
Genetic Variation in MicroRNA-423 Promotes Proliferation, Migration, Invasion, and Chemoresistance in Breast Cancer Cells

 , ,
, ,  ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. miR-423 Expression Was Upregulated by Rs6505162:C>A
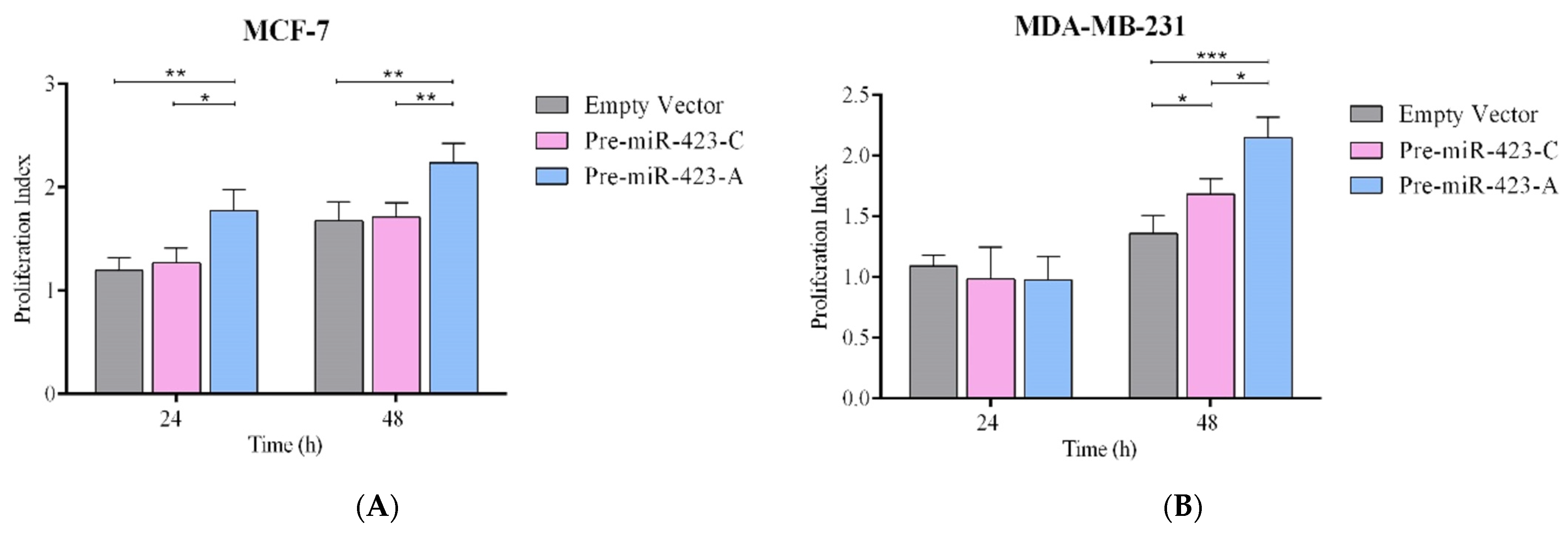
2.2. Pre-miR-423-A Enhanced Breast Cancer Cell Proliferation
2.3. Pre-miR-423-A Expression Promoted Cisplatin Resistance in Breast Cancer Cells
2.4. Pre-miR-423-A Expression Decreased Cisplatin-Induced Apoptosis of Breast Cancer Cells
2.5. Pre-miR-423-A Expression Increased Migration and Invasion in Breast Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. miR-423 Expression Vector Construction and Cell Transfection
4.3. Analysis of miR-423-5p and miR-423-3p Expression by RT-qPCR
4.4. Cell Proliferation Assay
4.5. Cell Viability Assay
4.6. Apoptosis Analysis
4.7. Migration and Invasion Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Government of Chile. Plan Nacional de Cáncer 2018–2028; Department of Comprehensive Management of Cancer and Other Tumors, Undersecretary of Public Health, Ministry of Health: Santiago, Chile, 2019.
- Anglian Breast Cancer Study Group. Prevalence and penetrance of BRCA1 and BRCA2 mutations in a population-based series of breast cancer cases. Br. J. Cancer 2000, 83, 1301–1308. [Google Scholar] [CrossRef]
- Rothe, F.; Ignatiadis, M.; Chaboteaux, C.; Haibe-Kains, B.; Kheddoumi, N.; Majjaj, S.; Badran, B.; Fayyad-Kazan, H.; Desmedt, C.; Harris, A.; et al. Global microRNA expression profiling identifies MiR-210 associated with tumor proliferation, invasion and poor clinical outcome in breast cancer. PLoS ONE 2011, 6, e20980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, F.J.; Chen, K.X. Single-nucleotide polymorphisms among microRNA: Big effects on cancer. Chin. J. Cancer 2011, 30, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erson, A.; Petty, E.M. MicroRNAs in development and disease. Clin. Genet. 2008, 74, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; Downing, J.R. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs-microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- O’Day, E.; Lal, A. MicroRNAs and their target gene networks in breast cancer. Breast Cancer Res. 2010, 12, 201. [Google Scholar] [CrossRef] [Green Version]
- Ryan, B.; Robles, A.; Harris, C.C. Genetic variation in microRNA networks: The implications for cancer research. Nat. Rev. Cancer 2010, 10, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Huang, S.; Wu, S.; Ding, J.; Zhao, Y.; Liang, L.; Tian, Q.; Zha, R.; Zhan, R.; He, X. MicroRNA-423 promotes cell growth and regulates G1/S transition by targeting p21Cip1/Waf1 in hepatocellular carcinoma. Carcinogenesis 2011, 32, 1641–1647. [Google Scholar] [CrossRef]
- Guan, G.; Zhang, D.; Zheng, Y.; Wen, L.; Yu, D.; Lu, Y.; Zhao, Y. microRNA-423-3p promotes tumor progression via modulation of AdipoR2 in laryngeal carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 5683–5691. [Google Scholar] [PubMed]
- Liu, J.; Wang, X.; Yang, X.; Liu, Y.; Shi, Y.; Ren, J.; Guleng, B. miRNA423-5p regulates cell proliferation and invasion by targeting trefoil factor 1 in gastric cancer cells. Cancer Lett. 2014, 347, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Fu, H.; Wang, B.; Zhang, X.; Mao, J.; Li, X.; Wang, M.; Sun, Z.; Qian, H.; Xu, W. Exosomal miR-423-5p targets SUFU to promote cancer growth and metastasis and serves as a novel marker for gastric cancer. Mol. Carcinog. 2018, 57, 1223–1236. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Zhu, X.; Geng, Q.; Xia, L.; Sun, X.; Chen, Y.; Li, W.; Zhou, Z.; Zhan, Y.; Xu, D. The microRNA-423-3p-Bim Axis Promotes Cancer Progression and Activates Oncogenic Autophagy in Gastric Cancer. Mol. Ther. 2017, 25, 1027–1037. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Deng, J.; Sun, Z.M.; Pan, A.P.; Xiang, X.J.; Zhang, L.; Yu, F.; Chen, J.; Sun, Z.; Feng, M.; et al. Interference with the beta-catenin gene in gastric cancer induces changes to the miRNA expression profile. Tumor Biol. 2015, 36, 6973–6983. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zeng, A.; Hu, Q.; Yan, W.; Liu, Y.; You, Y. miR-423-5p contributes to a malignant phenotype and temozolomide chemoresistance in glioblastomas. Neuro-Oncology 2017, 19, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; He, J.; Huang, H.; Peng, R.; Xi, J. MicroRNA-423-3p promotes glioma growth by targeting PANX2. Oncol. Lett. 2018, 16, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Fei, X.; Wang, Z.; Jiang, D.; Chen, H.; Wang, M.; Zhou, S. miR-423-5p knockdown enhances the sensitivity of glioma stem cells to apigenin through the mitochondrial pathway. Tumor Biol. 2017, 39, 1010428317695526. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Sun, S.; Zhai, Y.; Tian, Q.; Zhou, T.; Li, J. miR-423-5p inhibits the proliferation and metastasis of glioblastoma cells by targeting phospholipase C beta 1. Int. J. Clin. Exp. Pathol. 2019, 12, 2941–2950. [Google Scholar]
- Sun, R.; Gong, J.; Li, J.; Ruan, Z.; Yang, X.; Zheng, Y.; Qing, L.; He, X.; Jiang, J.; Peng, Y.; et al. A genetic variant rs13293512 in the promoter of let-7 is associated with an increased risk of breast cancer in Chinese women. Biosci. Rep. 2019, 39, BSR20182079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Li, G.; Zhuang, G.; Sun, S.; Song, Z. Overexpression of microRNA-423-3p indicates poor prognosis and promotes cell proliferation, migration, and invasion of lung cancer. Diagn. Pathol. 2019, 14, 53. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Fan, Z.; McGee, W.; Chen, M.; Kong, R.; Wen, P.; Xiao, T.; Chen, X.; Liu, J.; Zhu, L.; et al. TDP-43 regulates cancer-associated microRNAs. Protein Cell 2018, 9, 848–866. [Google Scholar] [CrossRef]
- Li, J.; Sun, H.; Liu, T.; Kong, J. MicroRNA-423 promotes proliferation, migration and invasion and induces chemoresistance of endometrial cancer cells. Exp. Ther. Med. 2018, 16, 4213–4224. [Google Scholar] [CrossRef]
- Yanokura, M.; Banno, K.; Iida, M.; Irie, H.; Umene, K.; Masuda, K.; Kobayashi, Y.; Tominaga, E.; Aoki, D. MicroRNAS in endometrial cancer: Recent advances and potential clinical applications. EXCLI J. 2015, 14, 190–198. [Google Scholar] [PubMed]
- Jia, W.; Yu, T.; An, Q.; Cao, X.; Pan, H. MicroRNA-423-5p inhibits colon cancer growth by promoting caspase-dependent apoptosis. Exp. Ther. Med. 2018, 16, 1225–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, R.; Goto, Y.; Kojima, S.; Enokida, H.; Chiyomaru, T.; Kinoshita, T.; Sakamoto, S.; Fuse, M.; Nakagawa, M.; Naya, Y.; et al. Tumor-suppressive microRNA-29s inhibit cancer cell migration and invasion via targeting LAMC1 in prostate cancer. Int. J. Oncol. 2014, 45, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Ke, J.; Tian, J.; Li, J.; Gong, Y.; Yang, Y.; Zhu, Y.; Zhang, Y.; Zhong, R.; Chang, J.; Gong, J. Identification of a functional polymorphism affecting microRNA binding in the susceptibility locus 1q25.3 for colorectal cancer. Mol. Carcinog 2017, 56, 2014–2021. [Google Scholar] [CrossRef]
- Li, H.T.; Li, H.T.; Zhang, H.; Chen, Y.; Liu, X.F.; Qian, J. MiR-423-3p enhances cell growth through inhibition of p21Cip1/Waf1 in colorectal cancer. Cell Physiol. Biochem. 2015, 37, 1044–1054. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Lin, T.; Lin, J.; Yang, M.; Shen, Z.; Liu, H.; Zou, Z.; Zheng, Z. Inhibition of miR-423-5p suppressed prostate cancer through targeting GRIM-19. Gene 2019, 688, 93–97. [Google Scholar] [CrossRef]
- Zhao, H.; Gao, A.; Zhang, Z.; Tian, R.; Luo, A.; Li, M.; Zhao, D.; Fu, L.; Fu, L.; Dong, J.-T.; et al. Genetic analysis and preliminary function study of miR-423 in breast cancer. Tumor Biol. 2015, 36, 4763–4771. [Google Scholar] [CrossRef] [PubMed]
- Morales, S.; Gulppi, F.; Gonzalez-Hormazabal, P.; Fernandez-Ramires, R.; Bravo, T.; Reyes, J.M.; Gomez, F.; Waugh, E.; Jara, L. Association of single nucleotide polymorphisms in Pre-miR-27a, Pre-miR-196a2, Pre-miR-423, miR-608 and Pre-miR-618 with breast cancer susceptibility in a South American population. BMC Genet. 2016, 17, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The role and mechanisms of action of microRNAs in cancer drug resistance. Clin. Epigenet. 2019, 11, 25. [Google Scholar] [CrossRef]
- Chen, P.-S.; Su, J.-L.; Hung, M.-C. Dysregulation of microRNAs in cancer. J. Biomed. Sci 2012, 19, 90. [Google Scholar] [CrossRef] [Green Version]
- Salzman, D.W.; Weidhaas, J.B. SNPing cancer in the bud: MicroRNA and microRNA-target site polymorphisms as diagnostic and prognostic biomarkers in cancer. Pharmacol. Ther. 2013, 137, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Nicoloso, M.S.; Sun, H.; Spizzo, R.; Kim, H.; Wickramasinghe, P.; Shimizu, M.; Wojcik, S.E.; Ferdin, J.; Kunej, T.; Xiao, L.; et al. Single-nucleotide polymorphisms inside microRNA target sites influence tumor susceptibility. Cancer Res. 2010, 70, 2789–2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Chendrimada, T.P.; Wang, Q.; Higuchi, M.; Seeburg, P.H.; Shiekhattar, R.; Nishikura, K. Modulation of microRNA processing and expression through RNA editing by ADAR deaminases. Nat. Struct. Mol. Biol. 2006, 13, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Ke, R.; Lv, L.; Zhang, S.; Zhang, F.; Jiang, Y. Functional mechanism and clinical implications of MicroRNA-423 in human cancers. Cancer Med. 2020, 9, 9036–9051. [Google Scholar] [CrossRef]
- Hui, A.B.; Lenarduzzi, M.; Krushel, T.; Waldron, L.; Pintilie, M.; Shi, W.; Perez-Ordonez, B.; Jurisica, I.; O’Sullivan, B.; Waldron, J.; et al. Comprehensive MicroRNA profiling for head and neck squamous cell carcinomas. Clin. Cancer Res. 2010, 16, 1129–1139. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, R.; Zhang, J.; Li, W.; Gao, C.; Liu, J.; Wang, J. Identification of miR-423 and miR-499 polymorphisms on affecting the risk of hepatocellular carcinoma in a large-scale population. Genet. Test. Mol. Biomark. 2014, 18, 516–524. [Google Scholar] [CrossRef] [Green Version]
- McDermott, A.M.; Miller, N.; Wall, D.; Martyn, L.M.; Ball, G.; Sweeney, K.J.; Kerin, M. Identification and validation of oncologic miRNA biomarkers for luminal A-like breast cancer. PLoS ONE 2014, 9, e87032. [Google Scholar] [CrossRef] [Green Version]
- Xing, J.; Wan, S.; Zhou, F.; Qu, F.; Li, B.; Myers, R.E.; Fu, X.; Palazzo, J.P.; He, X.; Chen, Z.; et al. Genetic polymorphisms in pre-microRNA genes as prognostic markers of colorectal cancer. Cancer Epidemiol. Biomark. Prev. 2012, 21, 217–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Zeng, X.; Huang, Y.; Chen, S.; Lin, F.; Yang, G.; Yang, N. miR-423-5p serves as a diagnostic indicator and inhibits the proliferation and invasion of ovarian cancer. Exp. Ther. Med. 2018, 15, 4723–4730. [Google Scholar] [CrossRef]
- Xia, W.; Liu, Y.; Du, Y.; Cheng, T.; Hu, X.; Li, X. MicroRNA-423 Drug Resistance and Proliferation of Breast Cancer Cells by Targeting ZFP36. OncoTargets Ther. 2020, 13, 769–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, T.; Zhao, X.; Li, Y.; Yu, L.; Li, Y.; Zhou, X.; Gong, Q. miR-423 Promotes Breast Cancer Invasion by Activating NF-κB Signaling. OncoTargets Ther. 2020, 13, 5467–5478. [Google Scholar] [CrossRef]
- Tan, H. The association between gene SNPs and cancer predisposition: Correlation or causality? EBioMedicine 2017, 16, 8–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.; Bao, J.; Zhou, X. Genome-wide mutational spectra analysis reveals significant cancer-specific heterogeneity. Sci. Rep. 2015, 5, 12566. [Google Scholar] [CrossRef] [Green Version]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Giannakakou, P. Targeting microtubules for cancer chemotherapy. Curr Med. Chem. Anticancer Agents 2005, 5, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Ohtsukasa, S.; Okabe, S.; Yamashita, H.; Iwai, T.; Sugihara, K. Increased expression of CEA and MHC class I in colorectal cancer cell lines exposed to chemotherapy drugs. J. Cancer Res. Clin. Oncol. 2003, 129, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Henson, R.; Lang, M.; Wehbe, H.; Maheshwari, S.; Mendell, J.T.; Jiang, J.; Schmittgen, T.D.; Patel, T. Involvement of human micro-RNA in growth and response to chemotherapy in human cholangiocarcinoma cell lines. Gastroenterology 2006, 130, 2113–2129. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Kong, W.; He, L.; Zhao, J.-J.; O’Donnell, J.D.; Wang, J.; Wenham, R.M.; Coppola, D.; Kruk, P.A.; Nicosia, S.V.; et al. MicroRNA expression profiling in human ovarian cancer: miR-214 induces cell survival and cisplatin resistance by targeting PTEN. Cancer Res. 2008, 68, 425–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, W.; He, L.; Coppola, M.; Guo, J.; Esposito, N.N.; Coppola, D.; Cheng, J.Q. MicroRNA-155 regulates cell survival, growth, and chemosensitivity by targeting FOXO3a in breast cancer. J. Biol. Chem. 2016, 291, 22855. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Xu, Z.; DiSante, G.; Wright, J.; Wang, M.; Li, Y.; Zhao, Q.; Ren, T.; Ju, X.; Gutman, E.; et al. miR-17/20 sensitization of breast cancer cells to chemotherapy-induced apoptosis requires Akt1. Oncotarget 2014, 5, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Zhi, Y.; Zhou, H.; Mubalake, A.; Chen, Y.; Zhang, B.; Zhang, K.; Chu, X.; Wang, R. Regulation and functions of MicroRNA-149 in human cancers. Cell Prolif. 2018, 51, e12465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zijl, F.; Krupitza, G.; Mikulits, W. Initial steps of metastasis: Cell invasion and endothelial transmigration. Mutat. Res. 2011, 728, 23–34. [Google Scholar] [CrossRef]
- Mir, R.; Al Balawi, I.A.; Duhier, F.A. Duhier, Involvement of microRNA-423 Gene Variability in Breast Cancer Progression in Saudi Arabia. Asian Pac. J. Cancer Prev. 2018, 19, 2581–2589. [Google Scholar]
- Huber, M.A.; Azoitei, N.; Baumann, B.; Grünert, S.; Sommer, A.; Pehamberger, H.; Kraut, N.; Beug, H.; Wirth, T. NF-κB is essential for epithelial-mesenchymal transition and metastasis in a model of breast cancer progression. J. Clin. Investig. 2004, 114, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Farazi, T.A.; Horlings, H.M.; Then Hoeve, J.J.; Mihailovic, A.; Halfwerk, H.; Morozov, P.; Brown, M.; Hafner, M.; Reyal, F.; Van Kouwenhove, M.; et al. MicroRNA sequence and expression analysis in breast tumors by deep sequencing. Cancer Res. 2011, 71, 4443–4453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Pison, S.; Jara, L.; Carrasco, V.; Gutiérrez-Vera, C.; Reyes, J.M.; Gonzalez-Hormazabal, P.; Carreño, L.J.; Tapia, J.C.; Contreras, H.R. Genetic Variation in MicroRNA-423 Promotes Proliferation, Migration, Invasion, and Chemoresistance in Breast Cancer Cells. Int. J. Mol. Sci. 2022, 23, 380. https://doi.org/10.3390/ijms23010380
Morales-Pison S, Jara L, Carrasco V, Gutiérrez-Vera C, Reyes JM, Gonzalez-Hormazabal P, Carreño LJ, Tapia JC, Contreras HR. Genetic Variation in MicroRNA-423 Promotes Proliferation, Migration, Invasion, and Chemoresistance in Breast Cancer Cells. International Journal of Molecular Sciences. 2022; 23(1):380. https://doi.org/10.3390/ijms23010380
Chicago/Turabian StyleMorales-Pison, Sebastian, Lilian Jara, Valentina Carrasco, Cristian Gutiérrez-Vera, José Miguel Reyes, Patricio Gonzalez-Hormazabal, Leandro J. Carreño, Julio C. Tapia, and Héctor R. Contreras. 2022. "Genetic Variation in MicroRNA-423 Promotes Proliferation, Migration, Invasion, and Chemoresistance in Breast Cancer Cells" International Journal of Molecular Sciences 23, no. 1: 380. https://doi.org/10.3390/ijms23010380
APA StyleMorales-Pison, S., Jara, L., Carrasco, V., Gutiérrez-Vera, C., Reyes, J. M., Gonzalez-Hormazabal, P., Carreño, L. J., Tapia, J. C., & Contreras, H. R. (2022). Genetic Variation in MicroRNA-423 Promotes Proliferation, Migration, Invasion, and Chemoresistance in Breast Cancer Cells. International Journal of Molecular Sciences, 23(1), 380. https://doi.org/10.3390/ijms23010380