
Brain-Derived Neurotrophic Factor Suppressed Proinflammatory Cytokines Secretion and Enhanced MicroRNA(miR)-3168 Expression in Macrophages



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
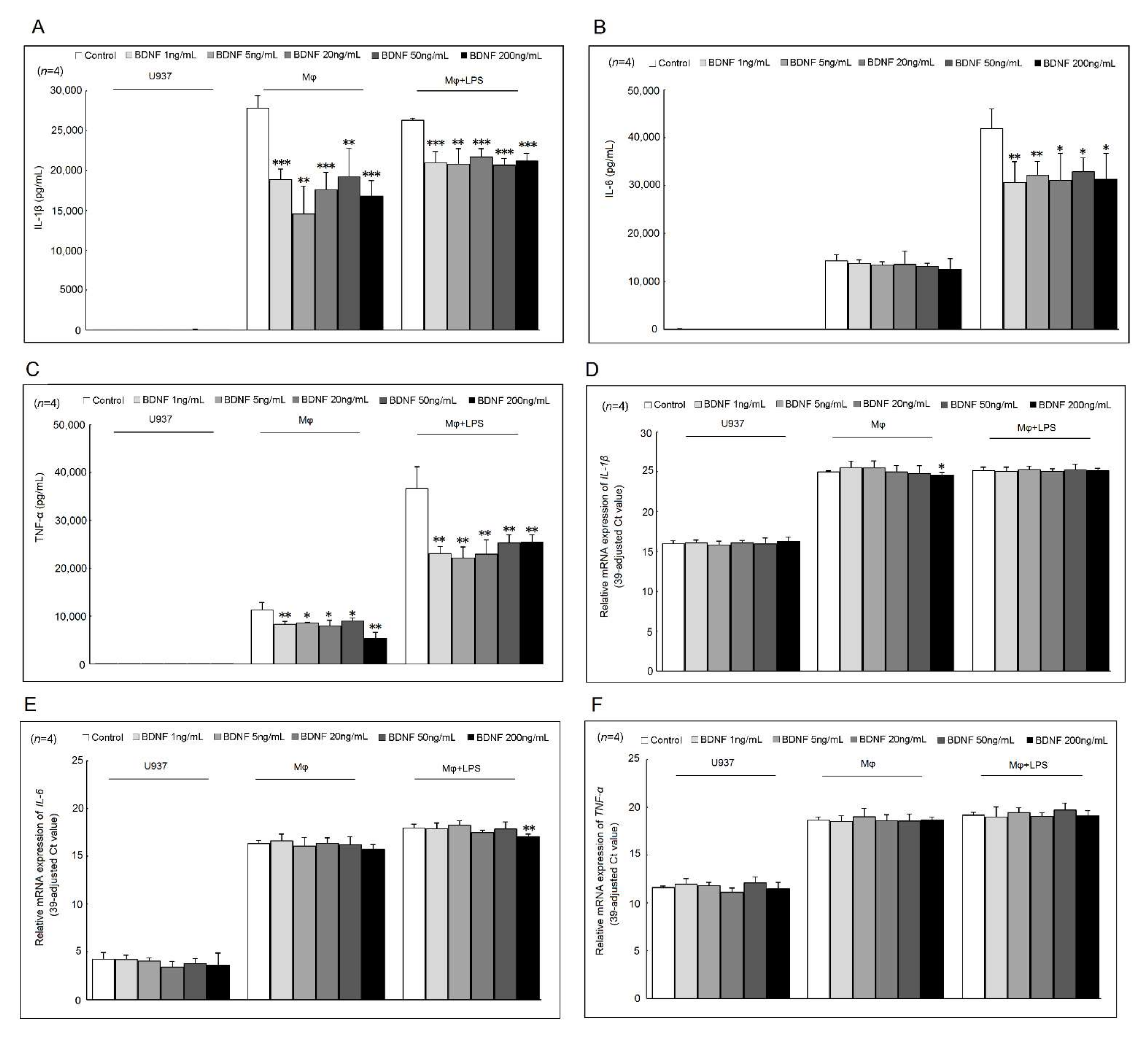
2.1. Effects of BDNF on IL-1β, IL-6, and TNF-α Secretion and mRNA Expression in U937 Cells, Differentiated Macrophages, and Lipopolysaccharide (LPS)-Stimulated Macrophages
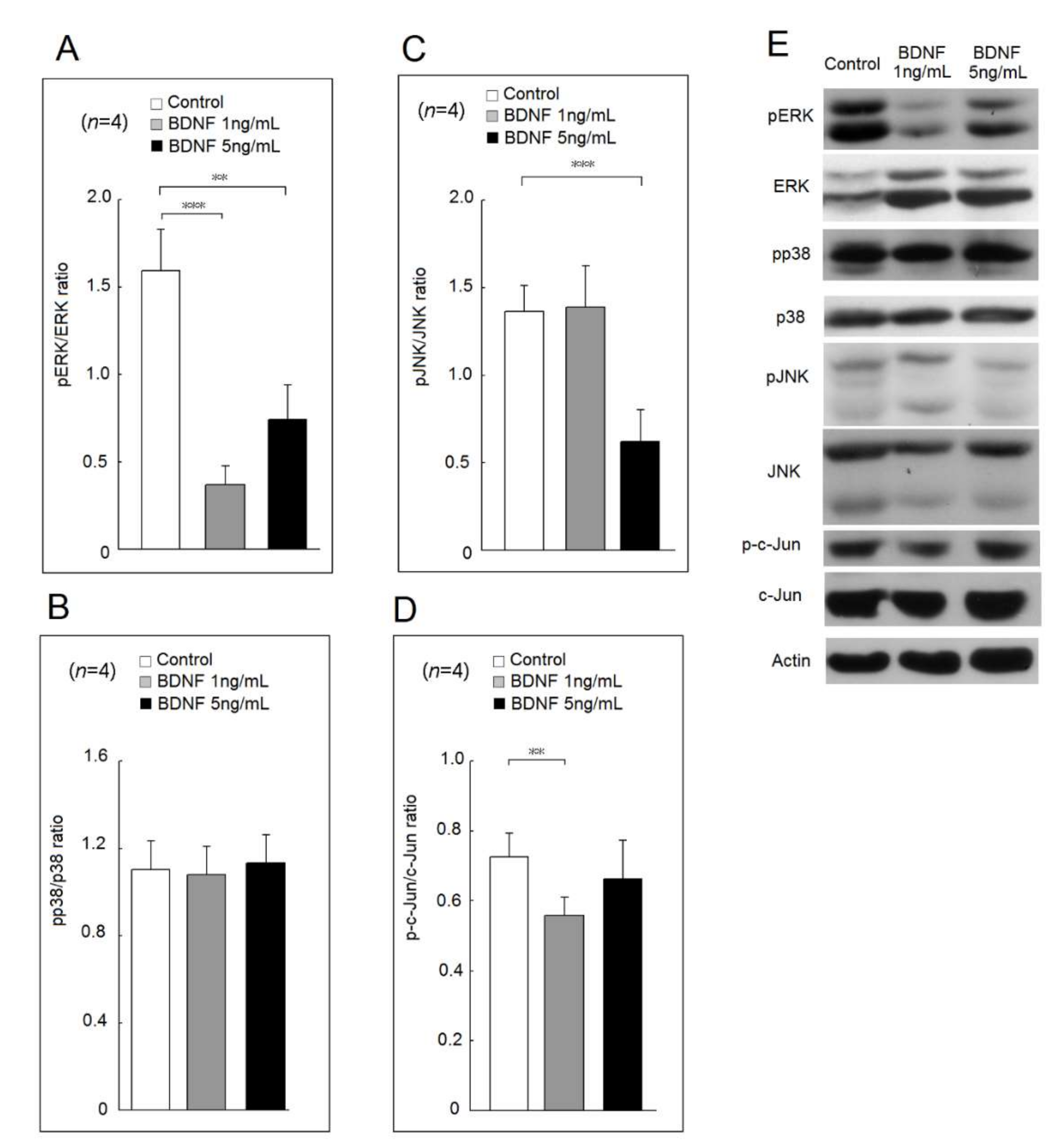
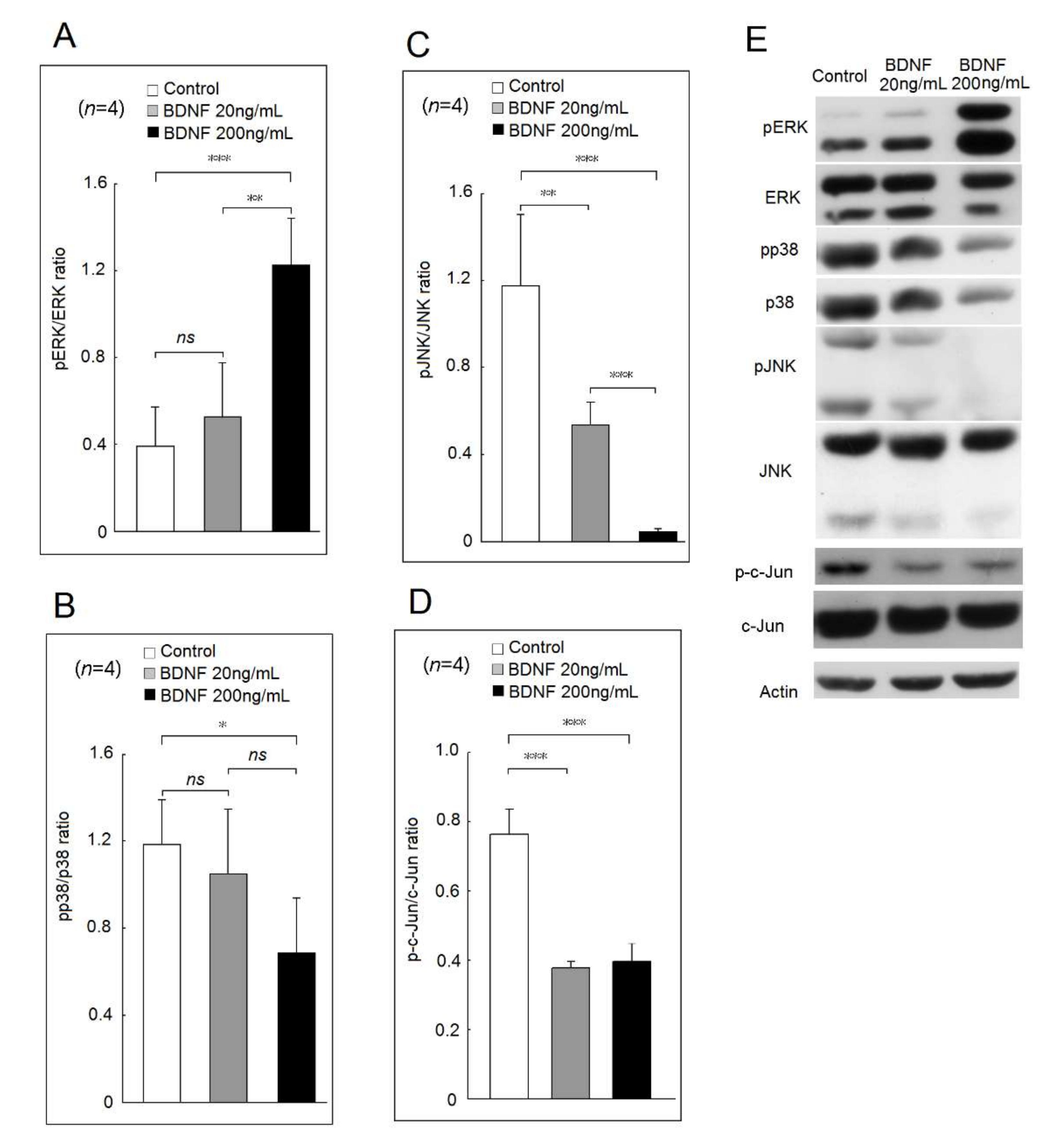
2.2. Effects of BDNF in Mitogen-Activated Protein Kinase (MAPKs) and c-Jun Phosphorylation
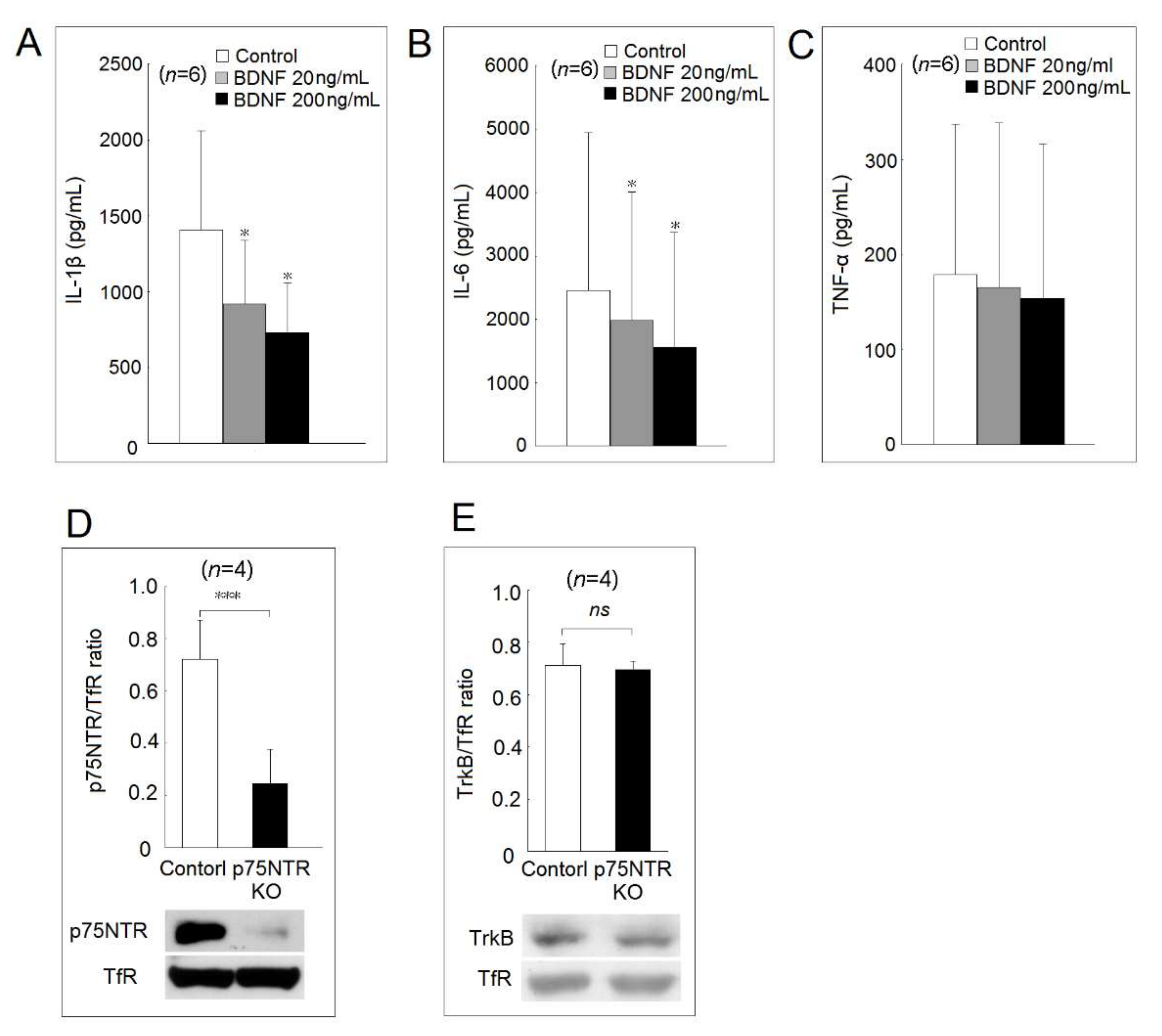
2.3. Effects of BDNF on Proinflammatory Cytokine Secretion in Human Monocyte-Derived Macrophages
2.4. The Expression of Two BDNF Receptors, TrkB and p75NTR Expression in p75NTR Knockout U937 Cells
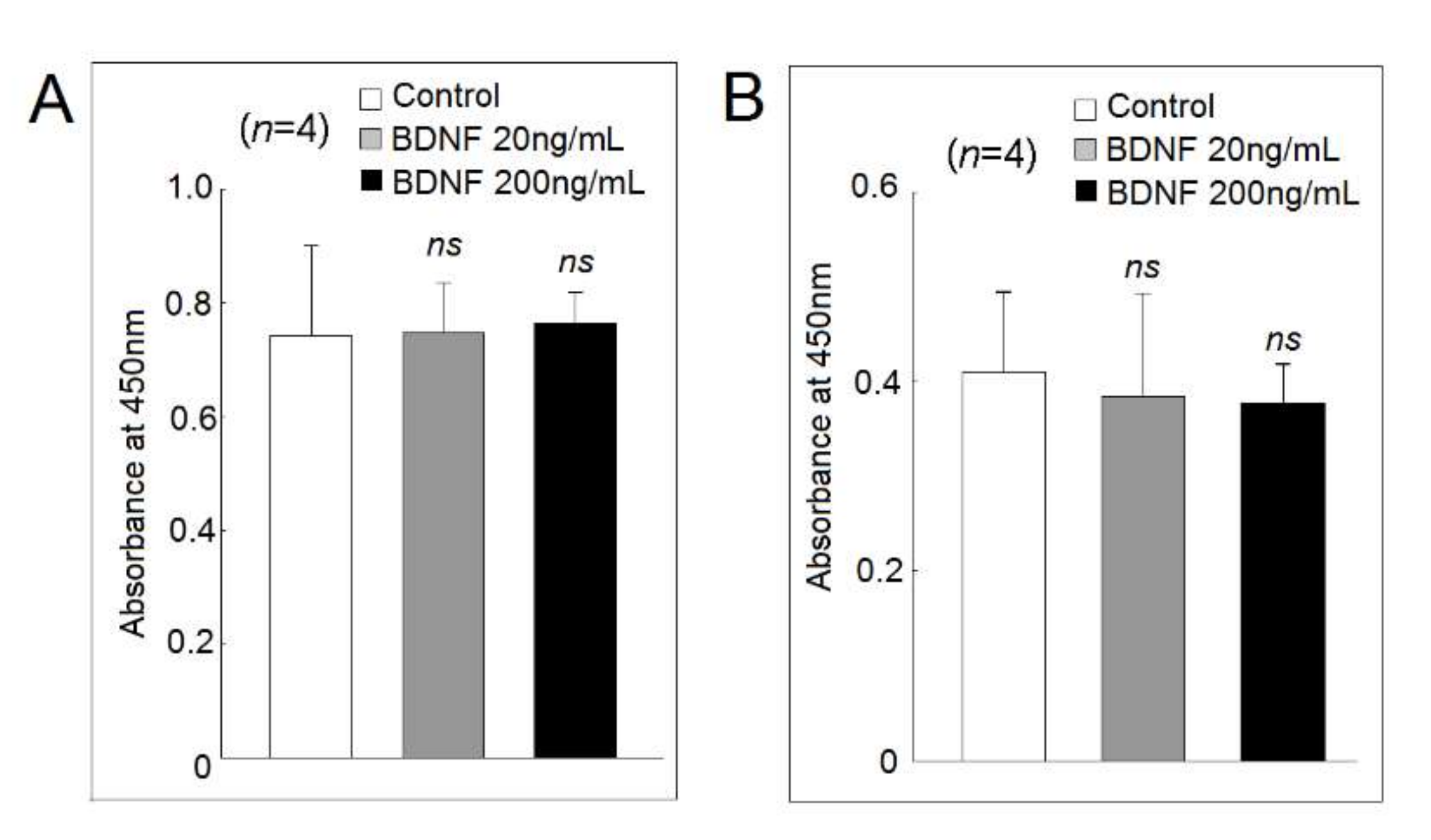
2.5. Effect of BDNF on U937 Cell Viability and Proliferation
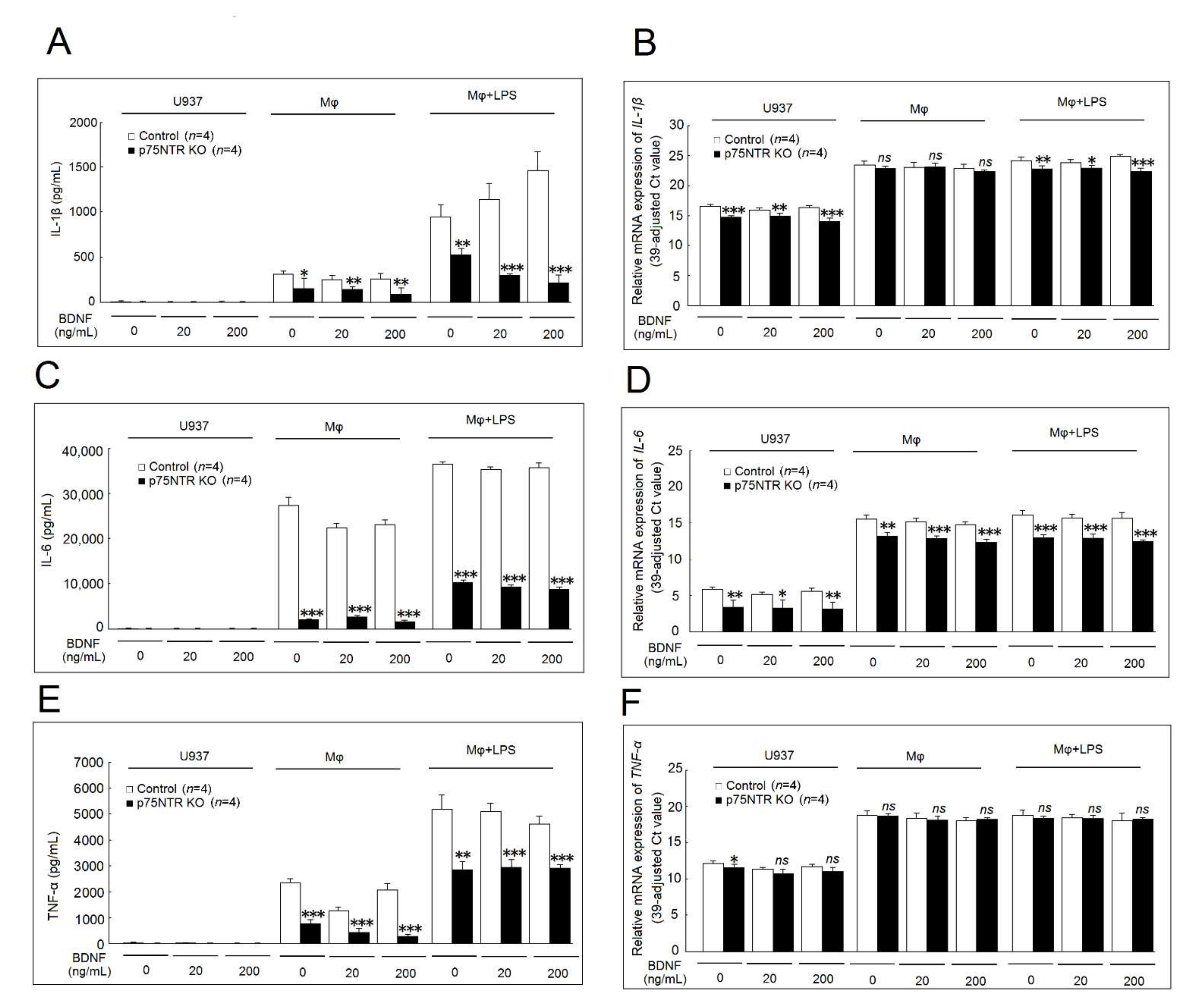
2.6. Effects of p75NTR Knockout on IL-1β, IL-6, and TNF-α Secretion and mRNA Expression
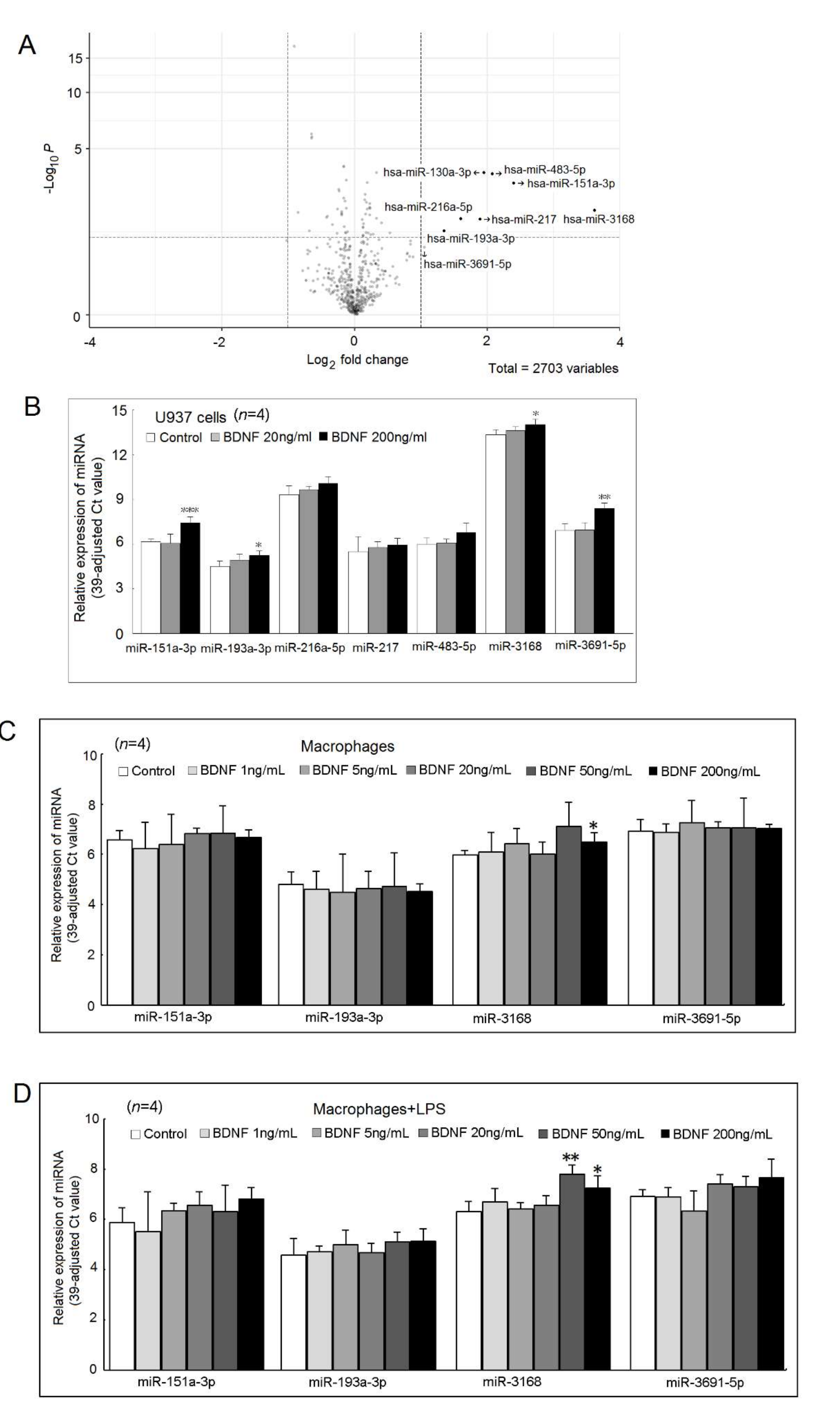
2.7. Expression of miRNAs Regulated by BDNF
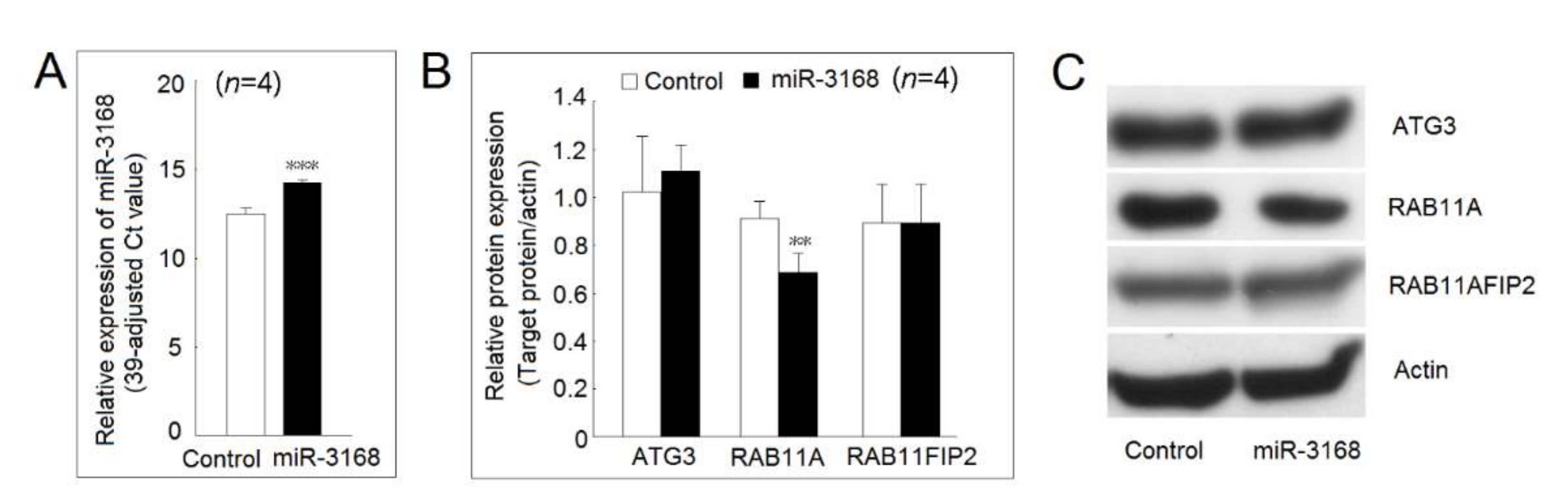
2.8. Identification of Protein Expression Regulated by miR-3168
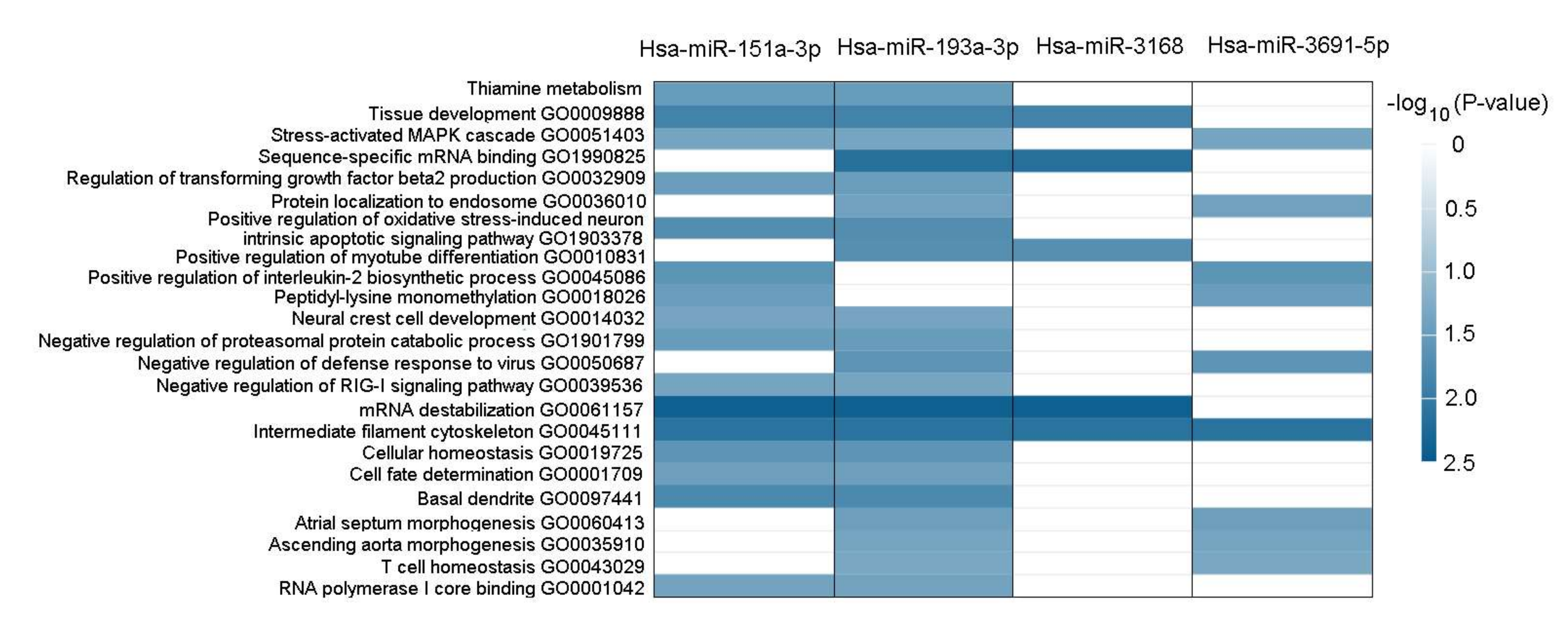
2.9. Functional Annotation of BDNF-Regulated miRNAs
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of Human Monocyte-Derived Macrophages
4.3. Measurement of Gene Expression Levels by RT-PCR
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Cell Viability and Proliferation Using the Mitochondrial Dehydrogenase Cleavage Assay
4.6. Preparation of Surface Membrane Extract
4.7. Western Blot Analysis
4.8. NGS for miRNAs
4.9. Measurement of miRNAs Expression
4.10. Preparation of p75NTR Knockout U937 Cells
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mariga, A.; Mitre, M.; Chao, M.-V. Consequences of brain-derived neurotrophic factor withdrawal in CNS neurons and implications in disease. Neurobiol. Dis. 2017, 97, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A key factor with multipotent impact on brain signaling and synaptic plasticity. Cell Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Li, W.; Lv, L.; Zhang, Z.; Zhan, X. BDNF as a biomarker in diagnosis and evaluation of treatment for schizophrenia and depression. Discov. Med. 2018, 26, 127–136. [Google Scholar] [PubMed]
- Najjar, S.; Pearlman, D.M.; Alper, K.; Najjar, A.; Devinsky, O. Neuroinflammation and psychiatric illness. J. Neuroinflamm. 2013, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Fauchais, A.L.; Boumediene, A.; Lalloue, F.; Gondran, G.; Loustaud-Ratti, V.; Vidal, E.; Jauberteau, M.O. Brain-derived neurotrophic factor and nerve growth factor correlate with T-cell activation in primary Sjogren’s syndrome. Scand. J. Rheumatol. 2009, 38, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Fauchais, A.L.; Lise, M.C.; Marget, P.; Lapeybie, F.X.; Bezanahary, H.; Martel, C.; Dumonteil, S.; Sparsa, A.; Lalloué, F.; Ly, K.; et al. Serum and lymphocytic neurotrophins profiles in systemic lupus erythematosus: A case-control study. PLoS ONE 2013, 8, e79414. [Google Scholar]
- Lai, N.-S.; Yu, H.-C.; Huang Tseng, H.-Y.; Hsu, C.-W.; Huang, H.-B.; Lu, M.-C. Increased serum levels of brain-derived neurotrophic factor contribute to inflammatory responses in patients with rheumatoid arthritis. Int. J. Mol. Sci. 2021, 22, 1841. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Yang, C.; Wang, J.; Hou, X.; Zhao, S.; Li, Y.; Yang, P. Peripheral blood brain-derived neurotrophic factor level and tyrosine kinase B expression on T lymphocytes in systemic lupus erythematosus: Implications for systemic involvement. Cytokine 2019, 123, 154764. [Google Scholar] [CrossRef]
- Kerschensteiner, M.; Gallmeier, E.; Behrens, L.; Leal, V.V.; Misgeld, T.; Klinkert, W.E.; Kolbeck, R.; Hoppe, E.; Oropeza-Wekerle, R.L.; Bartke, I.; et al. Activated human T cells, B cells, and monocytes produce brain-derived neurotrophic factor in vitro and in inflammatory brain lesions: A neuroprotective role of inflammation? J. Exp. Med. 1999, 189, 865–870. [Google Scholar] [CrossRef]
- Pedard, M.; Quirié, A.; Tessier, A.; Garnier, P.; Totoson, P.; Demougeot, C.; Marie, C. A reconciling hypothesis centred on brain-derived neurotrophic factor to explain neuropsychiatric manifestations in rheumatoid arthritis. Rheumatology 2021, 60, 1608–1619. [Google Scholar] [CrossRef]
- Kloc, M.; Uosef, A.; Kubiak, J.Z.; Ghobrial, R.M. Macrophage proinflammatory responses to microorganisms and transplanted organs. Int. J. Mol. Sci. 2020, 21, 9669. [Google Scholar] [CrossRef]
- Ji, X.-C.; Dang, Y.-Y.; Gao, H.-Y.; Wang, Z.T.; Gao, M.; Yang, Y.; Zhang, H.-T.; Xu, R.-X. Local injection of Lenti-BDNF at the lesion site promotes M2 macrophage polarization and inhibits inflammatory response after spinal cord injury in mice. Cell Mol. Neurobiol. 2015, 35, 881–890. [Google Scholar] [CrossRef]
- Garaci, E.; Caroleo, M.C.; Aloe, L.; Aquaro, S.; Piacentini, M.; Costa, N.; Amendola, A.; Micera, A.; Caliò, R.; Perno, C.F.; et al. Nerve growth factor is an autocrine factor essential for the survival of macrophages infected with HIV. Proc. Natl. Acad. Sci. USA 1999, 96, 14013–14018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Suárez, O.; Hannestad, J.; Esteban, I.; Sainz, R.; Naves, F.J.; Vega, J.A. Expression of the TrkB neurotrophin receptor by thymic macrophages. Immunology 1998, 94, 235–241. [Google Scholar] [CrossRef]
- Bi, C.; Fu, Y.; Li, B. Brain-derived neurotrophic factor alleviates diabetes mellitus-accelerated atherosclerosis by promoting M2 polarization of macrophages through repressing the STAT3 pathway. Cell. Signal. 2020, 70, 109569. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Lian, D.; Wu, J.; Liu, Y.; Zhu, M.; Sun, J.; He, D.; Li, L. Brain-derived neurotrophic factor reduces inflammation and hippocampal apoptosis in experimental Streptococcus pneumoniae meningitis. J. Neuroinflamm. 2017, 14, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Deng, G.; Huang, H. The activation of BDNF reduced inflammation in a spinal cord injury model by TrkB/p38 MAPK signaling. Exp. Ther. Med. 2019, 17, 1688–1696. [Google Scholar] [CrossRef] [Green Version]
- Düsedau, H.P.; Kleveman, J.; Figueiredo, C.A.; Biswas, A.; Steffen, J.; Kliche, S.; Haak, S.; Zagrebelsky, M.; Korte, M.; Dunay, I.R. p75(NTR) regulates brain mononuclear cell function and neuronal structure in Toxoplasma infection-induced neuroinflammation. Glia 2019, 67, 193–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Mattingly, A.; Lin, A.; Sacramento, J.; Mannent, L.; Castel, M.N.; Canolle, B.; Delbary-Gossart, S.; Ferzaz, B.; Morganti, J.M.; et al. A novel antagonist of p75NTR reduces peripheral expansion and CNS trafficking of pro-inflammatory monocytes and spares function after traumatic brain injury. J. Neuroinflamm. 2016, 13, 88. [Google Scholar] [CrossRef] [Green Version]
- Minnone, G.; De Benedetti, F.; Bracci-Laudiero, L. NGF and its receptors in the regulation of inflammatory response. Int. J. Mol. Sci. 2017, 18, 1028. [Google Scholar] [CrossRef]
- Williams, K.S.; Killebrew, D.A.; Clary, G.P.; Seawell, J.A.; Meeker, R.B. Differential regulation of macrophage phenotype by mature and pro-nerve growth factor. J. Neuroimmunol. 2015, 285, 76–93. [Google Scholar] [CrossRef] [Green Version]
- Berenbaum, F.; Blanco, F.J.; Guermazi, A.; Miki, K.; Yamabe, T.; Viktrup, L.; Junor, R.; Carey, W.; Brown, M.T.; West, C.R.; et al. Subcutaneous tanezumab for osteoarthritis of the hip or knee: Efficacy and safety results from a 24-week randomised phase III study with a 24-week follow-up period. Ann. Rheum. Dis. 2020, 79, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Kelliny, S.; Lam, H.-Y.; Parikh, A.; Wang, Y.-J.; Bobrovskaya, L.; Upton, R.; Zhou, X.-F. Preclinical study of the pharmacokinetics of p75ECD-Fc, a novel human recombinant protein for treatment of Alzheimer’s disease, in Sprague Dawley rats. Curr. Drug Metab. 2020, 21, 235–244. [Google Scholar] [CrossRef]
- Reefman, E.; Kay, J.-G.; Wood, S.-M.; Offenhäuser, C.; Brown, D.-L.; Roy, S.; Stanley, A.C.; Low, P.C.; Manderson, A.P.; Stow, J.L.; et al. Cytokine secretion is distinct from secretion of cytotoxic granules in NK cells. J. Immunol. 2010, 184, 4852–4862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, R.Z.; Kay, J.-G.; Sangermani, D.G.; Stow, J.L. A role for the phagosome in cytokine secretion. Science 2005, 310, 1492–1495. [Google Scholar] [CrossRef] [Green Version]
- Lai, N.-S.; Yu, H.-C.; Tung, C.-H.; Huang, K.-Y.; Huang, H.-B.; Lu, M.-C. Increased peptidylarginine deiminases expression during the macrophage differentiation and participated inflammatory responses. Arthritis Res. Ther. 2019, 21, 108. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.-C.; Lai, N.-S.; Yin, W.-Y.; Yu, H.-C.; Huang, H.-B.; Tung, C.-H.; Huang, K.-Y.; Yu, C.-L. Anti-citrullinated protein antibodies activated ERK1/2 and JNK mitogen-activated protein kinases via binding to surface-expressed citrullinated GRP78 on mononuclear cells. J. Clin. Immunol. 2013, 33, 558–566. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Friendländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Kern, F.; Fehlmann, T.; Solomon, J.; Schwed, L.; Grammes, N.; Backes, C.; Van Keuren-Jensen, K.; Craig, D.W.; Meese, E.; Keller, A. miEAA 2.0: Integrating multi-species microRNA enrichment analysis and workflow management systems. Nucleic Acids Res. 2020, 48, W521–W528. [Google Scholar] [CrossRef]
- Lu, M.-C.; Lai, N.-S.; Chen, H.-C.; Yu, H.-C.; Huang, K.-Y.; Tung, C.-H.; Huang, H.-B.; Yu, C.-L. Decreased microRNA(miR)-145 and increased miR-224 expression in T cells from patients with systemic lupus erythematosus involved in lupus immunopathogenesis. Clin. Exp. Immunol. 2013, 171, 91–99. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.-C.; Huang, H.-B.; Huang Tseng, H.-Y.; Lu, M.-C. Brain-Derived Neurotrophic Factor Suppressed Proinflammatory Cytokines Secretion and Enhanced MicroRNA(miR)-3168 Expression in Macrophages. Int. J. Mol. Sci. 2022, 23, 570. https://doi.org/10.3390/ijms23010570
Yu H-C, Huang H-B, Huang Tseng H-Y, Lu M-C. Brain-Derived Neurotrophic Factor Suppressed Proinflammatory Cytokines Secretion and Enhanced MicroRNA(miR)-3168 Expression in Macrophages. International Journal of Molecular Sciences. 2022; 23(1):570. https://doi.org/10.3390/ijms23010570
Chicago/Turabian StyleYu, Hui-Chun, Hsien-Bin Huang, Hsien-Yu Huang Tseng, and Ming-Chi Lu. 2022. "Brain-Derived Neurotrophic Factor Suppressed Proinflammatory Cytokines Secretion and Enhanced MicroRNA(miR)-3168 Expression in Macrophages" International Journal of Molecular Sciences 23, no. 1: 570. https://doi.org/10.3390/ijms23010570
APA StyleYu, H.-C., Huang, H.-B., Huang Tseng, H.-Y., & Lu, M.-C. (2022). Brain-Derived Neurotrophic Factor Suppressed Proinflammatory Cytokines Secretion and Enhanced MicroRNA(miR)-3168 Expression in Macrophages. International Journal of Molecular Sciences, 23(1), 570. https://doi.org/10.3390/ijms23010570