
Type B CTD Proteins Secreted by the Type IX Secretion System Associate with PorP-like Proteins for Cell Surface Anchorage

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. PorP, PorE and PG1035 Form a Complex
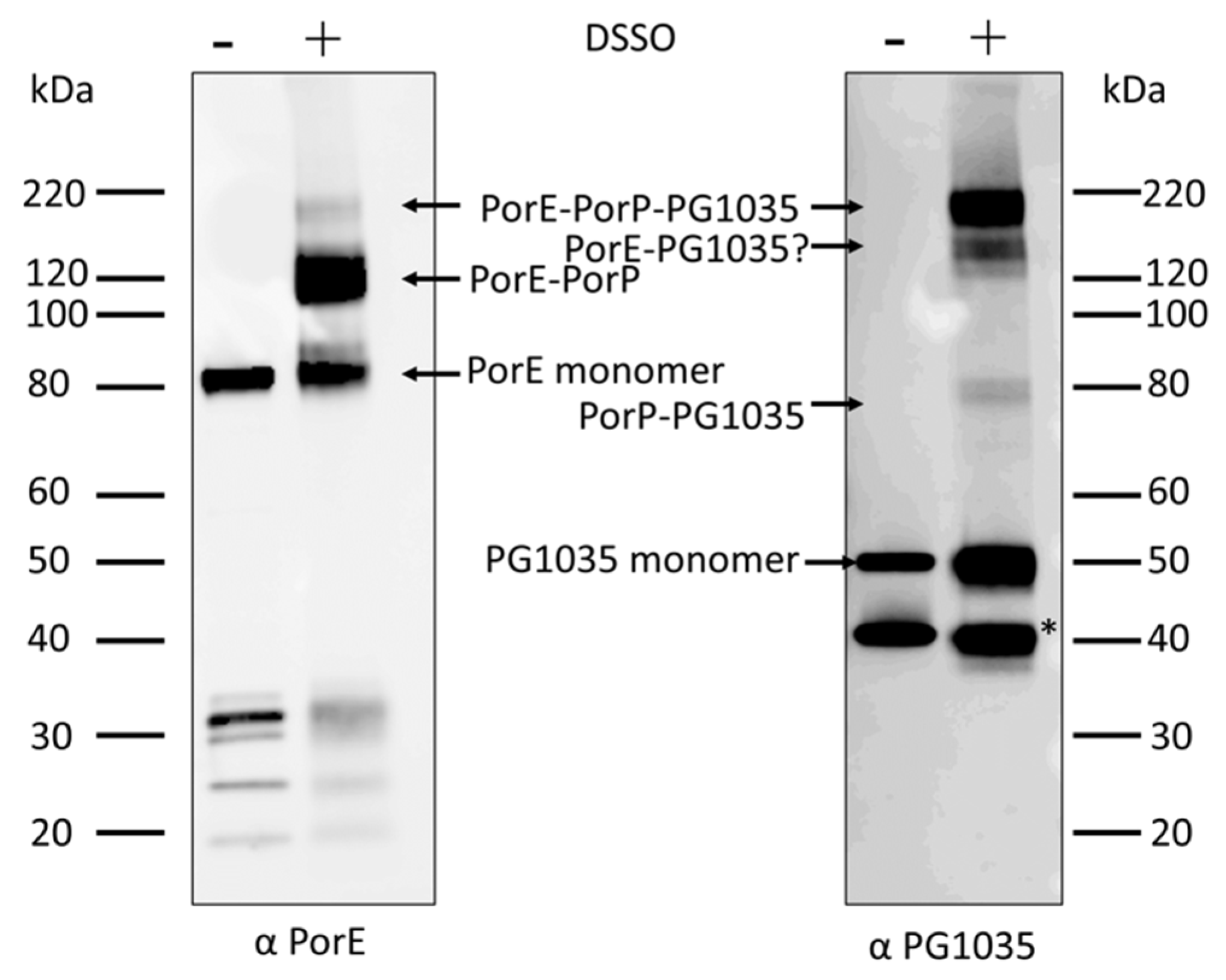
2.2. Chemical Cross-Linking Indicates Various Pairwise Interactions
2.3. PG1035 Complex Formation Is Independent of PorU and PorV
2.4. The PorP-PG1035 Complex Can Form Independently of PorE
2.5. PG1035 Is a Non-Essential Component of the T9SS
2.6. PG1035 Is Localised on the Cell Surface and Is Not Modified by A-LPS
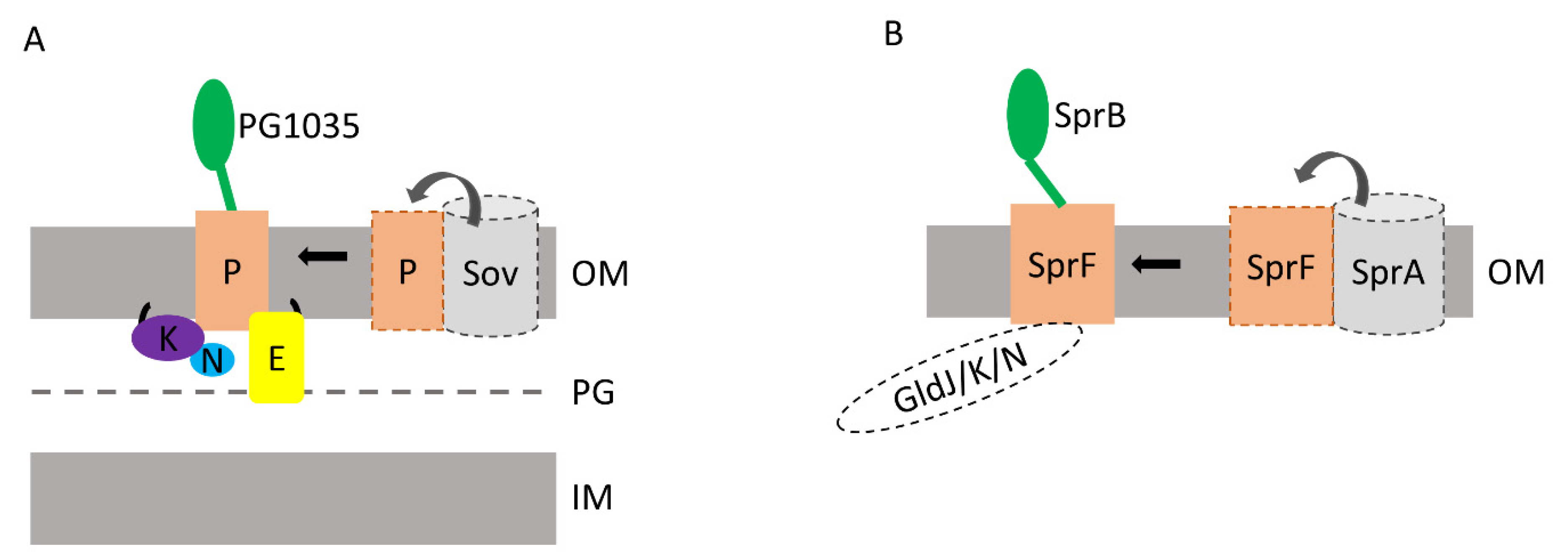
2.7. The F. johnsoniae Motility Protein SprB Binds to the PorP Homologue SprF
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Generation of P. gingivalis pg1035 Deletion Mutant
4.3. Generation of Strep-Tag II PorE
4.4. PG1035 Antibody
4.5. Blue Native Gel Electrophoresis
4.6. Immunoblot Analysis
4.7. Co-Immunoprecipitation
4.8. LC-MS/MS
4.9. Cross-Linking
4.10. Fluorescence Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McBride, M.J.; Zhu, Y. Gliding motility and por secretion system genes are widespread among members of the phylum Bacteroidetes. J. Bacteriol. 2013, 195, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veith, P.D.; Glew, M.D.; Gorasia, D.G.; Reynolds, E.C. Type IX secretion: The generation of bacterial cell surface coatings involved in virulence, gliding motility and the degradation of complex biopolymers. Mol. Microbiol. 2017, 106, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Lasica, A.M.; Ksiazek, M.; Madej, M.; Potempa, J. The type IX secretion system (T9SS): Highlights and recent insights into its structure and function. Front. Cell. Infect. Microbiol. 2017, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- McBride, M.J. Bacteroidetes gliding motility and the type IX secretion system. Microbiol. Spectr. 2019, 7, 7. [Google Scholar] [CrossRef]
- Gorasia, D.G.; Veith, P.D.; Reynolds, E.C. The type IX secretion system: Advances in structure, function and organisation. Microorganisms 2020, 8, 1173. [Google Scholar] [CrossRef]
- Silva, I.L.; Cascales, E. Molecular strategies underlying Porphyromonas gingivalis virulence. J. Mol. Biol. 2021, 433, 166836. [Google Scholar] [CrossRef]
- Sato, K.; Naito, M.; Yukitake, H.; Hirakawa, H.; Shoji, M.; McBride, M.J.; Rhodes, R.G.; Nakayama, K. A protein secretion system linked to Bacteroidete gliding motility and pathogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, A.; Johnston, J.J.; van Baaren, J.M.; McBride, M.J. Flavobacterium johnsoniae GldK, GldL, GldM, and SprA are required for secretion of the cell surface gliding motility adhesins SprB and RemA. J. Bacteriol. 2013, 195, 3201–3212. [Google Scholar] [CrossRef] [Green Version]
- Kharade, S.S.; McBride, M.J. Flavobacterium johnsoniae chitinase ChiA is required for chitin utilization and is secreted by the type IX secretion system. J. Bacteriol. 2014, 196, 961–970. [Google Scholar] [CrossRef] [Green Version]
- Shoji, M.; Sato, K.; Yukitake, H.; Kondo, Y.; Narita, Y.; Kadowaki, T.; Naito, M.; Nakayama, K. Por secretion system-dependent secretion and glycosylation of Porphyromonas gingivalis hemin-binding protein 35. PLoS ONE 2011, 6, e21372. [Google Scholar] [CrossRef] [Green Version]
- Seers, C.A.; Slakeski, N.; Veith, P.D.; Nikolof, T.; Chen, Y.-Y.; Dashper, S.G.; Reynolds, E.C. The RgpB C-terminal domain has a role in attachment of RgpB to the outer membrane and belongs to a novel C-terminal-domain family found in Porphyromonas gingivalis. J. Bacteriol. 2006, 188, 6376–6386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veith, P.D.; Nor Muhammad, N.A.; Dashper, S.G.; Likić, V.A.; Gorasia, D.G.; Chen, D.; Byrne, S.J.; Catmull, D.V.; Reynolds, E.C. Protein substrates of a novel secretion system are numerous in the Bacteroidetes phylum and have in common a cleavable C-terminal secretion signal, extensive post-translational modification, and cell-surface attachment. J. Proteome Res. 2013, 12, 4449–4461. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.S.; Zhu, Y.; Brendel, C.J.; McBride, M.J. Diverse C-terminal sequences involved in Flavobacterium johnsoniae protein secretion. J. Bacteriol. 2017, 199, e00884-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, S.S.; Johnston, J.J.; Zhu, Y.; Hying, Z.T.; McBride, M.J. The carboxy-terminal region of Flavobacterium johnsoniae SprB facilitates its secretion by the type IX secretion system and propulsion by the gliding motility machinery. J. Bacteriol. 2019, 201, e00218-19. [Google Scholar] [CrossRef] [Green Version]
- Nakane, D.; Sato, K.; Wada, H.; McBride, M.J.; Nakayama, K. Helical flow of surface protein required for bacterial gliding motility. Proc. Natl. Acad. Sci. USA 2013, 110, 11145–11150. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, A.; Berg, H.C. A molecular rack and pinion actuates a cell-surface adhesin and enables bacterial gliding motility. Sci. Adv. 2020, 6, eaay6616. [Google Scholar] [CrossRef] [Green Version]
- Glew, M.D.; Veith, P.D.; Peng, B.; Chen, Y.-Y.; Gorasia, D.G.; Yang, Q.; Slakeski, N.; Chen, D.; Moore, C.; Crawford, S.; et al. PG0026 is the C-terminal signal peptidase of a novel secretion system of Porphyromonas gingivalis. J. Biol. Chem. 2012, 287, 24605–24617. [Google Scholar] [CrossRef] [Green Version]
- Gorasia, D.G.; Veith, P.D.; Chen, D.; Seers, C.A.; Mitchell, H.A.; Chen, Y.-Y.; Glew, M.D.; Dashper, S.G.; Reynolds, E.C. Porphyromonas gingivalis type IX secretion substrates are cleaved and modified by a sortase-like mechanism. PLOS Pathog. 2015, 11, e1005152. [Google Scholar] [CrossRef]
- Heath, J.E.; Seers, C.A.; Veith, P.D.; Butler, C.A.; Nor Muhammad, N.A.; Chen, Y.-Y.; Slakeski, N.; Peng, B.; Zhang, L.; Dashper, S.G.; et al. PG1058 is a novel multidomain protein component of the bacterial type IX secretion system. PLoS ONE 2016, 11, e0164313. [Google Scholar] [CrossRef] [Green Version]
- Lasica, A.M.; Goulas, T.; Mizgalska, D.; Zhou, X.; De Diego, I.; Ksiazek, M.; Madej, M.; Guo, Y.; Guevara, T.; Nowak, M.; et al. Structural and functional probing of PorZ, an essential bacterial surface component of the type-IX secretion system of human oral-microbiomic Porphyromonas gingivalis. Sci. Rep. 2016, 6, 37708. [Google Scholar] [CrossRef]
- Saiki, K.; Konishi, K. Identification of a novel Porphyromonas gingivalis outer membrane protein, PG534, required for the production of active gingipains. FEMS Microbiol. Lett. 2010, 310, 168–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, M.; Tominaga, T.; Shoji, M.; Nakayama, K. PGN_0297 is an essential component of the type IX secretion system (T9SS) in Porphyromonas gingivalis: Tn-seq analysis for exhaustive identification of T9SS-related genes. Microbiol. Immunol. 2019, 63, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadowaki, T.; Yukitake, H.; Naito, M.; Sato, K.; Kikuchi, Y.; Kondo, Y.; Shoji, M.; Nakayama, K. A two-component system regulates gene expression of the type IX secretion component proteins via an ECF sigma factor. Sci. Rep. 2016, 6, 23288. [Google Scholar] [CrossRef] [PubMed]
- Lauber, F.; Deme, J.C.; Lea, S.M.; Berks, B.C. Type 9 secretion system structures reveal a new protein transport mechanism. Nature 2018, 564, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Gorasia, D.G.; Lunar Silva, I.; Butler, C.A.; Chabalier, M.; Doan, T.; Cascales, E.; Veith, P.D.; Reynolds, E.C. Protein interactome analysis of the type IX secretion system identifies PorW as the missing link between the PorK/N ring complex and the Sov translocon. Microbiol. Spectr. 2022, 10, e01602-21. [Google Scholar] [CrossRef]
- Gorasia, D.G.; Veith, P.D.; Hanssen, E.G.; Glew, M.D.; Sato, K.; Yukitake, H.; Nakayama, K.; Reynolds, E.C. Structural Insights into the PorK and PorN components of the Porphyromonas gingivalis type IX secretion system. PLoS Pathog. 2016, 12, e1005820. [Google Scholar] [CrossRef] [Green Version]
- Vincent, M.S.; Canestrari, M.J.; Leone, P.; Stathopulos, J.; Ize, B.; Zoued, A.; Cambillau, C.; Kellenberger, C.; Roussel, A.; Cascales, E. Characterization of the Porphyromonas gingivalis type IX secretion trans-envelope PorKLMNP core complex. J. Biol. Chem. 2017, 292, 3252–3261. [Google Scholar] [CrossRef] [Green Version]
- Hennell James, R.; Deme, J.C.; Kjær, A.; Alcock, F.; Silale, A.; Lauber, F.; Johnson, S.; Berks, B.C.; Lea, S.M. Structure and mechanism of the proton-driven motor that powers type 9 secretion and gliding motility. Nat. Microbiol. 2021, 6, 221–233. [Google Scholar] [CrossRef]
- Glew, M.D.; Veith, P.D.; Chen, D.; Gorasia, D.G.; Peng, B.; Reynolds, E.C. PorV is an outer membrane shuttle protein for the type IX secretion system. Sci. Rep. 2017, 7, 8790. [Google Scholar] [CrossRef]
- Veith, P.D.; Shoji, M.; O’Hair, R.A.J.; Leeming, M.G.; Nie, S.; Glew, M.D.; Reid, G.E.; Nakayama, K.; Reynolds, E.C. Type IX secretion system cargo proteins are glycosylated at the C terminus with a novel linking sugar of the Wbp/Vim pathway. mBio 2020, 11, e01497-20. [Google Scholar] [CrossRef]
- Trinh, N.T.T.; Tran, H.Q.; Van Dong, Q.; Cambillau, C.; Roussel, A.; Leone, P. Crystal structure of type IX secretion system PorE C-terminal domain from Porphyromonas gingivalis in complex with a peptidoglycan fragment. Sci. Rep. 2020, 10, 7384. [Google Scholar] [CrossRef] [PubMed]
- Kharade, S.S.; McBride, M.J. Flavobacterium johnsoniae PorV is required for secretion of a subset of proteins targeted to the type IX secretion system. J. Bacteriol. 2015, 197, 147–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorasia, D.G.; Glew, M.D.; Veith, P.D.; Reynolds, E.C. Quantitative proteomic analysis of the type IX secretion system mutants in Porphyromonas gingivalis. Mol. Oral Microbiol. 2020, 35, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Zeytuni, N.; Zarivach, R. Structural and functional discussion of the tetra-trico-peptide repeat, a protein interaction module. Structure 2012, 20, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Kopec, K.O.; Lupas, A.N. β-Propeller blades as ancestral peptides in protein evolution. PLoS ONE 2013, 8, e77074. [Google Scholar] [CrossRef] [Green Version]
- Veith, P.D.; Gorasia, D.G.; Reynolds, E.C. Towards defining the outer membrane proteome of Porphyromonas gingivalis. Mol. Oral Microbiol. 2021, 36, 25–36. [Google Scholar] [CrossRef]
- Braun, T.F.; McBride, M.J. Flavobacterium johnsoniae GldJ is a lipoprotein that is required for gliding motility. J. Bacteriol. 2005, 187, 2628–2637. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.J.; Shrivastava, A.; McBride, M.J. Untangling Flavobacterium johnsoniae gliding motility and protein secretion. J. Bacteriol. 2018, 200, e00362-17. [Google Scholar] [CrossRef] [Green Version]
- Veith, P.D.; Glew, M.D.; Gorasia, D.G.; Cascales, E.; Reynolds, E.C. The type IX secretion system and its role in bacterial function and pathogenesis. J. Dent. Res. 2021, 101, 374–383. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Peng, B.; Yang, Q.; Glew, M.D.; Veith, P.D.; Cross, K.J.; Goldie, K.N.; Chen, D.; O’Brien-Simpson, N.; Dashper, S.; et al. The outer membrane protein LptO is essential for the O-deacylation of LPS and the co-ordinated secretion and attachment of A-LPS and CTD proteins in Porphyromonas gingivalis. Mol. Microbiol. 2011, 79, 1380–1401. [Google Scholar] [CrossRef]
- McBride, M.J.; Kempf, M.J. Development of techniques for the genetic manipulation of the gliding bacterium Cytophaga johnsonae. J. Bacteriol. 1996, 178, 583–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, M.A.; Thickett, A.; Slaney, J.M.; Rangarajan, M.; Aduse-Opoku, J.; Shepherd, P.; Paramonov, N.; Hounsell, E.F. Variable carbohydrate modifications to the catalytic chains of the RgpA and RgpB proteases of Porphyromonas gingivalis W50. Infect. Immun. 1999, 67, 3816–3823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortz, E.; Krogh, T.N.; Vorum, H.; Gorg, A. Improved silver staining protocols for high sensitivity protein identification using matrix-assisted laser desorption/ionization-time of flight analysis. Proteomics 2001, 1, 1359–1363. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Glew, M.D.; Gorasia, D.G.; McMillan, P.J.; Butler, C.A.; Veith, P.D.; Reynolds, E.C. Complementation in trans of Porphyromonas gingivalis lipopolysaccharide biosynthetic mutants demonstrates lipopolysaccharide exchange. J. Bacteriol. 2021, 203, e00631-20. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | WT | porE | |
|---|---|---|---|
| LFQ | iBAQ (%) | iBAQ (%) | |
| PG1035 | 7266 | 100 | 100 |
| PorE | 589 | 17 | 0 |
| PorP | 1082 | 31 | 14 |
| PorK | 3 | 0.1 | 0.1 |
| PorN | 3 | 0.2 | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorasia, D.G.; Seers, C.A.; Heath, J.E.; Glew, M.D.; Soleimaninejad, H.; Butler, C.A.; McBride, M.J.; Veith, P.D.; Reynolds, E.C. Type B CTD Proteins Secreted by the Type IX Secretion System Associate with PorP-like Proteins for Cell Surface Anchorage. Int. J. Mol. Sci. 2022, 23, 5681. https://doi.org/10.3390/ijms23105681
Gorasia DG, Seers CA, Heath JE, Glew MD, Soleimaninejad H, Butler CA, McBride MJ, Veith PD, Reynolds EC. Type B CTD Proteins Secreted by the Type IX Secretion System Associate with PorP-like Proteins for Cell Surface Anchorage. International Journal of Molecular Sciences. 2022; 23(10):5681. https://doi.org/10.3390/ijms23105681
Chicago/Turabian StyleGorasia, Dhana G., Christine A. Seers, Jacqueline E. Heath, Michelle D. Glew, Hamid Soleimaninejad, Catherine A. Butler, Mark J. McBride, Paul D. Veith, and Eric C. Reynolds. 2022. "Type B CTD Proteins Secreted by the Type IX Secretion System Associate with PorP-like Proteins for Cell Surface Anchorage" International Journal of Molecular Sciences 23, no. 10: 5681. https://doi.org/10.3390/ijms23105681
APA StyleGorasia, D. G., Seers, C. A., Heath, J. E., Glew, M. D., Soleimaninejad, H., Butler, C. A., McBride, M. J., Veith, P. D., & Reynolds, E. C. (2022). Type B CTD Proteins Secreted by the Type IX Secretion System Associate with PorP-like Proteins for Cell Surface Anchorage. International Journal of Molecular Sciences, 23(10), 5681. https://doi.org/10.3390/ijms23105681