
Inhibition of Respiratory Syncytial Virus Infection by Small Non-Coding RNA Fragments

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
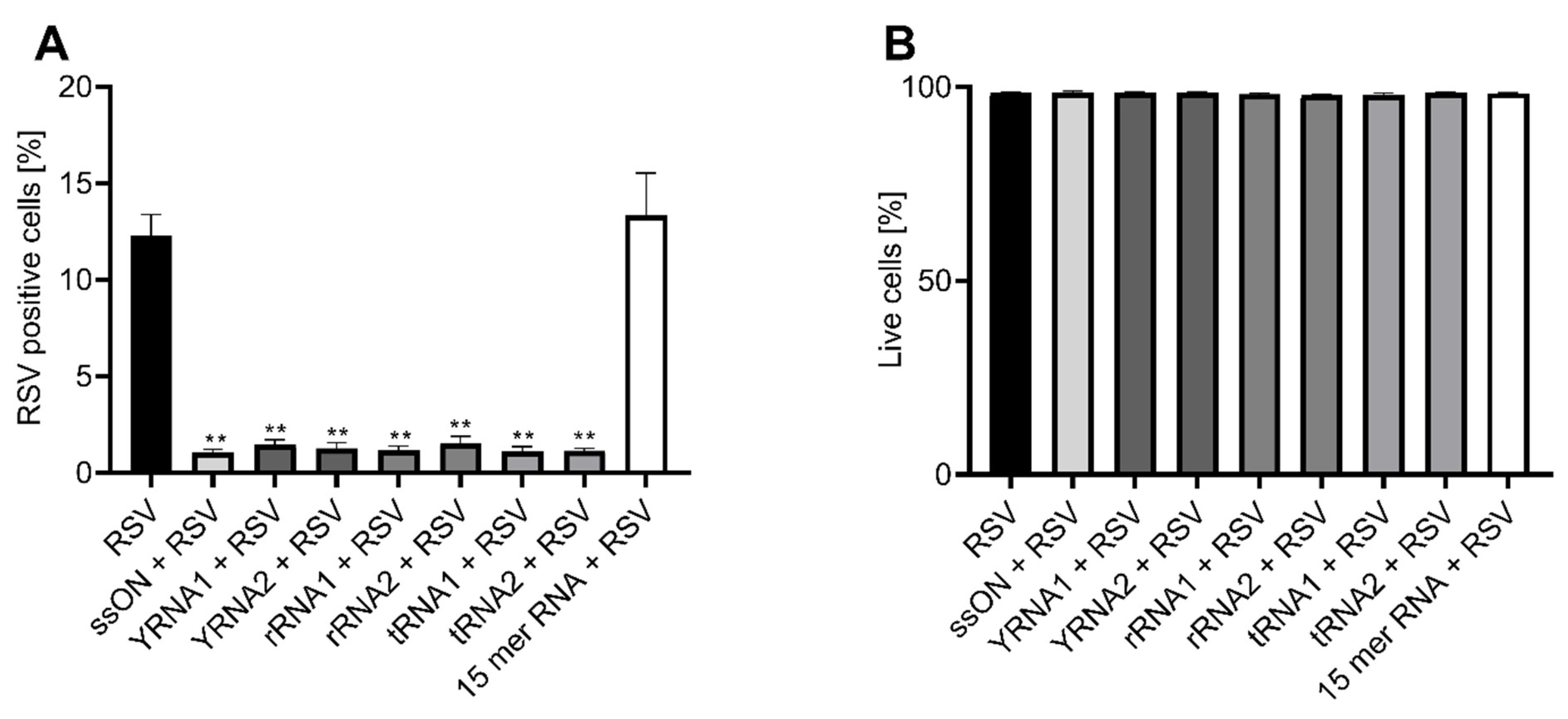
2.1. Naturally Occurring Small Non-Coding RNAs Inhibit RSV Infection In Vitro
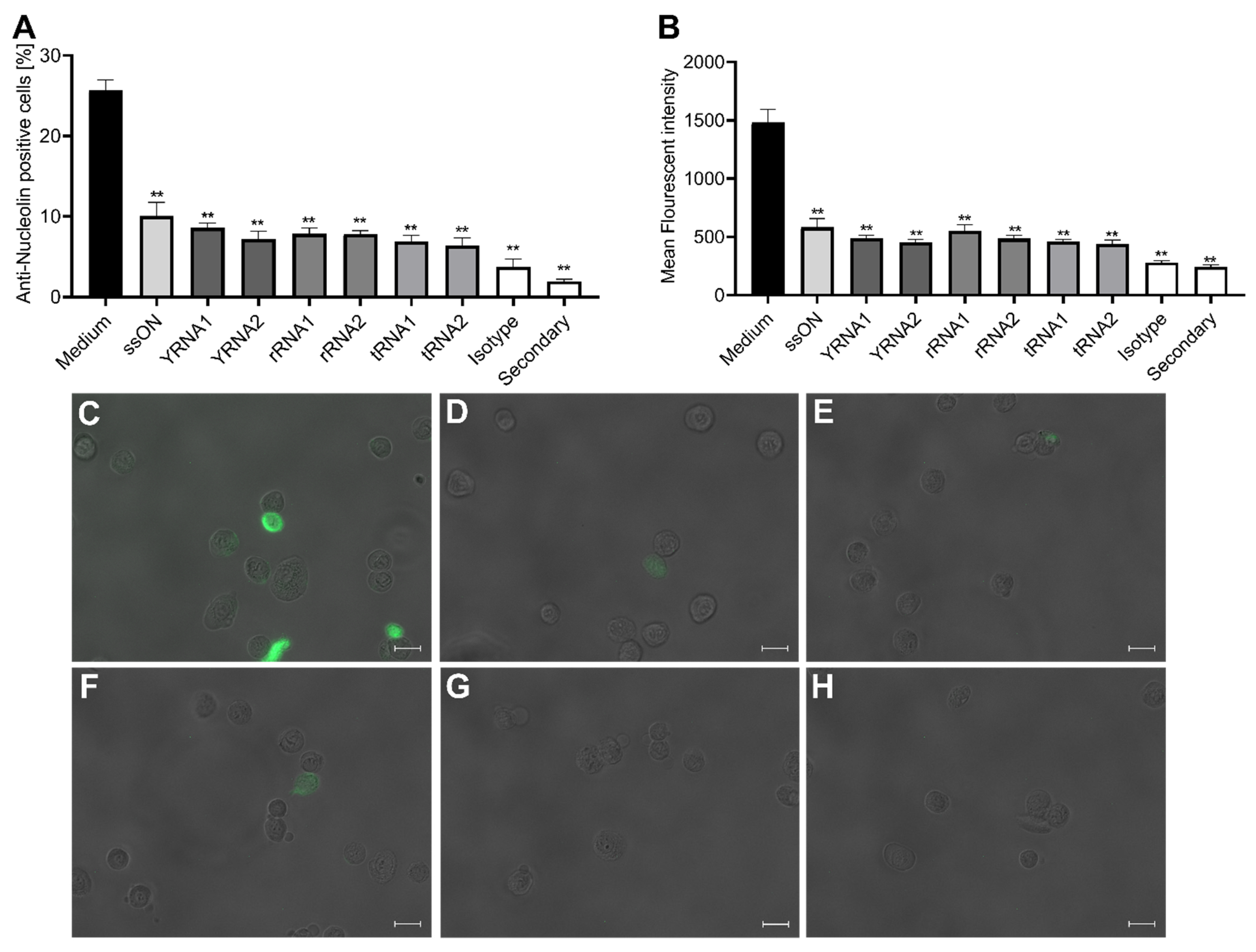
2.2. Small Non-Coding RNAs Bind to the RSV Entry Co-Receptor Nucleolin
3. Discussion
4. Materials and Methods
4.1. Cells and Virus
4.2. Oligonucleotides
4.3. RSV Infection and Analysis Using Flow Cytometry
4.4. Nucleolin Binding Assay
4.5. qPCR
4.6. TCID50 Assay
4.7. ELISA
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global Burden of Acute Lower Respiratory Infections Due to Respiratory Syncytial Virus in Young Children: A Systematic Review and Meta-Analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef] [Green Version]
- Stamm, P.; Sagoschen, I.; Weise, K.; Plachter, B.; Münzel, T.; Gori, T.; Vosseler, M. Influenza and RSV Incidence during COVID-19 Pandemic—an Observational Study from in-Hospital Point-of-Care Testing. Med. Microbiol. Immunol. 2021, 210, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Van Summeren, J.; Meijer, A.; Aspelund, G.; Casalegno, J.S.; Erna, G.; Hoang, U.; Lina, B.; de Lusignan, S.; Teirlinck, A.C.; VRS study group in Lyon; et al. Low Levels of Respiratory Syncytial Virus Activity in Europe during the 2020/21 Season: What Can We Expect in the Coming Summer and Autumn/Winter? Eurosurveillance 2021, 26, 2100639. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.L.; Fearns, R.; Graham, B.S. Respiratory Syncytial Virus: Virology, Reverse Genetics, and Pathogenesis of Disease. In Challenges and Opportunities for Respiratory Syncytial Virus Vaccines; Anderson, L.J., Graham, B.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 372, pp. 3–38. ISBN 978-3-642-38918-4. [Google Scholar]
- Bont, L.; Versteegh, J.; Swelsen, W.T.N.; Heijnen, C.J.; Kavelaars, A.; Brus, F.; Draaisma, J.M.T.; Pekelharing-Berghuis, M.; van Diemen-Steenvoorde, R.A.A.M.; Kimpen, J.L.L. Natural Reinfection with Respiratory Syncytial Virus Does Not Boost Virus-Specific T-Cell Immunity. Pediatr. Res. 2002, 52, 363–367. [Google Scholar] [CrossRef]
- Hall, C.B.; Walsh, E.E.; Long, C.E.; Schnabel, K.C. Immunity to and Frequency of Reinfection with Respiratory Syncytial Virus. J. Infect. Dis. 1991, 163, 693–698. [Google Scholar] [CrossRef]
- Antunes, K.H.; Becker, A.; Franceschina, C.; do Nascimento de Freitas, D.; Lape, I.; da Cunha, M.D.; Leitão, L.; Rigo, M.M.; Pinto, L.A.; Stein, R.T.; et al. Respiratory Syncytial Virus Reduces STAT3 Phosphorylation in Human Memory CD8 T Cells Stimulated with IL-21. Sci. Rep. 2019, 9, 17766. [Google Scholar] [CrossRef] [Green Version]
- Heylen, E.; Neyts, J.; Jochmans, D. Drug Candidates and Model Systems in Respiratory Syncytial Virus Antiviral Drug Discovery. Biochem. Pharmacol. 2017, 127, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shadman, K.A.; Wald, E.R. A Review of Palivizumab and Emerging Therapies for Respiratory Syncytial Virus. Expert Opin. Biol. Ther. 2011, 11, 1455–1467. [Google Scholar] [CrossRef]
- Gutiérrez-Ortega, A.; Sánchez-Hernández, C.; Gómez-García, B. Respiratory Syncytial Virus Glycoproteins Uptake Occurs through Clathrin-Mediated Endocytosis in a Human Epithelial Cell Line. Virol. J. 2008, 5, 127. [Google Scholar] [CrossRef] [Green Version]
- Tayyari, F.; Marchant, D.; Moraes, T.J.; Duan, W.; Mastrangelo, P.; Hegele, R.G. Identification of Nucleolin as a Cellular Receptor for Human Respiratory Syncytial Virus. Nat. Med. 2011, 17, 1132–1135. [Google Scholar] [CrossRef]
- Griffiths, C.D.; Bilawchuk, L.M.; McDonough, J.E.; Jamieson, K.C.; Elawar, F.; Cen, Y.; Duan, W.; Lin, C.; Song, H.; Casanova, J.-L.; et al. IGF1R Is an Entry Receptor for Respiratory Syncytial Virus. Nature 2020, 583, 615–619. [Google Scholar] [CrossRef]
- Järver, P.; Dondalska, A.; Poux, C.; Sandberg, A.; Bergenstråhle, J.; Sköld, A.E.; Dereuddre-Bosquet, N.; Martinon, F.; Pålsson, S.; Zaghloul, E.; et al. Single-Stranded Nucleic Acids Regulate TLR3/4/7 Activation through Interference with Clathrin-Mediated Endocytosis. Sci. Rep. 2018, 8, 15841. [Google Scholar] [CrossRef] [PubMed]
- Pålsson, S.A.; Dondalska, A.; Bergenstråhle, J.; Rolfes, C.; Björk, A.; Sedano, L.; Power, U.F.; Rameix-Welti, M.-A.; Lundeberg, J.; Wahren-Herlenius, M.; et al. Single-Stranded Oligonucleotide-Mediated Inhibition of Respiratory Syncytial Virus Infection. Front. Immunol. 2020, 11, 3202. [Google Scholar] [CrossRef]
- Wu, W.; Choi, E.-J.; Lee, I.; Lee, Y.S.; Bao, X. Non-Coding RNAs and Their Role in Respiratory Syncytial Virus (RSV) and Human Metapneumovirus (HMPV) Infections. Viruses 2020, 12, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beermann, J.; Piccoli, M.-T.; Viereck, J.; Thum, T. Non-Coding RNAs in Development and Disease: Background, Mechanisms, and Therapeutic Approaches. Physiol. Rev. 2016, 96, 1297–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perreault, J.; Perreault, J.-P.; Boire, G. Ro-Associated Y RNAs in Metazoans: Evolution and Diversification. Mol. Biol. Evol. 2007, 24, 1678–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripp, R.A.; Tompkins, S.M. (Eds.) Roles of host gene and non-coding RNA expression in virus infection. In Current Topics in Microbiology and Immunology; Springer International Publishing: Cham, Switzerland, 2018; Volume 419, ISBN 978-3-030-05368-0. [Google Scholar]
- Francisco-Garcia, A.S.; Garrido-Martín, E.M.; Rupani, H.; Lau, L.C.K.; Martinez-Nunez, R.T.; Howarth, P.H.; Sanchez-Elsner, T. Small RNA Species and MicroRNA Profiles Are Altered in Severe Asthma Nanovesicles from Broncho Alveolar Lavage and Associate with Impaired Lung Function and Inflammation. Non Coding RNA 2019, 5, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Järver, P.; O’Donovan, L.; Gait, M.J. A Chemical View of Oligonucleotides for Exon Skipping and Related Drug Applications. Nucleic Acid Ther. 2014, 24, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Meng, M.; Schmidtgall, B.; Ducho, C. Enhanced Stability of DNA Oligonucleotides with Partially Zwitterionic Backbone Structures in Biological Media. Molecules 2018, 23, 2941. [Google Scholar] [CrossRef] [Green Version]
- Rebane, A.; Roomere, H.; Metspalu, A. Locations of Several Novel 2′-O-Methylated Nucleotides in Human 28S RRNA. BMC Mol. Biol. 2002, 3, 1. [Google Scholar] [CrossRef]
- Somme, J.; Van Laer, B.; Roovers, M.; Steyaert, J.; Versées, W.; Droogmans, L. Characterization of Two Homologous 2′-O-Methyltransferases Showing Different Specificities for Their TRNA Substrates. RNA 2014, 20, 1257–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyzaniak, M.A.; Zumstein, M.T.; Gerez, J.A.; Picotti, P.; Helenius, A. Host Cell Entry of Respiratory Syncytial Virus Involves Macropinocytosis Followed by Proteolytic Activation of the F Protein. PLoS Pathog. 2013, 9, e1003309. [Google Scholar] [CrossRef] [Green Version]
- Poux, C.; Dondalska, A.; Bergenstråhle, J.; Pålsson, S.; Contreras, V.; Arasa, C.; Järver, P.; Albert, J.; Busse, D.C.; LeGrand, R.; et al. A Single-Stranded Oligonucleotide Inhibits Toll-Like Receptor 3 Activation and Reduces Influenza A (H1N1) Infection. Front. Immunol. 2019, 10, 2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheleznyakova, G.Y.; Piket, E.; Needhamsen, M.; Hagemann-Jensen, M.; Ekman, D.; Han, Y.; James, T.; Khademi, M.; Al Nimer, F.; Scicluna, P.; et al. Small Noncoding RNA Profiling across Cellular and Biofluid Compartments and Their Implications for Multiple Sclerosis Immunopathology. Proc. Natl. Acad. Sci. USA 2021, 118, e2011574118. [Google Scholar] [CrossRef]
- Guglas, K.; Kołodziejczak, I.; Kolenda, T.; Kopczyńska, M.; Teresiak, A.; Sobocińska, J.; Bliźniak, R.; Lamperska, K. YRNAs and YRNA-Derived Fragments as New Players in Cancer Research and Their Potential Role in Diagnostics. Int. J. Mol. Sci. 2020, 21, 5682. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Ptashkin, R.N.; Chen, Y.; Cheng, Z.; Liu, G.; Phan, T.; Deng, X.; Zhou, J.; Lee, I.; Lee, Y.S.; et al. Respiratory Syncytial Virus Utilizes a TRNA Fragment to Suppress Antiviral Responses Through a Novel Targeting Mechanism. Mol. Ther. 2015, 23, 1622–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrangelo, P.; Chin, A.A.; Tan, S.; Jeon, A.H.; Ackerley, C.A.; Siu, K.K.; Lee, J.E.; Hegele, R.G. Identification of RSV Fusion Protein Interaction Domains on the Virus Receptor, Nucleolin. Viruses 2021, 13, 261. [Google Scholar] [CrossRef] [PubMed]
- Abdelmohsen, K.; Gorospe, M. RNA-Binding Protein Nucleolin in Disease. RNA Biol. 2012, 9, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Mastrangelo, P.; Hegele, R.G. The RSV Fusion Receptor: Not What Everyone Expected It to Be. Microbes Infect. 2012, 14, 1205–1210. [Google Scholar] [CrossRef]
- Saxena, A.; Rorie, C.J.; Dimitrova, D.; Daniely, Y.; Borowiec, J.A. Nucleolin Inhibits Hdm2 by Multiple Pathways Leading to P53 Stabilization. Oncogene 2006, 25, 7274–7288. [Google Scholar] [CrossRef] [Green Version]
- Tonello, F.; Massimino, M.L.; Peggion, C. Nucleolin: A Cell Portal for Viruses, Bacteria, and Toxins. Cell. Mol. Life Sci. 2022, 79, 271. [Google Scholar] [CrossRef] [PubMed]
- Tosar, J.P.; Cayota, A. Extracellular TRNAs and TRNA-Derived Fragments. RNA Biol. 2020, 17, 1149–1167. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA Delivery by Extracellular Vesicles in Mammalian Cells and Its Applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lee, I.; Ren, J.; Ajay, S.S.; Lee, Y.S.; Bao, X. Identification and Functional Characterization of TRNA-Derived RNA Fragments (TRFs) in Respiratory Syncytial Virus Infection. Mol. Ther. 2013, 21, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Villenave, R.; Broadbent, L.; Douglas, I.; Lyons, J.D.; Coyle, P.V.; Teng, M.N.; Tripp, R.A.; Heaney, L.G.; Shields, M.D.; Power, U.F. Induction and Antagonism of Antiviral Responses in Respiratory Syncytial Virus-Infected Pediatric Airway Epithelium. J. Virol. 2015, 89, 12309–12318. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Eldfjell, Y.; Fromm, B.; Estivill, X.; Biryukova, I.; Friedländer, M.R. MiRTrace Reveals the Organismal Origins of MicroRNA Sequencing Data. Genome Biol. 2018, 19, 213. [Google Scholar] [CrossRef]
- Lei, C.; Yang, J.; Hu, J.; Sun, X. On the Calculation of TCID50 for Quantitation of Virus Infectivity. Virol. Sin. 2021, 36, 141–144. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pålsson, S.A.; Sekar, V.; Kutter, C.; Friedländer, M.R.; Spetz, A.-L. Inhibition of Respiratory Syncytial Virus Infection by Small Non-Coding RNA Fragments. Int. J. Mol. Sci. 2022, 23, 5990. https://doi.org/10.3390/ijms23115990
Pålsson SA, Sekar V, Kutter C, Friedländer MR, Spetz A-L. Inhibition of Respiratory Syncytial Virus Infection by Small Non-Coding RNA Fragments. International Journal of Molecular Sciences. 2022; 23(11):5990. https://doi.org/10.3390/ijms23115990
Chicago/Turabian StylePålsson, Sandra Axberg, Vaishnovi Sekar, Claudia Kutter, Marc R. Friedländer, and Anna-Lena Spetz. 2022. "Inhibition of Respiratory Syncytial Virus Infection by Small Non-Coding RNA Fragments" International Journal of Molecular Sciences 23, no. 11: 5990. https://doi.org/10.3390/ijms23115990
APA StylePålsson, S. A., Sekar, V., Kutter, C., Friedländer, M. R., & Spetz, A.-L. (2022). Inhibition of Respiratory Syncytial Virus Infection by Small Non-Coding RNA Fragments. International Journal of Molecular Sciences, 23(11), 5990. https://doi.org/10.3390/ijms23115990