
Effects of Maternal Resveratrol Intake on the Metabolic Health of the Offspring


Abstract
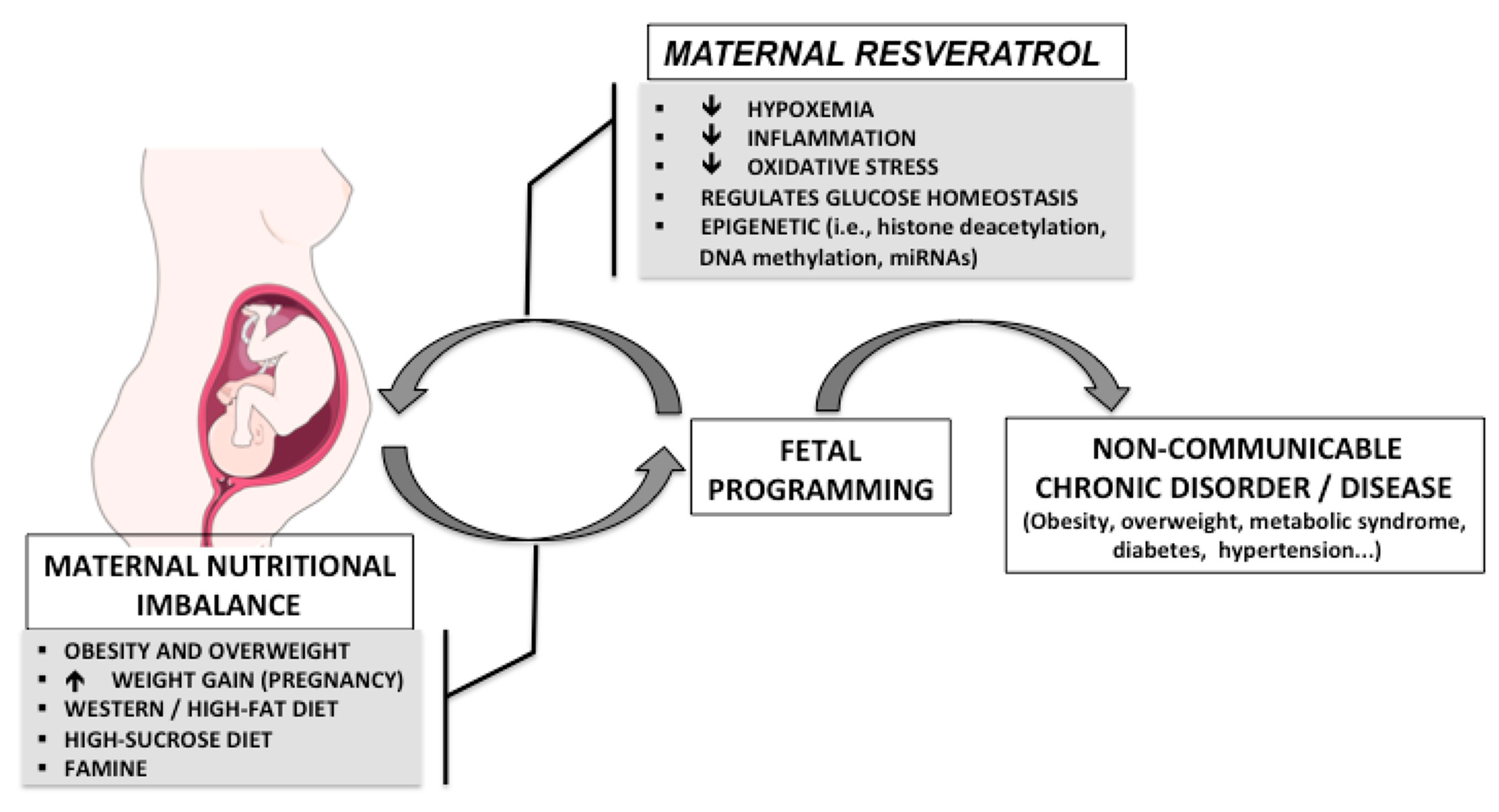
:1. Introduction
2. Metabolic Effects of Maternal Nutritional Imbalances
2.1. Experimental Animal Studies
2.2. Studies in Humans
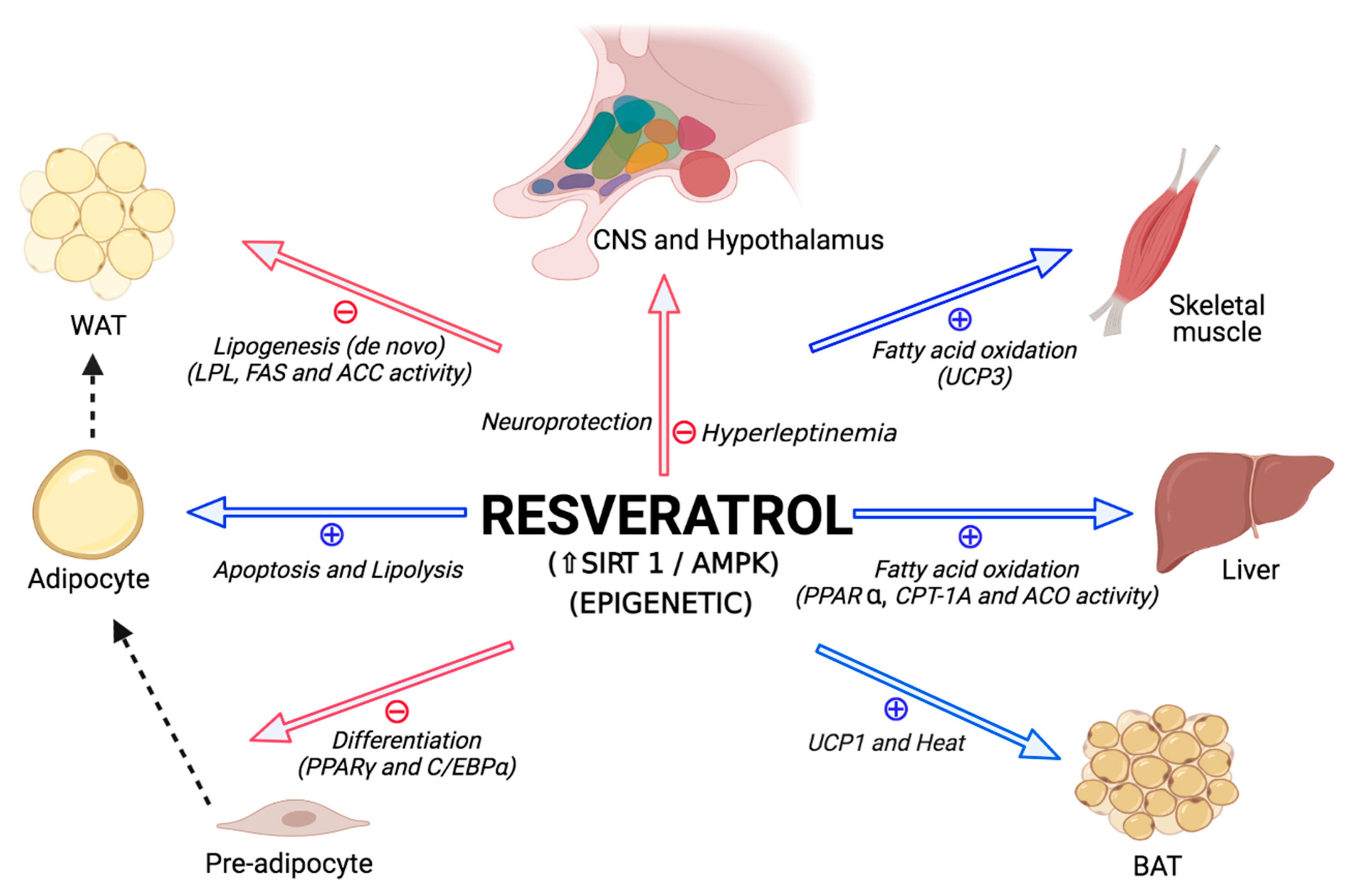
3. Effects of Resveratrol
3.1. Animal Studies
3.2. Human Studies
4. Sex Differences in Response to Perinatal Changes
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barker, D.J.P. Obesity and early life. Obes. Rev. 2007, 8, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Mcmillen, I.C.; Robinson, J.S. Developmental Origins of the Metabolic Syndrome: Prediction, Plasticity, and Programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef] [PubMed]
- Gillman, M.W. Developmental Origins of Health and Disease. N. Engl. J. Med. 2005, 353, 1848–1850. [Google Scholar] [CrossRef] [Green Version]
- Breton, C. The hypothalamus–adipose axis is a key target of developmental programming by maternal nutritional manipulation. J. Endocrinol. 2013, 216, R19–R31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haugen, A.C.; Schug, T.T.; Collman, G.; Heindel, J.J. Evolution of DOHaD: The impact of environmental health sciences. J. Dev. Orig. Health Dis. 2015, 6, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Finer, S.; Iqbal, M.S.; Lowe, R.; Ogunkolade, B.W.; Pervin, S.; Mathews, C.; Smart, M.; Alam, D.S.; Hitman, G.A. Is famine exposure during developmental life in rural Bangladesh associated with a metabolic and epigenetic signature in young adulthood? A historical cohort study. BMJ Open 2016, 6, e011768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksson, J.G. Developmental Origins of Health and Disease—from a small body size at birth to epigenetics. Ann. Med. 2016, 48, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Roseboom, T.J. Epidemiological evidence for the developmental origins of health and disease: Effects of prenatal undernutrition in humans. J. Endocrinol. 2019, 242, T135–T144. [Google Scholar] [CrossRef]
- Koletzko, B.; Brands, B.; Poston, L.; Godfrey, K.; Demmelmair, H. Early Nutrition Project Early nutrition programming of long-term health. Proc. Nutr. Soc. 2012, 71, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Bayol, S.A.; Simbi, B.H.; Bertrand, J.A.; Stickland, N.C. Offspring from mothers fed a ‘junk food’ diet in pregnancy and lactation exhibit exacerbated adiposity that is more pronounced in females: Maternal junk food diet and obesity in offspring. J. Physiol. 2008, 586, 3219–3230. [Google Scholar] [CrossRef]
- Şanlı, E.; Kabaran, S. Maternal Obesity, Maternal Overnutrition and Fetal Programming: Effects of Epigenetic Mechanisms on the Development of Metabolic Disorders. Curr. Genomics 2019, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-C.; Wu, C.-W.; Hung, P.-L.; Chan, J.Y.H.; Tain, Y.-L.; Fu, M.-H.; Chen, L.-W.; Liang, C.-K.; Hung, C.-Y.; Yu, H.-R.; et al. Environmental Stimulation Counteracts the Suppressive Effects of Maternal High-Fructose Diet on Cell Proliferation and Neuronal Differentiation in the Dentate Gyrus of Adult Female Offspring via Histone Deacetylase 4. Int. J. Environ. Res. Public Health 2020, 17, 3919. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.-A.; Tsai, C.-K.; Huang, L.-T.; Sheen, J.-M.; Tiao, M.-M.; Tain, Y.-L.; Chen, C.-C.; Lin, I.-C.; Lai, Y.-J.; Tsai, C.-C.; et al. Maternal Resveratrol Treatment Re-Programs and Maternal High-Fat Diet-Induced Retroperitoneal Adiposity in Male Offspring. Int. J. Environ. Res. Public Health 2020, 17, 2780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, J.; Hovorka, R. Closed-loop control in insulin pumps for type-1 diabetes mellitus: Safety and efficacy. Expert Rev. Med. Devices 2020, 17, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Hardy, D.B. The Fetal Origins of the Metabolic Syndrome: Can We Intervene? J. Pregnancy 2012, 2012, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-Inflammatory Action and Mechanisms of Resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef] [PubMed]
- Truong, V.-L.; Jun, M.; Jeong, W.-S. Role of resveratrol in regulation of cellular defense systems against oxidative stress: Cellular defense systems against oxidative stress. BioFactors 2018, 44, 36–49. [Google Scholar] [CrossRef]
- Franco, J.; Lisboa, P.; da Silva Lima, N.; Peixoto-Silva, N.; Maia, L.; Oliveira, E.; Passos, M.; de Moura, E. Resveratrol Prevents Hyperleptinemia and Central Leptin Resistance in Adult Rats Programmed by Early Weaning. Horm. Metab. Res. 2014, 46, 728–735. [Google Scholar] [CrossRef]
- Martel, J.; Ojcius, D.M.; Chang, C.-J.; Lin, C.-S.; Lu, C.-C.; Ko, Y.-F.; Tseng, S.-F.; Lai, H.-C.; Young, J.D. Anti-obesogenic and antidiabetic effects of plants and mushrooms. Nat. Rev. Endocrinol. 2017, 13, 149–160. [Google Scholar] [CrossRef]
- Aguirre, L.; Fernández-Quintela, A.; Arias, N.; Portillo, M. Resveratrol: Anti-Obesity Mechanisms of Action. Molecules 2014, 19, 18632–18655. [Google Scholar] [CrossRef] [Green Version]
- Fan, E.; Zhang, L.; Jiang, S.; Bai, Y. Beneficial Effects of Resveratrol on Atherosclerosis. J. Med. Food 2008, 11, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wu, C.; Qiu, S.; Yuan, X.; Li, L. Effects of resveratrol on glucose control and insulin sensitivity in subjects with type 2 diabetes: Systematic review and meta-analysis. Nutr. Metab. 2017, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Feng, Q.; Cheng, J.; Zheng, J. Maternal resveratrol consumption and its programming effects on metabolic health in offspring mechanisms and potential implications. Biosci. Rep. 2018, 38, BSR20171741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Zhou, J.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Health Benefits and Molecular Mechanisms of Resveratrol: A Narrative Review. Foods 2020, 9, 340. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.-L.; Hsu, C.-N. Developmental Programming of the Metabolic Syndrome: Can We Reprogram with Resveratrol? Int. J. Mol. Sci. 2018, 19, 2584. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.-H.; Chen, Y.-C.; Sheen, J.-M.; Huang, L.-T. Maternal Obesity Programs Offspring Development and Resveratrol Potentially Reprograms the Effects of Maternal Obesity. Int. J. Environ. Res. Public. Health 2020, 17, 1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darby, J.R.T.; Mohd Dollah, M.H.B.; Regnault, T.R.H.; Williams, M.T.; Morrison, J.L. Systematic review: Impact of resveratrol exposure during pregnancy on maternal and fetal outcomes in animal models of human pregnancy complications—Are we ready for the clinic? Pharmacol. Res. 2019, 144, 264–278. [Google Scholar] [CrossRef]
- Argente-Arizón, P.; Ros, P.; Díaz, F.; Fuente-Martin, E.; Castro-González, D.; Sánchez-Garrido, M.Á.; Barrios, V.; Tena-Sempere, M.; Argente, J.; Chowen, J.A. Age and sex dependent effects of early overnutrition on metabolic parameters and the role of neonatal androgens. Biol. Sex Differ. 2016, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Vega, C.C.; Reyes-Castro, L.A.; Rodríguez-González, G.L.; Bautista, C.J.; Vázquez-Martínez, M.; Larrea, F.; Chamorro-Cevallos, G.A.; Nathanielsz, P.W.; Zambrano, E. Resveratrol partially prevents oxidative stress and metabolic dysfunction in pregnant rats fed a low protein diet and their offspring: Resveratrol intervention prevents programming. J. Physiol. 2016, 594, 1483–1499. [Google Scholar] [CrossRef] [Green Version]
- Ros, P.; Díaz, F.; Freire-Regatillo, A.; Argente-Arizón, P.; Barrios, V.; Argente, J.; Chowen, J.A. Resveratrol Intake During Pregnancy and Lactation Modulates the Early Metabolic Effects of Maternal Nutrition Differently in Male and Female Offspring. Endocrinology 2018, 159, 810–825. [Google Scholar] [CrossRef]
- Ros, P.; Díaz, F.; Freire-Regatillo, A.; Argente-Arizón, P.; Barrios, V.; Argente, J.; Chowen, J.A. Sex Differences in Long-term Metabolic Effects of Maternal Resveratrol Intake in Adult Rat Offspring. Endocrinology 2020, 161, bqaa090. [Google Scholar] [CrossRef] [PubMed]
- Chowen, J.A.; Freire-Regatillo, A.; Argente, J. Neurobiological characteristics underlying metabolic differences between males and females. Prog. Neurobiol. 2019, 176, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Macotela, Y.; Boucher, J.; Tran, T.T.; Kahn, C.R. Sex and Depot Differences in Adipocyte Insulin Sensitivity and Glucose Metabolism. Diabetes 2009, 58, 803–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.C.K. Life history trade-offs and the partitioning of maternal investment. Evol. Med. Public Health 2018, 2018, 153–166. [Google Scholar] [CrossRef]
- Zhou, L.-Y.; Deng, M.-Q.; Zhang, Q.; Xiao, X.-H. Early-life nutrition and metabolic disorders in later life: A new perspective on energy metabolism. Chin. Med. J. 2020, 133, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Godfrey, K.M.; Poston, L.; Szajewska, H.; van Goudoever, J.B.; de Waard, M.; Brands, B.; Grivell, R.M.; Deussen, A.R.; Dodd, J.M.; et al. Nutrition During Pregnancy, Lactation and Early Childhood and its Implications for Maternal and Long-Term Child Health: The Early Nutrition Project Recommendations. Ann. Nutr. Metab. 2019, 74, 93–106. [Google Scholar] [CrossRef]
- Huang, L.-T. Maternal and Early-Life Nutrition and Health. Int. J. Environ. Res. Public Health 2020, 17, 7982. [Google Scholar] [CrossRef]
- Koletzko, B.; Brands, B.; Chourdakis, M.; Cramer, S.; Grote, V.; Hellmuth, C.; Kirchberg, F.; Prell, C.; Rzehak, P.; Uhl, O.; et al. The Power of Programming and the EarlyNutrition project: Opportunities for health promotion by nutrition during the first thousand days of life and beyond. Ann. Nutr. Metab. 2014, 64, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Cáceres, C.; Fuente-Martín, E.; Burgos-Ramos, E.; Granado, M.; Frago, L.M.; Barrios, V.; Horvath, T.; Argente, J.; Chowen, J.A. Differential Acute and Chronic Effects of Leptin on Hypothalamic Astrocyte Morphology and Synaptic Protein Levels. Endocrinology 2011, 152, 1809–1818. [Google Scholar] [CrossRef]
- Chang, G.-Q.; Gaysinskaya, V.; Karatayev, O.; Leibowitz, S.F. Maternal High-Fat Diet and Fetal Programming: Increased Proliferation of Hypothalamic Peptide-Producing Neurons That Increase Risk for Overeating and Obesity. J. Neurosci. 2008, 28, 12107–12119. [Google Scholar] [CrossRef] [Green Version]
- Grayson, B.E.; Allen, S.E.; Billes, S.K.; Williams, S.M.; Smith, M.S.; Grove, K.L. Prenatal development of hypothalamic neuropeptide systems in the nonhuman primate. Neuroscience 2006, 143, 975–986. [Google Scholar] [CrossRef]
- Ingvorsen, C.; Lelliott, C.J.; Brix, S.; Hellgren, L.I. Effects of maternal high-fat/high sucrose diet on hepatic lipid metabolism in rat offspring. Clin. Exp. Pharmacol. Physiol. 2021, 48, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Tobar-Bernal, F.A.; Zamudio, S.R.; Quevedo-Corona, L. The high-fructose intake of dams during pregnancy and lactation exerts sex-specific effects on adult rat offspring metabolism. J. Dev. Orig. Health Dis. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-C.; Wu, C.-W.; Fu, M.-H.; Tain, Y.-L.; Liang, C.-K.; Hung, C.-Y.; Chen, I.-C.; Lee, Y.-C.; Wu, C.-Y.; Wu, K.L.H. Maternal high fructose-induced hippocampal neuroinflammation in the adult female offspring via PPARγ-NF-κB signaling. J. Nutr. Biochem. 2020, 81, 108378. [Google Scholar] [CrossRef]
- Isganaitis, E.; Jimenez-Chillaron, J.; Woo, M.; Chow, A.; DeCoste, J.; Vokes, M.; Liu, M.; Kasif, S.; Zavacki, A.-M.; Leshan, R.L.; et al. Accelerated Postnatal Growth Increases Lipogenic Gene Expression and Adipocyte Size in Low-Birth Weight Mice. Diabetes 2009, 58, 1192–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Xiao, X.; Zhang, Q.; Wang, T.; Yu, M.; Xu, J. Maternal Low-Protein Diet Modulates Glucose Metabolism and Hepatic MicroRNAs Expression in the Early Life of Offspring. Nutrients 2017, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Wang, X.; Li, Q.; Han, B.; Chen, Y.; Zhu, C.; Chen, Y.; Lin, D.; Wang, B.; Jensen, M.D.; et al. The famine exposure in early life and metabolic syndrome in adulthood. Clin. Nutr. 2017, 36, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhao, L.; Ning, Z.; Li, Q.; Han, B.; Cheng, J.; Chen, Y.; Nie, X.; Xia, F.; Wang, N.; et al. Famine exposure in early life is associated with visceral adipose dysfunction in adult females. Eur. J. Nutr. 2019, 58, 1625–1633. [Google Scholar] [CrossRef]
- Clancy, B.; Finlay, B.L.; Darlington, R.B.; Anand, K.J.S. Extrapolating brain development from experimental species to humans. NeuroToxicology 2007, 28, 931–937. [Google Scholar] [CrossRef] [Green Version]
- Ravelli, A.; van der Meulen, J.; Michels, R.; Osmond, C.; Barker, D.; Hales, C.; Bleker, O. Glucose tolerance in adults after prenatal exposure to famine. Lancet 1998, 351, 173–177. [Google Scholar] [CrossRef]
- Lumey, L.; Stein, A.D.; Kahn, H.S.; Romijn, J. Lipid profiles in middle-aged men and women after famine exposure during gestation: The Dutch Hunger Winter Families Study. Am. J. Clin. Nutr. 2009, 89, 1737–1743. [Google Scholar] [CrossRef] [Green Version]
- Leonard, S.A.; Rasmussen, K.M.; King, J.C.; Abrams, B. Trajectories of maternal weight from before pregnancy through postpartum and associations with childhood obesity. Am. J. Clin. Nutr. 2017, ajcn158683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, K.M.; Reynolds, R.M.; Prescott, S.L.; Nyirenda, M.; Jaddoe, V.W.V.; Eriksson, J.G.; Broekman, B.F.P. Influence of maternal obesity on the long-term health of offspring. Lancet Diabetes Endocrinol. 2017, 5, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Tam, C.H.T.; Ma, R.C.W.; Yuen, L.Y.; Ozaki, R.; Li, A.M.; Hou, Y.; Chan, M.H.M.; Ho, C.S.; Yang, X.; Chan, J.C.N.; et al. The impact of maternal gestational weight gain on cardiometabolic risk factors in children. Diabetologia 2018, 61, 2539–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heslehurst, N.; Vieira, R.; Akhter, Z.; Bailey, H.; Slack, E.; Ngongalah, L.; Pemu, A.; Rankin, J. The association between maternal body mass index and child obesity: A systematic review and meta-analysis. PLoS Med. 2019, 16, e1002817. [Google Scholar] [CrossRef]
- Hou, C.-Y.; Tain, Y.-L.; Yu, H.-R.; Huang, L.-T. The Effects of Resveratrol in the Treatment of Metabolic Syndrome. Int. J. Mol. Sci. 2019, 20, 535. [Google Scholar] [CrossRef] [Green Version]
- Boccellino, M.; D’Angelo, S. Anti-Obesity Effects of Polyphenol Intake: Current Status and Future Possibilities. Int. J. Mol. Sci. 2020, 21, 5642. [Google Scholar] [CrossRef]
- Franco, J.G.; Dias-Rocha, C.P.; Fernandes, T.P.; Albuquerque Maia, L.; Lisboa, P.C.; Moura, E.G.; Pazos-Moura, C.C.; Trevenzoli, I.H. Resveratrol treatment rescues hyperleptinemia and improves hypothalamic leptin signaling programmed by maternal high-fat diet in rats. Eur. J. Nutr. 2016, 55, 601–610. [Google Scholar] [CrossRef]
- Silva, L.B.A.R.; Pinheiro-Castro, N.; Novaes, G.M.; Pascoal, G.L.; Ong, T.P. Bioactive food compounds, epigenetics and chronic disease prevention: Focus on early-life interventions with polyphenols. Food Res. Int. 2019, 125, 108646. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.S.; Cantó, C. The molecular targets of resveratrol. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2015, 1852, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Singh, C.K.; Kumar, A.; Hitchcock, D.B.; Fan, D.; Goodwin, R.; LaVoie, H.A.; Nagarkatti, P.; DiPette, D.J.; Singh, U.S. Resveratrol prevents embryonic oxidative stress and apoptosis associated with diabetic embryopathy and improves glucose and lipid profile of diabetic dam. Mol. Nutr. Food Res. 2011, 55, 1186–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, V.H.J.; Pound, L.D.; Thorn, S.R.; Gillingham, M.B.; Thornburg, K.L.; Friedman, J.E.; Frias, A.E.; Grove, K.L. Beneficial and cautionary outcomes of resveratrol supplementation in pregnant nonhuman primates. FASEB J. 2014, 28, 2466–2477. [Google Scholar] [CrossRef] [Green Version]
- Pound, L.D.; Comstock, S.M.; Grove, K.L. Consumption of a Western-style diet during pregnancy impairs offspring islet vascularization in a Japanese macaque model. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E115–E123. [Google Scholar] [CrossRef] [PubMed]
- O’Tierney-Ginn, P.; Roberts, V.; Gillingham, M.; Walker, J.; Glazebrook, P.A.; Thornburg, K.L.; Grove, K.; Frias, A.E. Influence of high fat diet and resveratrol supplementation on placental fatty acid uptake in the Japanese macaque. Placenta 2015, 36, 903–910. [Google Scholar] [CrossRef] [Green Version]
- Trejo-González, N.L.; Chirino-Galindo, G.; Palomar-Morales, M. Antiteratogenic capacity of resveratrol in streptozotocin-induced diabetes in rats. Rev. Peru. Med. Exp. Salud Publica 2015, 32, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.; Chen, D.; Yang, Q.; Wang, B.; Zhu, M.-J.; Nathanielsz, P.W.; Du, M. Resveratrol supplementation of high-fat diet-fed pregnant mice promotes brown and beige adipocyte development and prevents obesity in male offspring: Maternal resveratrol promotes beige adipogenesis in offspring. J. Physiol. 2017, 595, 1547–1562. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Tomihara, T.; Kimura, G.; Ueno, Y.; Ketema, R.M.; Sato, S.; Mukai, Y.; Sikder, T.; Kurasaki, M.; Hosokawa, T.; et al. Long-term effects of maternal resveratrol intake during lactation on cholesterol metabolism in male rat offspring. Int. J. Food Sci. Nutr. 2020, 71, 226–234. [Google Scholar] [CrossRef]
- Yao, L.; Wan, J.; Li, H.; Ding, J.; Wang, Y.; Wang, X.; Li, M. Resveratrol relieves gestational diabetes mellitus in mice through activating AMPK. Reprod. Biol. Endocrinol. 2015, 13, 118. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.-Y.; Yu, H.-R.; Tsai, C.-C.; Huang, L.-T.; Chen, C.-C.; Sheen, J.-M.; Tiao, M.-M.; Tain, Y.-L.; Lin, I.-C.; Lai, Y.-J.; et al. Resveratrol intake during pregnancy and lactation re-programs adiposity and ameliorates leptin resistance in male progeny induced by maternal high-fat/high sucrose plus postnatal high-fat/high sucrose diets via fat metabolism regulation. Lipids Health Dis. 2020, 19, 174. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [Green Version]
- Lappas, M.; Mitton, A.; Lim, R.; Barker, G.; Riley, C.; Permezel, M. SIRT1 Is a Novel Regulator of Key Pathways of Human Labor1. Biol. Reprod. 2011, 84, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.; Liong, S.; Lim, R.; Barker, G.; Lappas, M. Resveratrol ameliorates the chemical and microbial induction of inflammation and insulin resistance in human placenta, adipose tissue and skeletal muscle. PLoS ONE 2017, 12, e0173373. [Google Scholar] [CrossRef] [PubMed]
- Malvasi, A.; Kosmas, I.; Mynbaev, O.A. Can trans resveratrol plus d-chiro-inositol and myo-inositol improve maternal metabolic profile in overweight pregnant patients? Clin. Ther. 2017, 240–247. [Google Scholar] [CrossRef]
- Ding, J.; Kang, Y.; Fan, Y.; Chen, Q. Efficacy of resveratrol to supplement oral nifedipine treatment in pregnancy-induced preeclampsia. Endocr. Connect. 2017, 6, 595–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuente-Martín, E.; Granado, M.; García-Cáceres, C.; Sanchez-Garrido, M.A.; Frago, L.M.; Tena-Sempere, M.; Argente, J.; Chowen, J.A. Early nutritional changes induce sexually dimorphic long-term effects on body weight gain and the response to sucrose intake in adult rats. Metabolism 2012, 61, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Mela, V.; Llorente-Berzal, Á.; Díaz, F.; Argente, J.; Viveros, M.-P.; Chowen, J.A. Maternal Deprivation Exacerbates the Response to a High Fat Diet in a Sexually Dimorphic Manner. PLoS ONE 2012, 7, e48915. [Google Scholar] [CrossRef] [Green Version]
- Tibu, F.; Hill, J.; Sharp, H.; Marshall, K.; Glover, V.; Pickles, A. Evidence for sex differences in fetal programming of physiological stress reactivity in infancy. Dev. Psychopathol. 2014, 26, 879–888. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Model | Species/Sex | Resveratrol Dose and Route | Time | Age at Outcome | Offspring Outcome | Reference |
|---|---|---|---|---|---|---|
| Maternal diabetes | Rat (S-D)/NS | 100 mg/BW/d | 3–12 d GA | ED 12 | ↓ oxidative stress and apoptosis | [61] |
| Maternal diabetes | GDM mouse */NS | 10 mg/BW/d | 4 w pre-preg. and gest. | PND 1 | ↓ BW and ↑ AMPK | [68] |
| Maternal diabetes | Rat (Wi)/NS | 100 mg/BW/d | Gestation (8–12 d) | GD 19 | ↓ oxidative stress | [65] |
| Maternal LPD/CD | Rat (Wi)/M&F | 20 mg/BW/d | Gestation | GD 19 and PND 110 | Sex differences TG and ROS | [29] |
| Maternal HFD | Mouse **/M | 200 mg/BW/d | Preg to PND21 | PNDs 21 and 98 | ↑ BAT browning WAT | [66] |
| Maternal HFD/LFD | Rat (Wi)/ M and F | 50 mg/dl in drinking water | Preg to PND21 | PND 21 | Sex differences ↓BW, Lept, VAT, SCAT | [30] |
| Maternal CD | Rat (Wi)/M | 20 mg/BW/d | Lactation | PND 252 | ↓Plasma cholesterol ↓HMGCR | [67] |
| Maternal HFHS/CD | Rat (S-D)/M | 50 mg/dl in drinking water | Preg to PND21 | PND 180 | ↓BW, adiposity regulates SIRT 1 in RPT Lipid modulation | [70] |
| Maternal HFD | Rat (S-D)/M | 50 mg/dl in drinking water | Preg to PND21 | PND 120 | ↓ RP adiposity Improves leptin dysregulation | [13] |
| Maternal HFD/LFD | Rats (Wi)/ M and F | 50 mg/dl in drinking water | Preg to PND21 | PND 150 | VAT ↓in HFD↑ in LFD ↑ VAT adipocyte size | [31] |
| Maternal WD | Japanese macaque/NS | +0.37% in WD | 3m pre-G to 130 GA | ED130 | ↓ liver lipid and placental inflammation | [62] |
| Maternal WD | Japanese macaque/NS | 0.37% in WD | 3m pre-G to 130 GA | ED 130 | ↑↑ fetal islet vascularity | [63] |
| Maternal HFD/CD | Japanese macaque/NS | 0.37% in WD | 3m pre-G to 130 GA | ED 130 | ↑ Placental fatty acid uptake (DHA) | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ros, P.; Argente, J.; Chowen, J.A. Effects of Maternal Resveratrol Intake on the Metabolic Health of the Offspring. Int. J. Mol. Sci. 2021, 22, 4792. https://doi.org/10.3390/ijms22094792
Ros P, Argente J, Chowen JA. Effects of Maternal Resveratrol Intake on the Metabolic Health of the Offspring. International Journal of Molecular Sciences. 2021; 22(9):4792. https://doi.org/10.3390/ijms22094792
Chicago/Turabian StyleRos, Purificación, Jesús Argente, and Julie A. Chowen. 2021. "Effects of Maternal Resveratrol Intake on the Metabolic Health of the Offspring" International Journal of Molecular Sciences 22, no. 9: 4792. https://doi.org/10.3390/ijms22094792
APA StyleRos, P., Argente, J., & Chowen, J. A. (2021). Effects of Maternal Resveratrol Intake on the Metabolic Health of the Offspring. International Journal of Molecular Sciences, 22(9), 4792. https://doi.org/10.3390/ijms22094792