
The Distribution of GPR17-Expressing Cells Correlates with White Matter Inflammation Status in Brain Tissues of Multiple Sclerosis Patients

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. All the Histopathological Conditions of Human Brain MS Lesions Can Coexist and They Can Be Disseminated both in Grey and White Matter
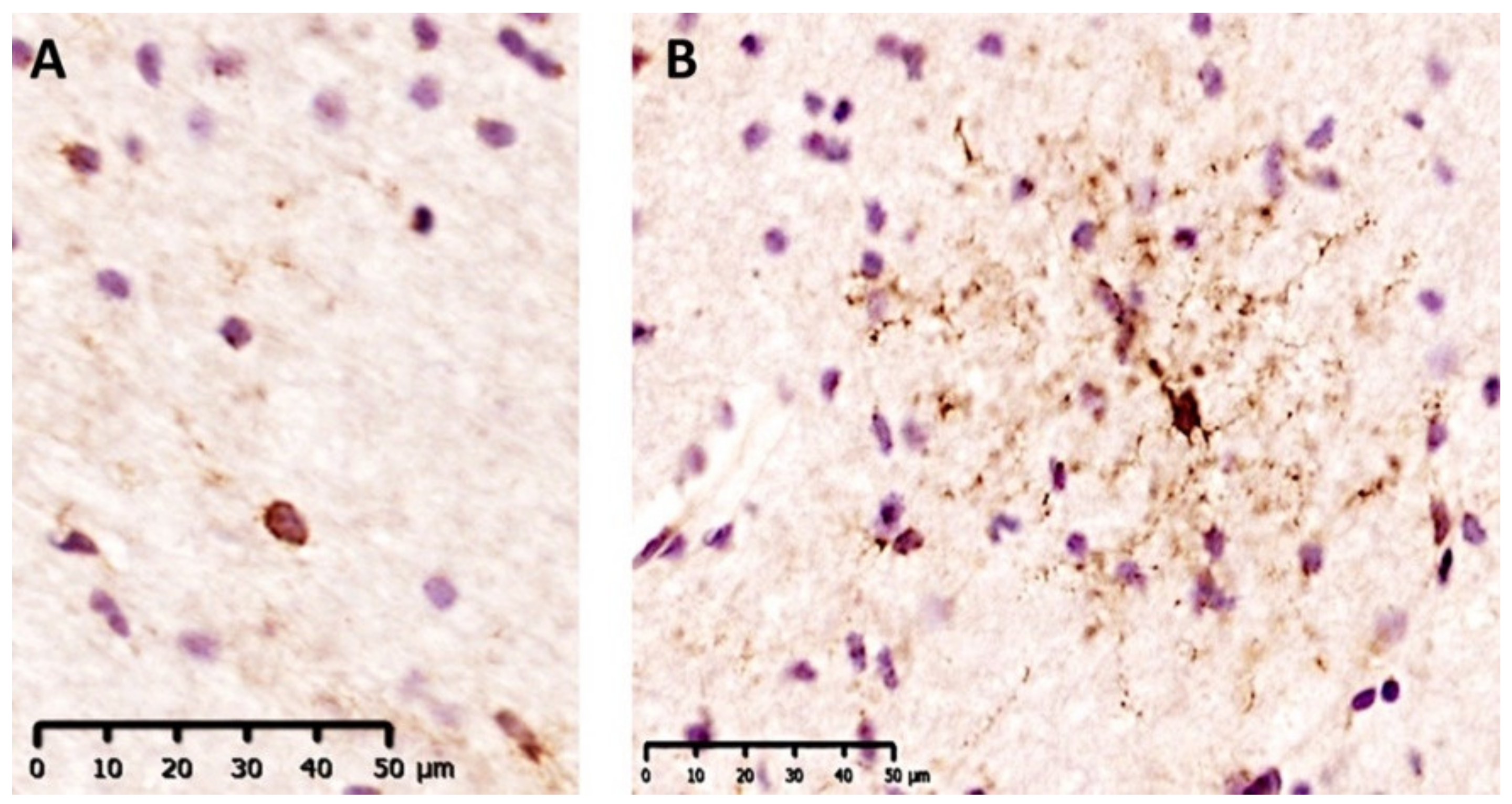
2.2. GPR17+ Cells Are More Represented in Inflamed White Matter of MS Patients and They Belong to Early Oligodendroglial Lineage
2.3. GPR17+ Cells Show Two Different Morphologies Which Are Associeted with Specific Histological Conditions
2.4. Rounded GPR17+ Cells Specifically Populate the Core of Acute Lesions
3. Discussion
4. Materials and Methods
4.1. Sample Acquisition
4.2. Immunohistochemistry on Human Tissues
4.3. Immunofluorescence on Human Tissues
4.4. Classification of Demyelinating Lesions
4.5. Descriptive and Quantitative Analyses
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Podbielska, M.; Banik, N.L.; Kurowska, E.; Hogan, E.L. Myelin Recovery in Multiple Sclerosis: The Challenge of Remyelination. Brain Sci. 2013, 3, 1282–1324. [Google Scholar] [CrossRef]
- Starost, L.; Lindner, M.; Herold, M.; Xu, Y.K.T.; Drexler, H.C.A.; Heß, K.; Ehrlich, M.; Ottoboni, L.; Ruffini, F.; Stehling, M.; et al. Extrinsic Immune Cell-Derived, but Not Intrinsic Oligodendroglial Factors Contribute to Oligodendroglial Differentiation Block in Multiple Sclerosis. Acta Neuropathol. 2020, 140, 715–736. [Google Scholar] [CrossRef]
- Rovaris, M.; Confavreux, C.; Furlan, R.; Kappos, L.; Comi, G.; Filippi, M. Secondary Progressive Multiple Sclerosis: Current Knowledge and Future Challenges. Lancet Neurol. 2006, 5, 343–354. [Google Scholar] [CrossRef]
- Lecca, D.; Raffaele, S.; Abbracchio, M.P.; Fumagalli, M. Regulation and Signaling of the GPR17 Receptor in Oligodendroglial Cells. Glia 2020. [Google Scholar] [CrossRef]
- Alavi, M.S.; Karimi, G.; Roohbakhsh, A. The Role of Orphan G Protein-Coupled Receptors in the Pathophysiology of Multiple Sclerosis: A Review. Life Sci. 2019, 224, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Lecca, D.; Trincavelli, M.L.; Gelosa, P.; Sironi, L.; Ciana, P.; Fumagalli, M.; Villa, G.; Verderio, C.; Grumelli, C.; Guerrini, U.; et al. The Recently Identified P2Y-like Receptor GPR17 Is a Sensor of Brain Damage and a New Target for Brain Repair. PLoS ONE 2008, 3, e3579. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, S.; Villa, G.; Genovese, T.; Mazzon, E.; Longhi, R.; Rosa, P.; Bramanti, P.; Cuzzocrea, S.; Abbracchio, M.P. The P2Y-like Receptor GPR17 as a Sensor of Damage and a New Potential Target in Spinal Cord Injury. Brain 2009, 132, 2206–2218. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, H.; Wang, S.; Koito, H.; Li, J.; Ye, F.; Hoang, J.; Escobar, S.S.; Gow, A.; Arnett, H.A.; et al. The Oligodendrocyte-Specific G Protein-Coupled Receptor GPR17 Is a Cell-Intrinsic Timer of Myelination. Nat. Neurosci. 2009, 12, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, M.; Daniele, S.; Lecca, D.; Lee, P.R.; Parravicini, C.; Fields, R.D.; Rosa, P.; Antonucci, F.; Verderio, C.; Trincavelli, M.L.; et al. Phenotypic Changes, Signaling Pathway, and Functional Correlates of GPR17-Expressing Neural Precursor Cells during Oligodendrocyte Differentiation. J. Biol. Chem. 2011, 286, 10593–10604. [Google Scholar] [CrossRef]
- Fumagalli, M.; Lecca, D.; Abbracchio, M.P. CNS Remyelination as a Novel Reparative Approach to Neurodegenerative Diseases: The Roles of Purinergic Signaling and the P2Y-like Receptor GPR17. Neuropharmacology 2016, 104, 82–93. [Google Scholar] [CrossRef]
- Boda, E.; Viganò, F.; Rosa, P.; Fumagalli, M.; Labat-Gest, V.; Tempia, F.; Abbracchio, M.P.; Dimou, L.; Buffo, A. The GPR17 Receptor in NG2 Expressing Cells: Focus on in Vivo Cell Maturation and Participation in Acute Trauma and Chronic Damage. Glia 2011, 59, 1958–1973. [Google Scholar] [CrossRef]
- Ciana, P.; Fumagalli, M.; Trincavelli, M.L.; Verderio, C.; Rosa, P.; Lecca, D.; Ferrario, S.; Parravicini, C.; Capra, V.; Gelosa, P.; et al. The Orphan Receptor GPR17 Identified as a New Dual Uracil Nucleotides/Cysteinyl-Leukotrienes Receptor. EMBO J. 2006, 25, 4615–4627. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, E.; Bonifacino, T.; Raffaele, S.; Milanese, M.; Morgante, E.; Bonanno, G.; Abbracchio, M.P.; Fumagalli, M. Abnormal Upregulation of GPR17 Receptor Contributes to Oligodendrocyte Dysfunction in SOD1 G93A Mice. Int. J. Mol. Sci. 2020, 21, 2395. [Google Scholar] [CrossRef]
- Coppolino, G.T.; Marangon, D.; Negri, C.; Menichetti, G.; Fumagalli, M.; Gelosa, P.; Dimou, L.; Furlan, R.; Lecca, D.; Abbracchio, M.P. Differential Local Tissue Permissiveness Influences the Final Fate of GPR17-Expressing Oligodendrocyte Precursors in Two Distinct Models of Demyelination. Glia 2018, 66, 1118–1130. [Google Scholar] [CrossRef] [PubMed]
- Paladini, M.S.; Marangon, D.; Rossetti, A.C.; Guidi, A.; Coppolino, G.T.; Negri, C.; Spero, V.; Abbracchio, M.P.; Lecca, D.; Molteni, R. Prenatal Stress Impairs Spinal Cord Oligodendrocyte Maturation via BDNF Signaling in the Experimental Autoimmune Encephalomyelitis Model of Multiple Sclerosis. Cell Mol. Neurobiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Parravicini, C.; Daniele, S.; Palazzolo, L.; Trincavelli, M.L.; Martini, C.; Zaratin, P.; Primi, R.; Coppolino, G.; Gianazza, E.; Abbracchio, M.P.; et al. A Promiscuous Recognition Mechanism between GPR17 and SDF-1: Molecular Insights. Cell. Signal. 2016, 28, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Franke, H.; Parravicini, C.; Lecca, D.; Zanier, E.R.; Heine, C.; Bremicker, K.; Fumagalli, M.; Rosa, P.; Longhi, L.; Stocchetti, N.; et al. Changes of the GPR17 Receptor, a New Target for Neurorepair, in Neurons and Glial Cells in Patients with Traumatic Brain Injury. Purinergic Signal. 2013, 9, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Jäkel, S.; Agirre, E.; Mendanha Falcão, A.; van Bruggen, D.; Lee, K.W.; Knuesel, I.; Malhotra, D.; Ffrench-Constant, C.; Williams, A.; Castelo-Branco, G. Altered Human Oligodendrocyte Heterogeneity in Multiple Sclerosis. Nature 2019, 566, 543–547. [Google Scholar] [CrossRef]
- Kuhlmann, T.; Ludwin, S.; Prat, A.; Antel, J.; Brück, W.; Lassmann, H. An Updated Histological Classification System for Multiple Sclerosis Lesions. Acta Neuropathol. 2017, 133, 13–24. [Google Scholar] [CrossRef]
- Van der Poel, M.; Ulas, T.; Mizee, M.R.; Hsiao, C.-C.; Miedema, S.S.M.; Adelia; Schuurman, K.G.; Helder, B.; Tas, S.W.; Schultze, J.L.; et al. Transcriptional Profiling of Human Microglia Reveals Grey-White Matter Heterogeneity and Multiple Sclerosis-Associated Changes. Nat. Commun. 2019, 10, 1139. [Google Scholar] [CrossRef]
- Mattiace, L.A.; Davies, P.; Dickson, D.W. Detection of HLA-DR on Microglia in the Human Brain Is a Function of Both Clinical and Technical Factors. Am. J. Pathol. 1990, 136, 1101–1114. [Google Scholar] [PubMed]
- Kutzelnigg, A.; Lucchinetti, C.F.; Stadelmann, C.; Brück, W.; Rauschka, H.; Bergmann, M.; Schmidbauer, M.; Parisi, J.E.; Lassmann, H. Cortical Demyelination and Diffuse White Matter Injury in Multiple Sclerosis. Brain 2005, 128, 2705–2712. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H. Multiple Sclerosis Pathology. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, R.; Kira, J.-I. Multiple Sclerosis. Adv. Exp. Med. Biol. 2019, 1190, 217–247. [Google Scholar] [CrossRef] [PubMed]
- Sensi, C.; Daniele, S.; Parravicini, C.; Zappelli, E.; Russo, V.; Trincavelli, M.L.; Martini, C.; Abbracchio, M.P.; Eberini, I. Oxysterols Act as Promiscuous Ligands of Class-A GPCRs: In Silico Molecular Modeling and in Vitro Validation. Cell. Signal. 2014, 26, 2614–2620. [Google Scholar] [CrossRef]
- Daniele, S.; Trincavelli, M.L.; Gabelloni, P.; Lecca, D.; Rosa, P.; Abbracchio, M.P.; Martini, C. Agonist-Induced Desensitization/Resensitization of Human G Protein-Coupled Receptor 17: A Functional Cross-Talk between Purinergic and Cysteinyl-Leukotriene Ligands. J. Pharmacol. Exp. Ther. 2011, 338, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Wolswijk, G. Oligodendrocyte Precursor Cells in the Demyelinated Multiple Sclerosis Spinal Cord. Brain 2002, 125, 338–349. [Google Scholar] [CrossRef]
- Schmidt, C.; Ohlemeyer, C.; Labrakakis, C.; Walter, T.; Kettenmann, H.; Schnitzer, J. Analysis of Motile Oligodendrocyte Precursor Cells in Vitro and in Brain Slices. Glia 1997, 20, 284–298. [Google Scholar] [CrossRef]
- Binamé, F.; Sakry, D.; Dimou, L.; Jolivel, V.; Trotter, J. NG2 Regulates Directional Migration of Oligodendrocyte Precursor Cells via Rho GTPases and Polarity Complex Proteins. J. Neurosci. 2013, 33, 10858–10874. [Google Scholar] [CrossRef]
- Bonfanti, E.; Gelosa, P.; Fumagalli, M.; Dimou, L.; Viganò, F.; Tremoli, E.; Cimino, M.; Sironi, L.; Abbracchio, M.P. The Role of Oligodendrocyte Precursor Cells Expressing the GPR17 Receptor in Brain Remodeling after Stroke. Cell Death Dis. 2017, 8, e2871. [Google Scholar] [CrossRef]
- Rone, M.B.; Cui, Q.-L.; Fang, J.; Wang, L.-C.; Zhang, J.; Khan, D.; Bedard, M.; Almazan, G.; Ludwin, S.K.; Jones, R.; et al. Oligodendrogliopathy in Multiple Sclerosis: Low Glycolytic Metabolic Rate Promotes Oligodendrocyte Survival. J. Neurosci. 2016, 36, 4698–4707. [Google Scholar] [CrossRef] [PubMed]
- Elkjaer, M.L.; Frisch, T.; Reynolds, R.; Kacprowski, T.; Burton, M.; Kruse, T.A.; Thomassen, M.; Baumbach, J.; Illes, Z. Molecular Signature of Different Lesion Types in the Brain White Matter of Patients with Progressive Multiple Sclerosis. Acta Neuropathol. Commun. 2019, 7, 205. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, K.A.; Sloane, J.A. Regulation of Remyelination in Multiple Sclerosis. FEBS Lett. 2011, 585, 3821–3828. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.S.Y.; Djelloul, M.; Steiner, E.; Bernard, S.; Salehpour, M.; Possnert, G.; Brundin, L.; Frisén, J. Dynamics of Oligodendrocyte Generation in Multiple Sclerosis. Nature 2019, 566, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Calderon, T.M.; Eugenin, E.A.; Lopez, L.; Kumar, S.S.; Hesselgesser, J.; Raine, C.S.; Berman, J.W. A Role for CXCL12 (SDF-1alpha) in the Pathogenesis of Multiple Sclerosis: Regulation of CXCL12 Expression in Astrocytes by Soluble Myelin Basic Protein. J. Neuroimmunol. 2006, 177, 27–39. [Google Scholar] [CrossRef]
- Clarke, L.E.; Liddelow, S.A.; Chakraborty, C.; Münch, A.E.; Heiman, M.; Barres, B.A. Normal Aging Induces A1-like Astrocyte Reactivity. Proc. Natl. Acad. Sci. USA 2018, 115, E1896–E1905. [Google Scholar] [CrossRef]
- Raj, D.; Yin, Z.; Breur, M.; Doorduin, J.; Holtman, I.R.; Olah, M.; Mantingh-Otter, I.J.; Van Dam, D.; De Deyn, P.P.; den Dunnen, W.; et al. Increased White Matter Inflammation in Aging-and Alzheimer’s Disease Brain. Front. Mol. Neurosci. 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Parravicini, C.; Lecca, D.; Marangon, D.; Coppolino, G.T.; Daniele, S.; Bonfanti, E.; Fumagalli, M.; Raveglia, L.; Martini, C.; Gianazza, E.; et al. Development of the First in Vivo GPR17 Ligand through an Iterative Drug Discovery Pipeline: A Novel Disease-Modifying Strategy for Multiple Sclerosis. PLoS ONE 2020, 15, e0231483. [Google Scholar] [CrossRef]
- Fratangeli, A.; Parmigiani, E.; Fumagalli, M.; Lecca, D.; Benfante, R.; Passafaro, M.; Buffo, A.; Abbracchio, M.P.; Rosa, P. The Regulated Expression, Intracellular Trafficking, and Membrane Recycling of the P2Y-like Receptor GPR17 in Oli-Neu Oligodendroglial Cells. J. Biol. Chem. 2013, 288, 5241–5256. [Google Scholar] [CrossRef]
- Daniele, S.; Trincavelli, M.L.; Fumagalli, M.; Zappelli, E.; Lecca, D.; Bonfanti, E.; Campiglia, P.; Abbracchio, M.P.; Martini, C. Does GRK-β Arrestin Machinery Work as a “Switch on” for GPR17-Mediated Activation of Intracellular Signaling Pathways? Cell. Signal. 2014, 26, 1310–1325. [Google Scholar] [CrossRef]
- Gelosa, P.; Bonfanti, E.; Castiglioni, L.; Delgado-Garcia, J.M.; Gruart, A.; Fontana, L.; Gotti, M.; Tremoli, E.; Lecca, D.; Fumagalli, M.; et al. Improvement of Fiber Connectivity and Functional Recovery after Stroke by Montelukast, an Available and Safe Anti-Asthmatic Drug. Pharmacol. Res. 2019, 142, 223–236. [Google Scholar] [CrossRef]
- Ou, Z.; Sun, Y.; Lin, L.; You, N.; Liu, X.; Li, H.; Ma, Y.; Cao, L.; Han, Y.; Liu, M.; et al. Olig2-Targeted G-Protein-Coupled Receptor Gpr17 Regulates Oligodendrocyte Survival in Response to Lysolecithin-Induced Demyelination. J. Neurosci. 2016, 36, 10560–10573. [Google Scholar] [CrossRef] [PubMed]
- De Groot, C.J.; Bergers, E.; Kamphorst, W.; Ravid, R.; Polman, C.H.; Barkhof, F.; van der Valk, P. Post-Mortem MRI-Guided Sampling of Multiple Sclerosis Brain Lesions: Increased Yield of Active Demyelinating and (p)Reactive Lesions. Brain 2001, 124, 1635–1645. [Google Scholar] [CrossRef] [PubMed]
- Donninelli, G.; Saraf-Sinik, I.; Mazziotti, V.; Capone, A.; Grasso, M.G.; Battistini, L.; Reynolds, R.; Magliozzi, R.; Volpe, E. Interleukin-9 Regulates Macrophage Activation in the Progressive Multiple Sclerosis Brain. J. Neuroinflamm. 2020, 17, 149. [Google Scholar] [CrossRef]
- Magliozzi, R.; Howell, O.; Vora, A.; Serafini, B.; Nicholas, R.; Puopolo, M.; Reynolds, R.; Aloisi, F. Meningeal B-Cell Follicles in Secondary Progressive Multiple Sclerosis Associate with Early Onset of Disease and Severe Cortical Pathology. Brain 2007, 130, 1089–1104. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | # Subjects | Sample | Identified Condition in WM | GPR17 Abundance Score |
| CASES | MS 179 | P4A1 | AL | + |
| NAWM | ++ | |||
| A3A2 | AL | + | ||
| NAWM | ++ | |||
| MS 230 | P5C2 | AL | ++ | |
| NAWM | ++ | |||
| P2D2 | AL | ++ | ||
| CIL | ++ | |||
| AL | ++ | |||
| NAWM | ++ | |||
| P2E1 | CAL | ++ | ||
| AL | ++ | |||
| NAWM | ++ | |||
| MS 234 | A2D4 | CIL | + | |
| CIL | + | |||
| AL | − | |||
| CIL | + | |||
| NAWM | ++ | |||
| MS 242 | P3B3 | AL | ++ | |
| NAWM | ++ | |||
| P4C3 | NAWM | + | ||
| A2B2 | AL | ++ | ||
| CIL | − | |||
| NAWM | ++ | |||
| P5A2 | CIL | − | ||
| AL | + | |||
| NAWM | ++ | |||
| NAWM | ++ | |||
| P1A2 | CAL | − | ||
| AL | − | |||
| NAWM | + | |||
| P3A3 | NAWM | ++ | ||
| MS 297 | A4B3 | AL | + | |
| NAWM | ++ | |||
| P1C2 | NAWM | ++ | ||
| MS 298 | A2B4 | AL | + | |
| NAWM | ++ | |||
| P1B5 | AL | + | ||
| NAWM | ++ | |||
| A4C3 | CAL | ++ | ||
| NAWM | ++ | |||
| MS 300 | A1A3 | AL | ++ | |
| NAWM | ++ | |||
| P1A2 | CAL | − | ||
| AL | − | |||
| NAWM | + | |||
| P3A3 | NAWM | ++ | ||
| (B) | # Subject | Sample | Identified Condition in WM | GPR17 Abundance Score |
| CONTROLS | C 014 | A1D7 | NWM | + |
| C 036 | A1B2 | NWM | + | |
| C 048 | P2F6 | NWM | ++ | |
| C 039 | A2B6 | NWM | + | |
| C 043 | P4D6 | NWM | + |
| Characteristics | Primary Progressive MS | Secondary Progressive MS | Total Secondary MS | Controls |
|---|---|---|---|---|
| Subjects, N | 1 | 8 | 9 | 5 |
| Age, years, mean (SD) | 39 (0) | 55.6 (±11.3) | 53.8 (±11.9) | 73.8 (±10) |
| Gender, women/men | 1/0 | 5/3 | 6/3 | 1/4 |
| Age at onset, years, mean (SD) | 24 (0) | 31.1 (±9) | 31.2 (±8.9) | / |
| Time to wheelchair, median (SD) | 8 (0) | 7.9 (±4.4) | 7.9 (±4.8) | / |
| Disease duration, years, mean (SD) | 15 (0) | 23.5 (±10.2) | 22.5 (±10) | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelini, J.; Marangon, D.; Raffaele, S.; Lecca, D.; Abbracchio, M.P. The Distribution of GPR17-Expressing Cells Correlates with White Matter Inflammation Status in Brain Tissues of Multiple Sclerosis Patients. Int. J. Mol. Sci. 2021, 22, 4574. https://doi.org/10.3390/ijms22094574
Angelini J, Marangon D, Raffaele S, Lecca D, Abbracchio MP. The Distribution of GPR17-Expressing Cells Correlates with White Matter Inflammation Status in Brain Tissues of Multiple Sclerosis Patients. International Journal of Molecular Sciences. 2021; 22(9):4574. https://doi.org/10.3390/ijms22094574
Chicago/Turabian StyleAngelini, Jacopo, Davide Marangon, Stefano Raffaele, Davide Lecca, and Maria P. Abbracchio. 2021. "The Distribution of GPR17-Expressing Cells Correlates with White Matter Inflammation Status in Brain Tissues of Multiple Sclerosis Patients" International Journal of Molecular Sciences 22, no. 9: 4574. https://doi.org/10.3390/ijms22094574
APA StyleAngelini, J., Marangon, D., Raffaele, S., Lecca, D., & Abbracchio, M. P. (2021). The Distribution of GPR17-Expressing Cells Correlates with White Matter Inflammation Status in Brain Tissues of Multiple Sclerosis Patients. International Journal of Molecular Sciences, 22(9), 4574. https://doi.org/10.3390/ijms22094574