mabondzo, A. Secretion of interleukin-1 by astrocytes mediates endothelin-1 and tumor necrosis factor- effects on human brain microvascular endothelial cell permeability. J. Neurochem. 2003, 86, 246–254. [Google Scholar] [CrossRef]
- Sagher, O.; Jin, Y.; Thai, Q.A.; Fergus, A.; Kassell, N.F.; Lee, K.S. Cerebral microvascular responses to endothelins: The role of ETA receptors. Brain Res. 1994, 658, 179–184. [Google Scholar] [CrossRef]
- Yu, J.C.; Pickard, J.D.; Davenport, A.P. Endothelin ETA receptor expression in human cerebrovascular smooth muscle cells. Br. J. Pharmacol. 1995, 116, 2441–2446. [Google Scholar] [CrossRef] [Green Version]
- LeComte, M.D.; Shimada, I.S.; Sherwin, C.; Spees, J.L. Notch1-STAT3-ETBR signaling axis controls reactive astrocyte proliferation after brain injury. Proc. Natl. Acad. Sci. USA 2015, 112, 8726–8731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, C.M.; Rogers, S.D.; Pomonis, J.D.; Egnaczyk, G.F.; Keyser, C.P.; Schmidt, J.A.; Ghilardi, J.R.; Maggio, J.E.; Mantyh, P.W. Endothelin receptor expression in the normal and injured spinal cord: Potential involvement in injury-induced ischemia and gliosis. Exp. Neurol. 2003, 180, 1–13. [Google Scholar] [CrossRef]
- Wilhelmsson, U.; Li, L.; Pekna, M.; Berthold, C.H.; Blom, S.; Eliasson, C.; Renner, O.; Bushong, E.; Ellisman, M.; Morgan, T.E.; et al. Absence of glial fibrillary acidic protein and vimentin prevents hypertrophy of astrocytic processes and improves post-traumatic regeneration. J. Neurosci. 2004, 24, 5016–5021. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Baba, A. Endothelin-induced protein tyrosine phosphorylation of cultured astrocytes: Its relationship to cytoskeletal actin organization. Glia 1999, 26, 324–332. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, S.Y.; Cha, J.H.; Choi, Y.S.; Sung, K.W.; Oh, S.T.; Kim, O.N.; Chung, J.W.; Chun, M.H.; Lee, S.B.; et al. Upregulation of gp130 and STAT3 activation in the rat hippocampus following transient forebrain ischemia. Glia 2003, 41, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Justicia, C.; Gabriel, C.; Planas, A.M. Activation of the JAK/STAT pathway following transient focal cerebral ischemia: Signaling through Jak1 and Stat3 in astrocytes. Glia 2000, 30, 253–270. [Google Scholar] [CrossRef] [Green Version]
- Oliva, A.A., Jr.; Kang, Y.; Sanchez-Molano, J.; Furones, C.; Atkins, C.M. STAT3 signaling after traumatic brain injury. J. Neurochem. 2012, 120, 710–720. [Google Scholar] [CrossRef]
- Di Giovanni, S.; Movsesyan, V.; Ahmed, F.; Cernak, I.; Schinelli, S.; Stoica, B.; Faden, A.I. Cell cycle inhibition provides neuroprotection and reduces glial proliferation and scar formation after traumatic brain injury. Proc. Natl. Acad. Sci. USA 2005, 102, 8333–8338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Y.; Cheng, C.; Chen, Y.; Shi, S.; Qin, J.; Xiao, F.; Zhou, D.; Lu, M.; Lu, Q.; et al. A relationship between p27(kip1) and Skp2 after adult brain injury: Implications for glial proliferation. J. Neurotrauma. 2010, 27, 361–371. [Google Scholar] [CrossRef]
- Wu, J.; Pajoohesh-Ganji, A.; Stoica, B.A.; Dinizo, M.; Guanciale, K.; Faden, A.I. Delayed expression of cell cycle proteins contributes to astroglial scar formation and chronic inflammation after rat spinal cord contusion. J. Neuroinflammation. 2012, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, J.E.; Imura, T.; Song, B.; Qi, J.; Ao, Y.; Nguyen, T.K.; Korsak, R.A.; Takeda, K.; Akira, S.; Sofroniew, M.V. STAT3 is a critical regulator of astrogliosis and scar formation after spinal cord injury. J. Neurosci. 2008, 28, 7231–7243. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Nakamura, M.; Katoh, H.; Miyao, T.; Shimazaki, T.; Ishii, K.; Yamane, J.; Yoshimura, A.; Iwamoto, Y.; Toyama, Y.; et al. Conditional ablation of Stat3 or Socs3 discloses a dual role for reactive astrocytes after spinal cord injury. Nat. Med. 2006, 12, 829–834. [Google Scholar] [CrossRef]
- Hung, V.K.; Yeung, P.K.; Lai, A.K.; Ho, M.C.; Lo, A.C.; Chan, K.C.; Wu, E.X.; Chung, S.S.; Cheung, C.W.; Chung, S.K. Selective astrocytic endothelin-1 overexpression contributes to dementia associated with ischemic stroke by exaggerating astrocyte-derived amyloid secretion. J. Cereb. Blood Flow Metab. 2015, 35, 1687–1696. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.C.; Baig, S.; Kehoe, P.G.; Love, S. Endothelin-converting enzyme-2 is increased in Alzheimer’s disease and up-regulated by A. Am. J. Pathol. 2009, 175, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Quinto, J.; Eckman, E.A. Endothelin-converting enzymes degrade intracellular beta-amyloid produced within the endosomal/lysosomal pathway and autophagosomes. J. Biol. Chem. 2013, 88, 5606–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elesber, A.A.; Bonetti, P.O.; Woodrum, J.E.; Zhu, X.Y.; Lerman, L.O.; Younkin, S.G.; Lerman, A. Bosentan preserves endothelial function in mice overexpressing app. Neurobiol. Aging 2006, 27, 446–450. [Google Scholar] [CrossRef]
- Koyama, Y.; Tsujikawa, K.; Matsuda, T.; Baba, A. Endothelin increases expression of exon III- and exon IV-containing brain-derived neurotrophic factor transcripts in cultured astrocytes and rat brain. J. Neurosci. Res. 2005, 80, 809–816. [Google Scholar] [CrossRef]
- Ladenheim, R.G.; Lacroix, I.; Foignant-Chaverot, N.; Strosberg, A.D.; Couraud, P.O. Endothelins stimulate c-fos and nerve growth factor expression in astrocytes and astrocytoma. J. Neurochem. 1993, 60, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Tsujikawa, K.; Matsuda, T.; Baba, A. Intracerebroventricular administration of an endothelin ETB receptor agonist increases expressions of GDNF and BDNF in rat brain. Eur. J. Neurosci. 2003, 18, 887–894. [Google Scholar] [CrossRef]
- Khodorova, A.; Montmayeur, J.P.; Strichartz, G. Endothelin receptors and pain. J. Pain. 2009, 10, 4–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwak, Y.S.; Kang, J.; Unabia, G.C.; Hulsebosch, C.E. Spatial and temporal activation of spinal glial cells: Role of gliopathy in central neuropathic pain following spinal cord injury in rats. Exp. Neurol. 2012, 234, 362–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuda, M.; Kohro, Y.; Yano, T.; Tsujikawa, T.; Kitano, J.; Tozaki-Saitoh, H.; Koyanagi, S.; Ohdo, S.; Ji, R.R.; Salter, M.W.; et al. JAK-STAT3 pathway regulates spinal astrocyte proliferation and neuropathic pain maintenance in rats. Brain 2011, 134 Pt 4, 1127–1139. [Google Scholar] [CrossRef]
- Yamasaki, R.; Fujii, T.; Wang, B.; Masaki, K.; Kido, M.A.; Yoshida, M.; Matsushita, T.; Kira, J.I. Allergic inflammation leads to neuropathic pain via glial cell activation. J. Neurosci. 2016, 36, 11929–11945. [Google Scholar] [CrossRef] [Green Version]
- Michinaga, S.; Koyama, Y. Dual roles of astrocyte-derived factors in regulation of blood-brain barrier function after brain damage. Int. J. Mol. Sci. 2019, 20, 571. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Maebara, Y.; Hayashi, M.; Nagae, R.; Tokuyama, S.; Michinaga, S. Endothelins reciprocally regulate VEGF-A and angiopoietin-1 production in cultured rat astrocytes: Implications on astrocytic proliferation. Glia 2012, 60, 1954–1963. [Google Scholar] [CrossRef]
- Koyama, Y.; Tanaka, K. Intracerebroventricular administration of an endothelin ETB receptor agonist increases expression of matrix metalloproteinase-2 and -9 in rat brain. J. Pharmacol. Sci. 2010, 114, 433–443. [Google Scholar] [CrossRef] [Green Version]
- Michinaga, S.; Tanabe, A.; Nakaya, R.; Fukutome, C.; Inoue, A.; Iwane, A.; Minato, Y.; Tujiuchi, Y.; Miyake, D.; Mizuguchi, H.; et al. Angiopoietin-1/Tie-2 signal after focal traumatic brain injury is potentiated by BQ788, an ETB receptor antagonist, in the mouse cerebrum: Involvement in recovery of blood-brain barrier function. J. Neurochem. 2020, 154, 330–348. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Kotani, M.; Sawamura, T.; Kuribayashi, M.; Konishi, R.; Michinaga, S. Different actions of endothelin-1 on chemokine production in rat cultured astrocytes: Reduction of CX3CL1/fractalkine and an increase in CCL2/MCP-1 and CXCL1/CINC-1. J. Neuroinflamm. 2013, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Michinaga, S.; Inoue, A.; Sonoda, K.; Mizuguchi, H.; Koyama, Y. Down-regulation of astrocytic sonic hedgehog by activation of endothelin ETB receptors: Involvement in traumatic brain injury-induced disruption of blood brain barrier in a mouse model. Neurochem. Int. 2021, 7, 105042. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Tsuboi, S.; Mizogui, F. Endothelin-1 decreases the expression of ephrin-A and B subtypes in cultured rat astrocytes through ETB receptors. Neurosci. Lett. 2021, 741, 135393. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Agonist | Antagonist | |
|---|---|---|
| ET receptor non-selective | ET-1 | Bosentan, Macitentan |
| ETA selective | sarafotoxin 6b | Ambrisentan, Sitaxsentan, Atrasentan, Clazosentan, Zibotentan, S-0139, SB234551, Ro-61-1790 |
| ETB selective | sarafotoxin 6c, IRL-1620, BQ3020, Ala1,3,11,15-ET-1 | BQ788, IRL-2500, A192621, RES-701-1 |
| Neurotrophic Factors | Vascular Permeability Regulators | Others | |
|---|---|---|---|
| Up-regulation | GDNF [72], BDNF [111], NGF [112] | VEGF [119], MMP2 [120], MMP3 [73], MMP9 [71,120] ET-1 [24] | CCL2/MCP-1 [122], CXCL1/CINC-1 [122] |
| Down-regulation | angiopoietin-1 [119,121] sonic hedgehog [123] | CX3CL1/fractalkine [122], ephrin-A2, -A4, -B2, -B3 [124] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koyama, Y. Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders. Int. J. Mol. Sci. 2021, 22, 4333. https://doi.org/10.3390/ijms22094333
Koyama Y. Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders. International Journal of Molecular Sciences. 2021; 22(9):4333. https://doi.org/10.3390/ijms22094333
Chicago/Turabian StyleKoyama, Yutaka. 2021. "Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders" International Journal of Molecular Sciences 22, no. 9: 4333. https://doi.org/10.3390/ijms22094333
APA StyleKoyama, Y. (2021). Endothelin ETB Receptor-Mediated Astrocytic Activation: Pathological Roles in Brain Disorders. International Journal of Molecular Sciences, 22(9), 4333. https://doi.org/10.3390/ijms22094333