
Molecular Mechanism of Thymidylate Synthase Inhibition by N4-Hydroxy-dCMP in View of Spectrophotometric and Crystallographic Studies

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
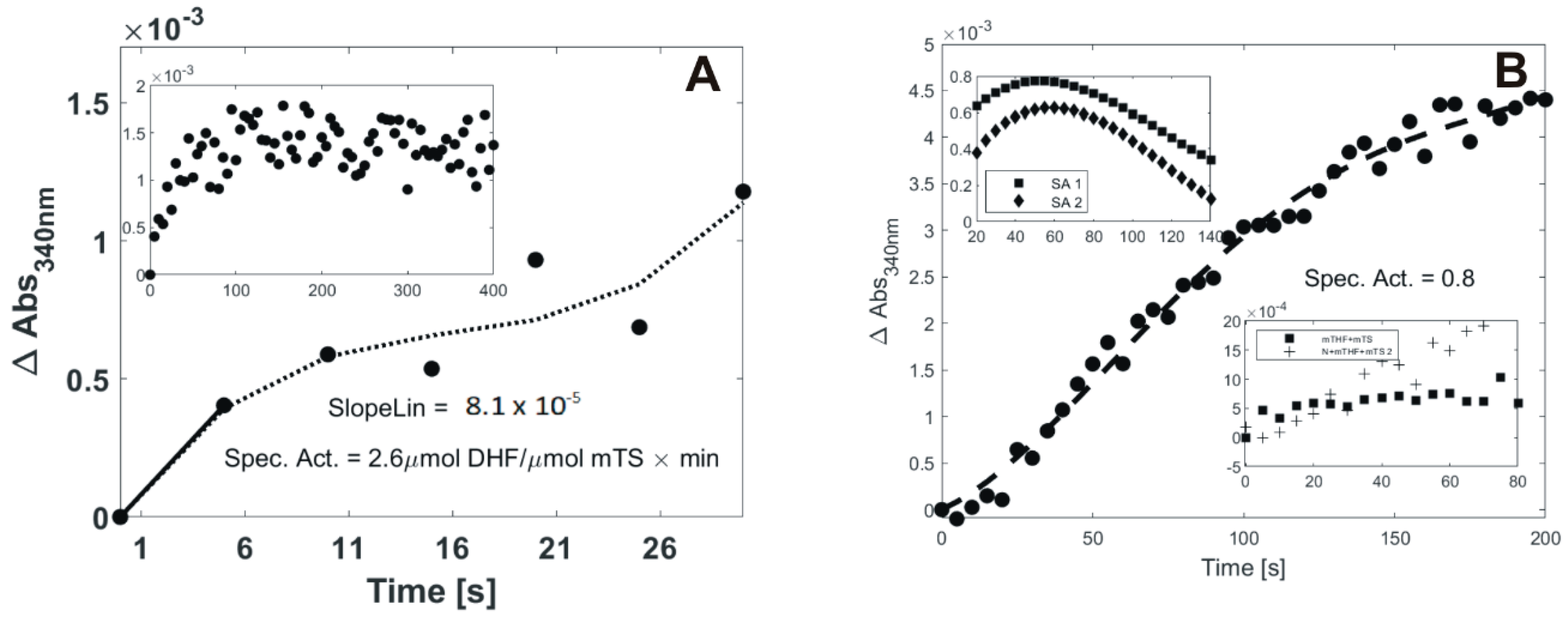
2.1. DHF Production in the Course of N4-OH-dCMP Incubation with TS and meTHF
2.2. Crystallographic Studies
2.3. Crystal Structure of CeTS Soaked with N4-OH-dCMP (PDB ID: 4PSG)
2.4. Crystal Structure of TspTS Cocrystallized with N4-OH-dCMP and meTHF (PDB ID:5M4Z)
2.5. Crystal Structure of mTS Cocrystallized with N4-OH-dCMP and Soaked with meTHF (PDB ID: 6F6Z)
3. Discussion and Conclusions
4. Material and Methods
4.1. Enzyme Preparation
4.2. Spectrophotometric Monitoring of a Time-Dependent DHF Production
4.3. Crystallization and Data Collection
4.4. Structure Determination and Refinement
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| N4-OH-dCMP | N4-hydroxy-dCMP |
| meTHF | N5,10-methylenetetrahydrofolate |
| THF | tetrahydrofolate |
| DHF | dihydrofolate |
| TS | thymidylate synthase |
| m | mouse |
| Tsp | Trichinella spiralis |
| Ce | Caenorhabditis elegans |
References
- Carreras, C.W.; Santi, D.V. The catalytic mechanism and structure of thymidylate synthase. Annu. Rev. Biochem. 1995, 64, 721–762. [Google Scholar] [CrossRef]
- Rode, W.; Leś, A. Molecular mechanism of thymidylate synthase-catalyzed reaction and interaction of the enzyme with 2- and/or 4-substituted analogues of dUMP and 5-fluoro-dUMP. Acta Biochim. Pol. 1996, 43, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Costi, M.P.; Ferrari, S.; Venturelli, A.; Calò, S.; Tondi, D.; Barlocco, D. Thymidylate synthase structure, function and implication in drug discovery. Curr. Med. Chem. 2005, 12, 2241–2258. [Google Scholar] [CrossRef] [PubMed]
- Jarmuła, A. Antifolate inhibitors of thymidylate synthase as anticancer drugs. Mini Rev. Med. Chem. 2010, 10, 1211–1222. [Google Scholar] [CrossRef] [PubMed]
- Taddia, L.; D’Arca, D.; Ferrari, S.; Marraccini, C.; Severi, L.; Ponterini, G.; Assaraf, Y.G.; Marverti, G.; Costi, M.P. Inside the biochemical pathways of thymidylate synthase perturbed by anticancer drugs: Novel strategies to overcome cancer chemoresistance. Drug Resist. Updat. 2015, 23, 20–54. [Google Scholar] [CrossRef]
- Morrison, J.F. The slow-binding and slow, tight-binding inhibition of enzyme-catalysed reactions. Trends Biochem. Sci. 1982, 7, 102–105. [Google Scholar] [CrossRef]
- Lorenson, M.Y.; Maley, G.F.; Maley, F. The purification and properties of thymidylate synthetase from chick embryo extracts. J. Biol. Chem. 1967, 242, 3332–3344. [Google Scholar] [CrossRef]
- Goldstein, S.; Pogolotti, A.L., Jr.; Garvey, E.P.; Santi, D.V. Interaction of N4-hydroxy-2′-deoxycytidilic acid with thymidylate synthetase. J. Med. Chem. 1984, 27, 1259–1262. [Google Scholar] [CrossRef]
- Rode, W.; Zieliński, Z.; Dzik, J.M.; Kulikowski, T.; Bretner, M.; Kierdaszuk, B.; Cieśla, J.; Shugar, D. Mechanism of inhibition of mammalian tumor and other thymidylate synthases by N4-hydroxy-dCMP, N4-hydroxy-5-fluoro-dCMP, and related analogues. Biochemistry 1990, 29, 10835–10842. [Google Scholar] [CrossRef] [PubMed]
- Rode, W.; Dąbrowska, M.; Zieliński, Z.; Gołos, B.; Wranicz, M.; Felczak, K.; Kulikowski, T. Trichinella spiralis and Trichinella pseudospiralis: Developmental patterns of enzymes involved in thymidylate biosynthesis and pyrimidine salvage. Parasitology 2000, 120, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Felczak, K.; Miazga, A.; Poznański, J.; Bretner, M.; Kulikowski, T.; Dzik, J.M.; Gołos, B.; Zieliński, Z.; Cieśla, J.; Rode, W. 5-Substituted N4-hydroxy-2′-deoxycytidines and their 5′-monophosphates: Synthesis, conformation, interaction with tumor thymidylate synthase, and in vitro antitumor activity. J. Med. Chem. 2000, 43, 4647–4656. [Google Scholar] [CrossRef] [PubMed]
- Stroud, R.M.; Finer-Moore, J.S. Conformational Dynamics along an Enzymatic Reaction Pathway: Thymidylate Synthase, “the Movie”. Biochemistry 2003, 42, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Dowierciał, A.; Jarmuła, A.; Wilk, P.; Rypniewski, W.; Kowalska, M.; Frączyk, T.; Cieśla, J.; Rode, W. Mouse thymidylate synthase does not show the inactive conformation, observed for the human enzyme. Struct. Chem. 2017, 28, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Dowierciał, A.; Jarmuła, A.; Wilk, P.; Rypniewski, W.; Kierdaszuk, B.; Rode, W. Crystal structures of complexes of mouse thymidylate synthase crystallized with N4-OH-dCMP alone or in the presence of N5,10-methylenetetrahydrofolate. Pteridines 2013, 24, 93–98. [Google Scholar] [CrossRef]
- Ludwiczak, J.; Maj, P.; Wilk, P.; Frączyk, T.; Ruman, T.; Kierdaszuk, B.; Jarmuła, A.; Rode, W. Phosphorylation of thymidylate synthase affects slow-binding inhibition by 5-fluoro-dUMP and N4-hydroxy-dCMP. Mol. BioSyst. 2016, 12, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Konagurthu, A.S.; Whisslock, J.C.; Stuckey, P.J.; Lesk, A.M. MUSTANG: A multiple structural alignment algorithm. Proteins 2006, 64, 559–574. [Google Scholar] [CrossRef]
- Sotelo-Mundo, R.R.; Cieśla, J.; Dzik, J.M.; Rode, W.; Maley, F.; Maley, G.F.; Hardy, L.W.; Montfort, W.R. Crystal structures of rat thymidylate synthase inhibited by Tomudex, a potent anticancer drug. Biochemistry 1999, 38, 1087–1094. [Google Scholar] [CrossRef]
- Jarmuła, A.; Wilk, P.; Maj, P.; Ludwiczak, J.; Dowierciał, A.; Banaszak, K.; Rypniewski, W.; Cieśla, J.; Dąbrowska, M.; Frączyk, T.; et al. Crystal structures of nematode (parasitic T. spiralis and free living C. elegans), compared to mammalian, thymidylate synthases (TS). Molecular docking and molecular dynamics simulations in search for nematode-specific inhibitors of TS. J. Mol. Graph. Model. 2017, 77, 33–50. [Google Scholar] [CrossRef]
- Dowierciał, A.; Wilk, P.; Rypniewski, W.; Rode, W.; Jarmuła, A. Crystal Structure of Mouse Thymidylate Synthase in Tertiary Complex with dUMP and Raltitrexed Reveals N-Terminus Architecture and Two Different Active Site Conformations. BioMed Res. Int. 2014, 2014, 945803. [Google Scholar] [CrossRef]
- Dowierciał, A.; Jarmuła, A.; Rypniewski, W.; Sokołowska, M.; Frączyk, T.; Cieśla, J.; Rode, W. Crystal structures of substrate- and sulfate-bound mouse thymidylate synthase. Pteridines 2009, 20, 163–167. [Google Scholar]
- Dowierciał, A.; Wilk, P.; Rypniewski, W.; Frączyk, T.; Jarmuła, A.; Banaszak, K.; Dąbrowska, M.; Cieśla, J.; Rode, W. Crystal structures of thymidylate synthase from nematodes, Trichinella spiralis and Caenorhabditis elegans, as a potential template for species-specific drug design. Pteridines 2013, 24, 87–91. [Google Scholar] [CrossRef]
- Wilk, P.; Jarmuła, A.; Ruman, T.; Banaszak, K.; Rypniewski, W.; Cieśla, J.; Dowierciał, A.; Rode, W. Crystal structure of phosphoramide-phosphorylated thymidylate synthase reveals pSer127, reflecting probably pHis to pSer phosphotransfer. Bioorg. Chem. 2014, 52, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Sobich, J.; Prokopowicz, M.; Maj, P.; Wilk, P.; Zieliński, Z.; Frączyk, T.; Rode, W. Thymidylate synthase-catalyzed, tetrahydrofolate-dependent self-inactivation by 5-FdUMP. Arch. Biochem. Biophys. 2019, 674, 108106. [Google Scholar] [CrossRef]
- Garg, D.; Skouloubris, S.; Briffotaux, J.; Myllykallio, H.; Wade, R.C. Conservation and role of electrostatics in thymidylate synthase. Sci. Rep. 2015, 5, 17356. [Google Scholar] [CrossRef] [Green Version]
- Burns, A.R.; Luciani, G.M.; Musso, G.; Bagg, R.; Yeo, M.; Zhang, Y.; Rajendran, L.; Glavin, J.; Hunter, R.; Redman, E.; et al. Caenorhabditis elegans is a useful model for anthelmintic discovery. Nat. Commun. 2015, 6, 7485. [Google Scholar] [CrossRef]
- Jägerstad, M.; Jastrebova, J. 5,10-Methylene-tetrahydrofolate dissociates into tetrahydrofolate and formaldehyde at physiological pH and acidic pH, typical conditions used during sample extraction and LC-MS/MS analysis of biological samples. Biomed. Chromatogr. 2014, 28, 1041–1042. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.; Maley, F.; Kohen, A. Role of Y94 in proton and hydride transfers catalyzed by thymidylate synthase. Biochemistry 2007, 46, 14188–14197. [Google Scholar] [CrossRef] [Green Version]
- Moffat, K.; Henderson, R. Freeze trapping of reaction intermediates. Curr. Opin. Struct. Biol. 1995, 5, 656–663. [Google Scholar] [CrossRef]
- Kumar, M.; Prashar, V.; Mahale, S.; Hosur, M.V. Observation of a tetrahedral reaction intermediate in the HIV-1 protease-substrate complex. Biochem. J. 2005, 389, 365–371. [Google Scholar] [CrossRef]
- Bourgeois, D.; Weik, M. Kinetic protein crystallography: A tool to watch proteins in action. Crystallogr. Rev. 2009, 15, 87–118. [Google Scholar] [CrossRef]
- Nakamura, T.; Zhao, Y.; Yamagata, Y.; Hua, Y.J.; Yang, W. Watching DNA polymerase η make a phosphodiester bond. Nature 2012, 487, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Volbeda, A.; Saez Cabodevilla, J.; Darnault, C.; Gigarel, O.; Han, T.H.; Renoux, O.; Hamelin, O.; Ollagnierde-Choudens, S.; Amara, P.; Fontecilla-Camps, J.C. Crystallographic trapping of reaction intermediates in quinolinic acid synthesis by NadA. ACS Chem. Biol. 2018, 13, 1209–1217. [Google Scholar] [CrossRef] [PubMed]
- Albesa-Jové, D.; Cifuente, J.O.; Trastoy, B.; Guerin, M.E. Quick-soaking of crystals reveals unprecedented insights into the catalytic mechanism of glycosyltransferases. Methods Enzymol. 2019, 621, 261–279. [Google Scholar] [PubMed]
- Finer-Moore, J.S.; Liu, L.; Birdsall, D.L.; Brem, R.; Apfeld, J.; Santi, D.V.; Stroud, R.M. Contributions of orientation and hydrogen bonding to catalysis in Asn229 mutants of thymidylate synthase. J. Mol. Biol. 1998, 276, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Cieśla, J.; Gołos, B.; Wałajtys-Rode, E.; Jagielska, E.; Płucienniczak, A.; Rode, W. The effect of Arg 209 to Lys mutation in mouse thymidylate synthase. Acta Biochim. Pol. 2002, 49, 651–658. [Google Scholar] [CrossRef] [Green Version]
- Wińska, P.; Gołos, B.; Cieśla, J.; Zieliński, Z.; Frączyk, T.; Wałajtys-Rode, E.; Rode, W. Developmental arrest in Caenorhabditis elegans dauer larvae causes high expression of enzymes involved in thymidylate biosynthesis, similar to that found in Trichinella muscle larvae. Parasitology 2005, 131, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Frączyk, T.; Kubiński, K.; Masłyk, M.; Cieśla, J.; Hellman, U.; Shugar, D.; Rode, W. Phosphorylation of thymidylate synthase from various sources by human protein kinase CK2 and its catalytic subunits. Bioorg. Chem. 2010, 38, 124–131. [Google Scholar] [CrossRef]
- Frączyk, T.; Ruman, T.; Wilk, P.; Palmowski, P.; Rogowska-Wrzesinska, A.; Cieśla, J.; Zieliński, Z.; Nizioł, J.; Jarmuła, A.; Maj, P.; et al. Properties of phosphorylated thymidylate synthase. Biochim. Biophys. Acta 2015, 1854, 1922–1934. [Google Scholar] [CrossRef]
- Wahba, A.J.; Friedkin, M. The enzymatic synthesis of thymidylate. I. Early steps in the purification of thymidylate synthetase of Escherichia coli. J. Biol. Chem. 1962, 237, 3794–3801. [Google Scholar] [CrossRef]
- Rode, W.; Kulikowski, T.; Kędzierska, B.; Shugar, D. Studies on the interaction with thymidylate synthase of analogues of 2′-deoxyuridine-5′-phosphate and 5-fluoro-2′-deoxyuridine-5′-phosphate with modified phosphate groups. Biochem. Pharmacol. 1987, 36, 203–210. [Google Scholar] [CrossRef]
- Wolschin, F.; Wienkoop, S.; Weckwerth, W. Enrichment of phosphorylated proteins and peptides from complex mixtures using metal oxide/hydroxide affinity chromatography (MOAC). Proteomics 2005, 5, 4389–4397. [Google Scholar] [CrossRef]
- Stoscheck, C.M. Quantitation of protein. Methods Enzymol. 1990, 182, 50–68. [Google Scholar] [CrossRef] [PubMed]
- Wahba, A.J.; Friedkin, M. Direct spectrophotometric evidence for the oxidation of tetrahydrofolate during the enzymatic synthesis of thymidylate. J. Biol. Chem. 1961, 236, PC11–PC12. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. DENZO and SCALEPACK. In International Tables for Crystallography Volume F: Crystallography of Biological Macromolecules; Rossmann, M.G., Arnold, E., Eds.; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar] [CrossRef]
- Evans, P. Scaling and assessment of data quality. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Vagin, A.A.; Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr. 1997, 53, 240–255. [Google Scholar] [CrossRef]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK—A program to check the stereochemical quality of protein structures. J. Appl. Cryst. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urzhumtseva, L.; Afonine, P.V.; Adams, P.D.; Urzhumtsev, A. Crystallographic model quality at a glance. Acta Crystallogr. D Biol. Crystallogr. 2009, 65, 297–300. [Google Scholar] [CrossRef]
- Vaguine, A.A.; Richelle, J.; Wodak, S.J. SFCHECK: A unified set of procedures for evaluating the quality of macromolecular structure-factor data and their agreement with the atomic model. Acta Crystallogr. D Biol. Crystallogr. 1999, 55, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Afonine, P.V.; Moriarty, N.W.; Mustyakimov, M.; Sobolev, O.V.; Terwilliger, T.C.; Turk, D.; Urzhumtsev, A.; Adams, P.D. FEM: Feature-enhanced map. Acta Crystallogr. D Biol. Crystallogr. 2015, 71, 646–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Liu, Q.; Qu, G.; Feng, Y.; Reetz, M.T. Utility of B-factors in protein science: Interpreting rigidity, flexibility, and internal motion and engineering thermostability. Chem. Rev. 2019, 119, 1626–1665. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distance [Å] between: | 4EZ8 | 5M4Z A | 5M4Z B | 6F6Z A | 6F6Z B | 4EIN A | 4EIN B | 4PSG A * | 4PSG B * |
|---|---|---|---|---|---|---|---|---|---|
| Tyr129 hydroxyl and N(4)-OH oxygen atom | 4.57 | 3.67 | 3.47 | 4.09 | 3.98 | 4.38 | 4.33 | 3.76 | 4.46 |
| Centroid of imidazole ring of His190 and oxygen atom of N(4)-OH | 4.54 | 4.20 | 4.13 | 4.17 | 4.20 | 4.35 | 4.29 | 4.57 | 5.03 |
| Centroid of imidazole ring of His190 and centroid of phenyl ring of Tyr129 | 5.81 | 3.95 | 3.90 | 5.53 | 5.60 | 6.08 | 6.08 | 4.15 | 4.25 |
| System | PDB ID | TS Concentration in the Protein-Ligand Solution [mg/mL] | Ligand Concentration in the Protein-Ligand Solution [mM] | Ligand Concentration Used for Crystal Soaking [mM] | Composition of the Well Solution c |
|---|---|---|---|---|---|
| TspTS-N4-OH-dCMP cocrystallized with meTHF | 5M4Z | 20 | N4-OH-dCMP: 7 meTHF: 10 | --- | 0.1M NaF pH 7.1 19% (w/v) PEG 3350 |
| mTS-N4-OH-dCMP a soaked with meTHF | 6F6Z | 20 | N4-OH-dCMP: 6 | meTHF: ≈50 | 0.1M MES pH 6.7 0.15M Mg(CH3COO)2 14% (w/v) PEG 8000 |
| CeTS b soaked with N4-OH-dCMP | 4PSG | 24 | ---- | N4-OH-dCMP: ≈36 | 0.1M MES pH 5.6 0.2M Mg(CH3COO)2 15% (w/v) PEG 8000 |
| CeTS Soaked with N4-OH-dCMP | TspTS Cocrystallized with N4-OH-dCMP and meTHF | mTS Cocrystallized with N4-OH-dCMP and Soaked with meTHF | |
|---|---|---|---|
| PDB ID | 4PSG | 5M4Z | 6F6Z |
| Crystal properties and data collection | |||
| Matthews coefficient [Å3/Da] (solvent content [%]) | 3.17 (61.2) | 2.07 (40.5) | 3.37 (63.5) |
| Space group | P 1 21 1 | P 1 | C 1 2 1 |
| Cell dimensions | a = 59.4 Å, b = 95.9 Å, c = 82.2 Å β = 103.8° α = γ = 90° | a = 51.3 Å, b = 52.8 Å, c = 63.9 Å α = 72.5°, β = 68.0°, γ = 61.2° | a = 160.4 Å, b = 89.2 Å, c = 66.6 Å β = 97.5° α = γ = 90° |
| Wavelength [Å] | 1.54 | 0.918 | 0.918 |
| Resolution [Å] a | 2.80–79.80 (2.80–2.95) | 1.18–45.72 (1.18–1.21) | 2.13–48.81 (2.13–2.26) |
| Unique reflections | 22154 | 172936 | 51781 |
| Completeness [%] a | 99.8 (99.9) | 97.4 (89.0) | 98.2 (98.35) |
| Redundancy | 8.7 | 3.61 | 3.42 |
| I/σ(I) a | 8.1 (5.53) | 16.43 (1.76) | 14.6 (1.11) |
| Refinement | |||
| Resolution [Å] | 2.80–25.11 | 1.18–28.09 | 2.13–47.82 |
| Number of reflections | 22128 | 172936 | 50161 |
| Rwork [%] a | 23.1 (33.5) | 13.8 (28.1) | 21.0 (37.2) |
| Rfree [%] a | 26.7 (41.4) | 15.1 (28.4) | 24.3 (40.3) |
| Average B, all atoms [Å2] | 16.6 b | 23.7 | 49.8 |
| RMS deviations from ideal values for refined atoms | |||
| RMSbond [Å] | 0.011 | 0.008 | 0.006 |
| RMSangle [°] | 1.607 | 1.086 | 0.848 |
| Non-hydrogen atoms used in refinement | |||
| Protein atoms | 4632 | 4676 | 4638 |
| Solvent atoms | 62 | 291 | 250 |
| Heterogen atoms | 42 | 125 | 108 |
| Ramachandran’s plot assignments | |||
| Favored | 556 (96%) | 569 (98%) | 553 (97%) |
| Allowed | 19 (3%) | 9 (2%) | 17 (3%) |
| Disallowed | 2 (0%) | 2 (0%) | 2 (0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maj, P.; Jarmuła, A.; Wilk, P.; Prokopowicz, M.; Rypniewski, W.; Zieliński, Z.; Dowierciał, A.; Bzowska, A.; Rode, W. Molecular Mechanism of Thymidylate Synthase Inhibition by N4-Hydroxy-dCMP in View of Spectrophotometric and Crystallographic Studies. Int. J. Mol. Sci. 2021, 22, 4758. https://doi.org/10.3390/ijms22094758
Maj P, Jarmuła A, Wilk P, Prokopowicz M, Rypniewski W, Zieliński Z, Dowierciał A, Bzowska A, Rode W. Molecular Mechanism of Thymidylate Synthase Inhibition by N4-Hydroxy-dCMP in View of Spectrophotometric and Crystallographic Studies. International Journal of Molecular Sciences. 2021; 22(9):4758. https://doi.org/10.3390/ijms22094758
Chicago/Turabian StyleMaj, Piotr, Adam Jarmuła, Piotr Wilk, Małgorzata Prokopowicz, Wojciech Rypniewski, Zbigniew Zieliński, Anna Dowierciał, Agnieszka Bzowska, and Wojciech Rode. 2021. "Molecular Mechanism of Thymidylate Synthase Inhibition by N4-Hydroxy-dCMP in View of Spectrophotometric and Crystallographic Studies" International Journal of Molecular Sciences 22, no. 9: 4758. https://doi.org/10.3390/ijms22094758