Abstract
The C57BL/6.NOD-Aec1Aec2 mouse has been extensively studied to define the underlying cellular and molecular basis for the onset and development of Sjögren’s syndrome (SS), a human systemic autoimmune disease characterized clinically as the loss of normal lacrimal and salivary gland functions leading respectively to dry eye and dry mouth pathologies. While an overwhelming majority of SS studies in both humans and rodent models have long focused primarily on pathophysiological events and the potential role of T lymphocytes in these events, recent studies in our murine models have indicated that marginal zone B (MZB) lymphocytes are critical for both development and onset of SS disease. Although migration and function of MZB cells are difficult to study in vivo and in vitro, we have carried out ex vivo investigations that use temporal global RNA transcriptomic analyses to track early cellular and molecular events in these exocrine glands of C57BL/6.NOD-Aec1Aec2 mice. In the present report, genome-wide transcriptome analyses of lacrimal glands indicate that genes and gene-sets temporally upregulated during early onset of disease define the Notch2/NF-kβ14 and Type1 interferon signal transduction pathways, as well as identify chemokines, especially Cxcl13, and Rho-GTPases, including DOCK molecules, in the cellular migration of immune cells to the lacrimal glands. We discuss how the current results compare with our recently published salivary gland data obtained from similar studies carried out in our C57BL/6.NOD-Aec1Aec2 mice, pointing out both similarities and differences in the etiopathogeneses underlying the autoimmune response within the two glands. Overall, this study uses the power of transcriptomic analyses to identify temporal molecular bioprocesses activated during the preclinical covert pathogenic stage(s) of SS disease and how these findings may impact future intervention therapies as the disease within the two exocrine glands may not be identical.
1. Introduction
Autoimmunity is generally recognized as a multi-step process initiated by environmental triggers that activate an innate inflammatory reaction which progresses to an adaptive immune response targeting a critical biological process in genetically or physiologically predisposed hosts whose dysfunction eventually results in an overt clinical pathology [1,2,3,4,5,6]. Autoimmune diseases, therefore, are considered to have both an early covert disease phase and a late overt disease phase. Unfortunately, patients most often present in clinic only after the adaptive immune phase is active and irreversible pathology has occurred. This scenario is especially true for Sjögren’s syndrome (SS), a highly debilitating, yet fascinating systemic autoimmune disease marked primarily by leukocytic infiltrations of the salivary and lacrimal glands with the concomitant loss of exocrine secretion leading to clinical symptoms of severe dry mouth and dry eye problems (reviewed in [7]). Furthermore, SS is a systemic autoimmune disease having a skewed prevalence towards women, potentially capable of involving multiple organ systems, exhibiting a wide variety of symptoms, and associated with lymphomagenesis [8]. An additional complication is the time between disease onset and a correct diagnosis which ranges from 4 to 10 or more years [7,9,10].
B cells and their cellular products, together with their ability to act as antigen-presenting cells and secrete auto-antibodies, are important factors in rheumatoid autoimmune diseases in general (reviewed in [11,12,13]), and Sjögren’s syndrome (SS) specifically [7,14,15,16]. Over the past couple decades, we have studied a number of rodent models, including NOD/ShiLtJ [17] C57BL/6.NOD-Aec1Aec2 [18], (C57BL/6.NOD-Aec1Aec2 X C57BL/6J)F1 recombinant inbred (RI) lines [19], and C57BL/6.Il14a transgenic (TG) [16,20] NOD-−/− [21], NOD-scid [22], NOD-Ifng−/− [23], NOD-IL4−/− [24], NOD-Stat6−/− [25], NOD-C3−/− [20,26], and NOD-Il17a−/− [27]. In those strains that develop a SS-like pathology, the disease appears spontaneously, develops temporally, and is characterized by: (a) aberrant proteolytic enzyme activity, (b) progressive loss of saliva and tear flow rates accompanied by increased protein content, (c) decline in amylase and carbonic anhydrase activities, and (d) appearance of autoantibodies, all manifestations dependent on the presence of B cells and occurring concomitantly with increasing glandular leukocyte infiltrates. Recent studies have also shown a direct correlation between an upregulated expression of IL14α and late-stage B cell lymphomagenesis [28,29] as well as the fact that full development of disease is a multi-phase process involving an innate inflammatory response, an adaptive response, and in some individuals, a late lymphomagenesis phase and/or organ involvement of kidneys and lungs. This progression of autoimmune pathologies in these models is significant when compared against sex- and aged-matched mice that do not develop SS-like disease despite developing occasional glandular infiltrations [30,31].
Studies in both BAFF [32,33,34] and B6.Il14α transgenic (TG) mice [28,29] have provided strong evidence that MZB cells are a critical cell population for onset and early disease phase development. Elimination of the MZB cell population or blocking the lymphotoxin activity required for MZB cell ontogeny [35] were shown to prevent development of SS-like disease, including lymphomagenesis. In support of these findings, our most recent genome-wide transcriptome studies of ex vivo salivary glands from the C57BL/6.NOD-Aec1Aec2 mice [36,37] have provided further evidence that MZB cells are recruited to the salivary glands during early-stage disease, thus apparently establishing an environment conducive for a subsequent destructive T-cell-mediated cytotoxic autoimmune attack. To follow up on these results, we have now carried out a similar transcriptome analysis using ex vivo lacrimal glands from our C57BL/6.NOD-Aec1Aec2 mice. The study has permitted a direct data comparison between the two glandular tissues as the tissues were derived from the same mice and the gene sets compared data from identical aptamers/probes. Results reveal overall similar, but not necessarily identical, profiles.
2. Results
2.1. Early and Temporal Transcriptome Expressions in Lacrimal Glands of C57BL/6.NOD-Aec1Aec2 Mice Identify MZB Cell Involvement in SS Pathology but with Potentially Unique BCRs
In support of the hypothesis that MZB cells are strongly involved in the early immune attack on the lacrimal glands of C57BL/6.NOD-Aec1Aec2 SS-susceptible (SSS) mice, our temporal global transcriptome data reveal a unique upregulated Notch2 signal transduction profile, as presented in Figure 1. Based on the specific set of upregulated genes concomitantly expressed, the cellular profile and signal transduction pathway defined is: Notch2 >Furin > Rfng > Jag1 > Adam10 > γSecretase (Psen1 and Ncstn) > Dtx4 > Hdac > Rbpk > Cbfat2 > Hes6, a pathway leading to Notch-specific gene transcription capable of activating multiple cell functions, including that of Nfkβ1 (nuclear factor kappa b subunit 1), Mapk14 (mitogen-activated protein kinase 14), and Infa1 (interferon type1). This early appearance also corresponds with the first observation of migration and infiltration of lymphocytes into the lacrimal glands noted histologically [38]. In addition, the transcriptome data reveal that MZB cells entering the lacrimal glands between 8 and 12 weeks of age are exhibiting both ontogenesis (increase receptor synthesis) and cell activation (increased Notch2 signal transduction). Furthermore, by 16 weeks of age, the transcriptome profile reveals a strong continued Notch2 profile, thereby suggesting a relatively prolonged activated state. While this overall profile corresponds with the onset of an inflammatory/innate response and initiation of early pathology within the lacrimal gland, respectively, it does not specify if this represents a single continuous cellular event or two time-dependent activities of multiple independent events. Interestingly, this lacrimal gland Notch2 profile differs slightly from the Notch2 gene profile seen in our recently published profile of salivary glands [36], specifically, (a) the lacrimal gland profile shows an upregulation of the Furin, Rfng, and Jag1 gene set in the Notch2 pathway, while the salivary gland profile shows an upregulated Furin, Mfng, Rfng, and Jag2 set of genes, and (b) the lacrimal gland profile indicates a strong upregulation of Deltex family gene Dtx4, while the salivary gland shows a strong upregulated Dtx1 gene instead. Nevertheless, since these two gene sets encode for MZB cell receptors and proteins that regulate Type 1 interferon (IFN1) production, these data raise the possibility that different MZB cell subpopulations are emigrating to the two different exocrine glands and recognizing different autoantigens.
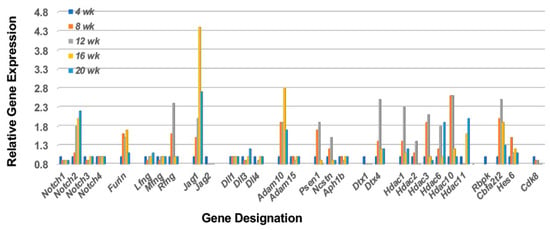
Figure 1.
Prolonged upregulated activation of the Notch2 signal transduction pathway in lacrimal glands prior to and during onset of the adaptive response. Notch2 is a transmembrane protein receptor whose extracellular portion is post-translationally modified in the Golgi bodies by furin and fringe molecules (i.e., Lfng, Mfng, or Rfng) prior to its insertion into the cellular membrane, thus enhancing subsequent MZB cell functions. On activation by a Notch2 ligand (i.e., Jagged and/or Delta molecules), the Notch2 molecule undergoes sequential cleavage first by Adam10 and then by γ-secretase, thereby releasing the cytoplasmic protein Dtx (an E3 ubiquitin ligase), as well as the Notch intracellular domain (NICD) that gets transported into the nucleus. In the nucleus, NICD displaces corepressors, including HDACs (histone deacetylase complex), allowing direct interaction with Rbpk to activate Hes, thus driving Notch2-regulated gene transcription. Downregulation of the Notch2 pathway occurs through phosphorylation of NCID by Cdk8 that initiates polyubiquitination and proteasome degradation. In this model, however, while the Notch2 pathway remains activated, Cdk8 exhibits, at best, minimal upregulated expression, suggesting the MZB cells may persist and over time differentiate from their effector function to their APC function for invoking the adaptive response.
A major function of the Notch2 pathway in stimulated MZB cells is to activate an IFN1 response, a critical activity for stimulating the interferon signature and subsequent SS disease. Regulation of Ifn1 synthesis is, in part, dependent on activation of Dtx4 (an E3 ubiquitin ligase) by Nlrp4. The Nlrp4 regulates Ifn1 synthesis by targeting Tbk1 (Tank-binding kinase 1) for degradation by Dtx4. Tbk1 degradation, therefore, is a critical component in down-regulation of Ifn1 in the activation pathway involving Tmem173 (Sting) > Tbk1 > Irf3 (Interferon regulatory factor 3). As shown in Figure 2, despite an upregulated Dtx4 expression, none of the possible Nlrp4 genes expressed in mice (i.e., Nlrp4a, b, c, e and f) exhibit temporal upregulated expressions, thereby permitting Tbk1 function to persist long-term. This profile, which is similarly seen in the salivary glands of C57BL/6.NOD-Aec1Aec2 mice [37], predicts a strong and prolonged Ifn1 response in the lacrimal glands of these SSS mice during the early disease state as well.
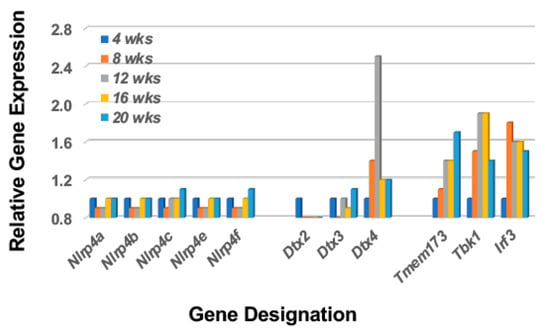
Figure 2.
A transcriptome profile that predicts activation of the type 1 IFN response despite upregulation of Dtx expression. The transcriptome profile of the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice indicate a lack of an upregulated Nlrp4 gene expression and thereby a concomitant targeting of Tbk1 by Dtx4. This profile would explain the strong upregulation of the signal transduction pathway for Type 1 interferon and activation of the interferon-signature characteristic of SS.
2.2. The Notch2 Signaling Pathway Predicts Activation of Mitogen-Activated Protein Kinase 14 (p38Mapk14α)
Although each Notch2 pathway component is essential in dictating specific MZB cell characteristics and functions that form the final immune profile, there are five interesting critical factors of regulation that help determine MZB cell differentiation and activation. These are Taok3 (Tao kinase 3), Adam10 (A disintegrin and metallopeptidase domain), Ncstn (Nicastrin), Btk (Bruton’s tyrosine kinase), and Ibtk (Inhibitor of Btk) [39,40,41,42]. While Taok3 is a factor expressed in virtually all tissues, one function is to commit T1 B cells to a MZB cell fate by mediating surface expression and activation of Adam10 via the BCR and Notch2 ligand pathways. Adam10 is expressed on mature MZB cells, but not on follicular B cells, and activates Notch2 downstream signal transductions. Nicastrin, a member of the γ-secretase complex, also plays an important role in the transitioning of T2 B cells towards MZB cells by glycosylating γ-secretase. In contrast, Btk blocks T2 B cell differentiation into MZB cells, instead promoting the differentiation of T2 B cells into follicular B cells. In contrast, Ibtk suppresses Btk function. As presented in Figure 3, the genes encoding Taok3, Adam10, and Nicastrin are each upregulated in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice starting as early as 8 weeks of age. At the same time, the gene encoding Btk remains quiescent, apparently suppressed by an upregulated Ibtk, thus further supporting the promotion of T2B cell differentiation into MZB cells. Interestingly, Taok3 gene expression reveals a different profile from Adam10 and Ncstn, but this overall profile is strongly reflective of MZB cell activation of p38Mapk14 (a dimeric protein composed of p38β p38α which, when coupled, can activate multiple systems) in the lacrimal glands even though demonstrating a relatively weak temporal upregulation.
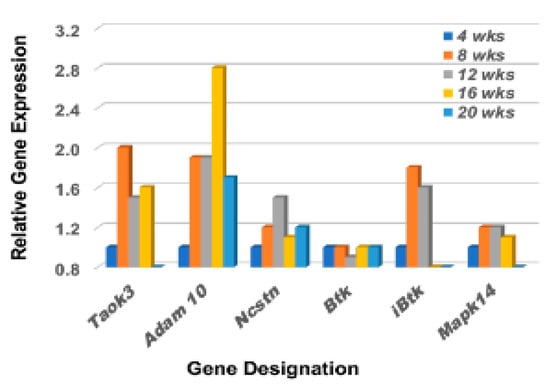
Figure 3.
Differential expression profiles for genes encoding various factors involved in the upregulation of MZB cell signal transductions. Comparative transcriptome data showing the temporal expression of genes encoding factors critical in regulating the Notch2 signal transduction pathway of MZB cells, i.e., Taok3 (TAO kinase 3), Adam10, Ncstn (Nicastrin), Btk (Bruton’s tyrosine kinase), iBtk (Inhibitor of Bruton’s tyrosine kinase), Prp38 (Proline rich protein 38), and Mapk14a (Mitogen-activated protein kinase 14a) active in lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice starting at 8 wks of age.
2.3. Limited Receptor Gene Expressions That Can Activate Early-Phase Immune Functions of MZB
In addition to B cell receptors (BCRs) and Toll-like receptors (TLRs), MZB cells also express Cr2 (the complement receptor encoded by Cd21), CD40 (the receptor for Cd40-ligand encoded by Cd40), IL22r (the interleukin 22 receptor encoded by Il22ra), S1pr (the sphingosine-1-phosphate receptor encoded by S1pr4), and TACI (encoded by Tnfrsf13b) [43,44,45,46,47]. Furthermore, MZB cells can be activated by either MZ macrophages expressing SIGN-R1 (encoded by Cd209a), Marco (macrophage receptor with collagenous structure encoded by Marco), and DCIR (encoded by Clec4a), or by metallophilic macrophages expressing Cd169 (encoded by Siglec1). As shown in Figure 4, of these genes, only Tnfsf13, Sign-R1 and Clec4a3 exhibit upregulated expressions in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice that start around 8 wks of age, with Marco exhibiting an upregulated expression at a later age. These data also suggest an absence of metallophilic macrophages, since there is no upregulated Cd209a expression. Interestingly, this lacrimal gland profile contrasts with the profile reported for this gene set in salivary glands [36] where genes encoding for Cd21, S1pr4, Tnfrsf13b (Baff), Tnfsf13 (April), Marco, Cd209b, and Clec4a4 are each upregulated simultaneously at 16 weeks of age. Unexpectedly, neither Cd40 nor Cd40lg are upregulated in the lacrimal gland, thus mimicking what we reported in the salivary glands of C57BL/6.NOD-Aec1Aec2 mice. These data further support the concept that the B cell population in the exocrine glands are MZB cells and not B2 cells.
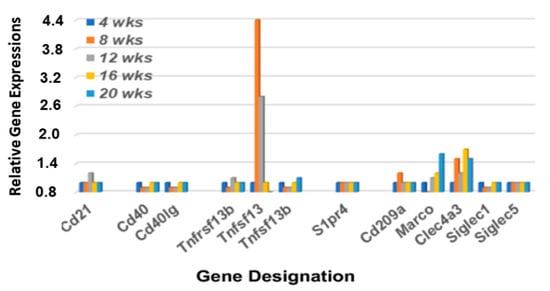
Figure 4.
Differential expression profiles for genes encoding various receptors associated with MZB cell functions. Transcriptome data showing the temporal expression of MZB cell-associated receptor genes (Cd21, Cd40, S1pr4, and Tnfrsf13b), receptor ligands (Cd40lg, Tnfsf13, and Tnfsf13b) and macrophage receptors (Cd209b, Marco, Clec4a3 and Siglec1) in lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice.
2.4. Receptor-Ligand Dependent Emigration of Immune Cells to the Lacrimal Glands
An interesting phenomenon that distinguishes MZB cells from follicular B cells, particularly in mice, is the relatively non-circulatory state of MZB cells compared to the more robust migratory activity of follicular B cells in the normal state. As proposed by Lu and Cyster [48], this difference occurs because of the upregulated surface levels of integrin expressions on MZB cells, especially LFA-1. LFA-1 is a heterodimeric integrin, comprised of ItgαL and Itgβ2 subunits that, in a normal state, binds to ICAM-1 expressed in marginal zones (MZs). Development of splenic MZ areas during ontogeny requires lymphotoxin-dependent integrin-mediated induction of ICAM-1 and VCAM-1, with ICAM-1 being the primary receptor for LFA-1. High levels of integrin expression on MZB cells raises the threshold levels of chemokines required to dissociate MZB cells from splenic MZs for emigration to other tissue sites. As presented in Figure 5, our transcriptomic data reveal that genes encoding LFA-1 integrin subunit Itgal is minimally upregulated while integrin subunit Itgb2 is highly upregulated, starting around 12 weeks of age. At the same time, expression of Icam1 and Vcam1 show opposing upregulated profiles from 8 to 20 wks of age, indicating development of an environment in the lacrimal glands suitable for capturing MZB cells during this time frame.
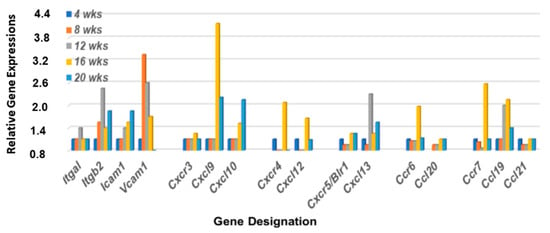
Figure 5.
Differential expression profiles for genes encoding integrin, chemokine, and chemokine receptors critical for MZB cell emigration from the MZ regions. Transcriptomic data reveal that genes encoding LFA-1 subunits (Itga1 and Itgβ2), together with Cxcl13, show a coordinated upregulated expression at 12 wks of age in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice. Chemokine receptors Cxcr4 and Ccr7, together with their ligands Cxcl12 and Ccl19, show strong upregulations at 16 wks of age, while chemokine receptor Cxcr3 and chemokine ligands Ccl20 and Ccl21 exhibit no upregulated expressions.
MZB cells also express multiple chemokine receptors, especially Cxcr3, Cxcr4, Ccr5, Ccr6, and Ccr7. Our previously published data [36,37] have shown that genes encoding these five chemokine receptors are highly upregulated in the salivary glands of C57BL/6.NOD-Aec1Aec2 mice at 4 months of age, along with the genes encoding chemokine ligands Cxcl9, Cxcl10, Cxcl12, Cxcl13, Ccl19, and Ccl21. In contrast, chemokine receptor genes Ccr1, Ccr2, Ccr3, Ccr8, Ccr9, and Ccr10 showed no upregulation in the salivary glands, suggesting limited or no involvement of neutrophils up to 20 wks of age. As presented in Figure 5, the gene expression profile for these chemokines in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice exhibit a different transcriptome profile with chemokine receptor gene Cxcr4, Ccr6, and Ccr7 showing a strong upregulation, Cxcr3 and Ccr5 a weak upregulation, and the chemokine ligands Cxcl9, Cxcl10, Cxcl12, and Ccl19 a substantial upregulation.
2.5. Identification of Cell Emigration to the Lacrimal Glands in C57BL/6.NOD-Aec1Aec2 Mice
The temporal gene expression profile presented above strongly suggests an emigration of MZB cells in the earliest stages of SS development in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice. In addition, the upregulation of the Notch2 signal transduction pathway suggests this specific B cell population is undergoing both ontogenesis and cell activation followed by transcription supporting a relatively prolonged functional state. This profile also corresponds temporally with the onset of the inflammatory/innate response and initiation of the adaptive response within the lacrimal glands observed histologically [38]. One of the main manifestations of SS disease is the progressive periductal leukocytic infiltrations of the lacrimal and salivary glands, but while this emigration and expanding leukocytic infiltration is considered an important pathological event, its molecular basis remains poorly defined.
Cell migration is highly dependent on at least three bio-entities: chemokine signaling, focal adhesion assembly, and Rho-GTP family protein activations. Although we have presented a chemokine > chemokine receptor profile above and have previously published the molecular composition of the focal adhesion(s) activated in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice [49], no profiling of Rho-GTP gene expressions in this SSS model have yet been performed. In a quiescent state, Rho-GTP proteins mostly localize to cellular membranes but dissociate from these membranes during initiation of migration. In mice, the Rho-GTP family consists of 21 members subdivided into several subfamilies: 11 Rho proteins, 3 Rnd proteins, 3 Rac proteins, 3 Rhobtb proteins and Cdc42 [50]. Interestingly, the profile of these molecules in the lacrimal glands indicates that seven Rho genes (a, b, c, g, j, q, and u), two Rac genes (1 and 2), one Rhobtb gene (3), and the Cdc42 gene are upregulated, and in each case, upregulated at 8 to 12 weeks of age (Figure 6).
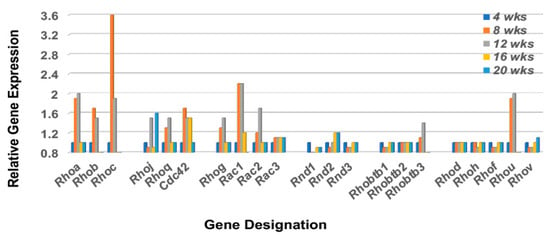
Figure 6.
Transcriptomic profile of the Rho-GTP family of proteins in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice. Rho-GTPase family proteins are important in regulating the cellular homeostasis of the GTP<>GDP system. The Rho-GTP family consists of 21 members of which 16 are divided among 5 subfamilies (Rsf1, Rsf2, Rsf3, Rsf4 and Rsf5) with 5 unassigned (designated Rsfx). For cells in the non-activated state, Rho-GTP molecules are mostly associated with cellular membranes, but during cell activation the molecules dissociate to the cytoplasm by an as yet unknown mechanism.
The Rho-GTP proteins are further divided into two functional classes, those that are incapable of hydrolyzing GTP (i.e., Rhoh and Rnd subfamily members that constitutively bind GTP), and those that hydrolyze GTP through an opposing equilibrium of Rho-specific guanine nucleotide exchange factors (GEFs) and GTPase-activating proteins (GAPs) [50,51]. GEFs activate Rho-GTPases by exchanging a bound GDP with a GTP, while GAPs inactivate Rho GTPases vis-a-vis GTP by catalysis. Temporal gene expression profiles for GAP and GEF molecules in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice are summarized in Figure 7 left and right, respectively. These gene expressions again reveal a restricted activation with ten of the 23 Arhgap gene family members (1, 5, 6, 9, 11a, 12, 17, 18, 21, and 29) and eight of the 17 Arhgef gene family members (1, 3, 5, 6, 7, 12, 16, and 18) upregulated. In addition to the Arhgef protein family, a second subset of Rho-GEF proteins are the eleven DOCK (Dedicator of cytokinesis) family proteins [52,53,54]. DOCK molecules, also known as CZH proteins, possess functional CZH2 domains that promote the exchange of GDP to GTP in the formation of Rho-GTPases. Based on sequence homologies represented within the eleven identified DOCK molecules, this molecular family of proteins is currently divided into four major subgroups, (A) Dock1, 2 and 5, (B) Dock3 and 4, (C) Dock6, 7, and 8, and (D) Dock9, 10 and 11. Besides playing an important role in multiple cellular functions, an additional critical function is cell migration, especially Dock2, 9, 10, and 11. As presented in Figure 8, the temporal gene expression profiles of the Dock molecules in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice is consistent with this concept, as highest upregulated temporal expression activity is in Dock 2, 9, 10, and 11 occurring concomitantly with development and onset of SS in this mouse model. The upregulation of Dock1 is also consistent with glandular structural changes taking place at 8–12 weeks of age [55,56]. Also of interest is the fact that development and accumulation of MZB cells in MZs, followed by their subsequent migrations, appear to require the Dock2 > Rho-GEF > Pyk2 molecular pathway [50,53]. Furthermore, the recently published work from Gotoh et al. [54] has shown that Dock2 is indispensable for the co-migration of plasmacytoid dendritic cells (pDC), a highly relevant fact considering the extensive type 1 IFN-signature seen in exocrine glands of both human and mouse SS diseases.
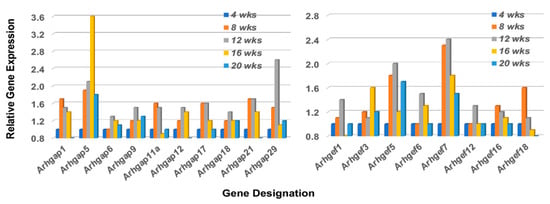
Figure 7.
Temporal gene expressions for the Rho-GTP Arhgap and Arhgef gene family members upregulated in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice during onset of SS. Transcriptome data indicating the temporal expressions of the 8 of 23 Arhgap genes (left), and 8 of 17 Arhgef genes (right) found to be upregulated in the lacrimal glands.
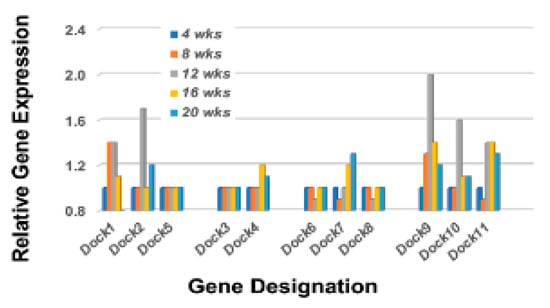
Figure 8.
Temporal gene expressions for the Rho-GTP Dock gene family members. Transcriptome data showing the temporal expressions of the Dock genes upregulated in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice. Two genes, Dock1 and Dock9, exhibit an upregulated expression starting at 8 wks of age, while Dock2, Dock10, and Dock11 exhibit an upregulated expression starting at 12 wks of age.
Lastly, Rho-GTPases are known to interact with numerous proteins that subsequently form active signal transduction pathways capable of regulating multiple activations of bioprocesses critical to cellular functions, especially migrations. These include members of several protein families including Cdc42 interactive proteins, Vav proteins, Bai1-associated proteins, Stat proteins, Forkhead box factors, and various RhoGTPase signaling proteins. A transcriptome profile of these interactive molecules from the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice is shown in Figure 9. While most of the genes listed also have roles in multiple signal transduction pathways and cellular functions, an interesting profile is revealed when considering those genes upregulated 8 to 12 weeks of age, in-line with the temporal profiles of upregulated Rho-GTPases in the lacrimal glands presented above. These include Cdc42, Cdc42bpb, Cdc42bpg, Cdc42ep3, Cdc42ep5, Cdc42se1, Cdc42se2, Vav2, Tiam1, Setd3, Setd5, Setd7, Grb2, Flii, Stat6, Stat1, Crk1, Crk7, Grb14, Creb3, Creb3l1, Foxo3a, Baiap2, Baiap2l1, Pak1, and Scrib. Moreover, the genes Stat6 and Stat1 show temporal upregulated expression profiles consistent with their known functional activities in SS, i.e., Stat6 being critical during early stages of SS and Stat1 being important in later stages of disease, while Pard3 exhibits a later upregulated expression at 20 weeks.
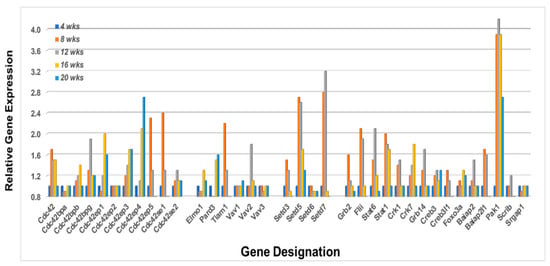
Figure 9.
Temporal gene expressions for various proteins important in signal transduction pathways involving the Rho-GTPases. Transcriptome profiles for multiple family genes (e.g., Cdc42, Vav, Setd, Crkb, Stat, Crk, and Baiap)), plus individual genes (e.g., Tiam1, Grb2, Flii, Creb3, Foxo3a and Pak1) in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice. Examination of the temporal differential gene expressions, it appears that the heightened transcriptome profile at 8 week is Cd42 > Cdc42ep5 > Cdc42se1 > Tiam1 > Setd3 > Setd7 > Grb2 > Flii > Stat6 > Stat1 > Crk1 > Greb3> Creb3l1 > Baiap2 > Pak1, while at 16 weeks it is Cdc42 > Cdc42bpb > Cdc42ep1 > Cdc42ep4 > Elmo1 > Pard3> Crk7 > Foxo3a > Pak1.
3. Discussion
More than two decades ago, studies by Robinson et al. [57] using the NOD-Igµ−/− gene knockout (KO) mouse model revealed an absolute requirement for B cells and immunoglobulin in the development and onset of SS-like disease that normally develops spontaneously in parental NOD/ShiLtJ autoimmune mice. Studies in both BAFF [32,33,34] and B6.Il14α transgenic (TG) mice [28,29] not only support this earlier finding, but have further shown that elimination of the MZB cell population or blocking the lymphotoxin activity required for MZB cell ontogeny [35] prevents development of SS-like diseases, including lymphomagenesis. MZB cells are an unique subpopulation of bone marrow-derived B lymphocytes characterized by limited expression of immunoglobulin variable region genes that produce predominantly IgM antibodies, many of which are self-reactive [46]. In addition, they are strategically located within mucosal surfaces, function as innate and/or transitional cells capable of rapid responses to both T cell-independent and T-cell dependent antigens, and regulate the activation of subsequent adaptive immune responses by T and B2 lymphocytes in association with monocytic and neutrophilic antigen-presenting cells (APCs) (reviewed in [46]). Functionally, MZB cells differentiate from transitional type-1 (T1) B cells under the influence of low affinity B cell receptor (BCR) signals and transcription factors such as Notch-2, RBP-J and MAML1 [40]. They are enriched within splenic marginal zones (MZs), retained there by interactions between integrin and integrin receptors (e.g., LFA1, ICAM, VLA-4, and SIP1 [48] and respond to Toll-like receptor signaling, especially TLR2, 4 and 9 [15,48,58]. Despite their ability to respond to self-antigens, MZB cells reportedly are more adept at responding to pathogens due to an apparent weak signaling through their BCRs [53,59]. Importantly, MZB cells have been identified within the salivary glands of patients with salivary gland disease, and these infiltrating cells secrete cytokines cytotoxic to salivary gland cells [14,60,61]. Despite these observations and recognized extensive cellular attributes, MZB cells have generally been ignored in studies of SS in both humans and mouse models, in part because MZB cells are a difficult cell population to isolate functionally and then manipulate. However, using temporal global transcriptomic microarrays, we are now able to follow MZB cell markers to identify their temporal presence and potential molecular bioprocesses involved in autoimmune diseases.
Over the past two decades, we have studied multiple murine models of Sjögren’s syndrome to identify the molecular and cellular factors involved in development and onset of those pathophysiological manifestations associated with destruction and loss of lacrimal and salivary gland tissue and subsequent secretory functions. Although it is clear that a T cell-mediated autoimmune process is involved in glandular destruction and onset of clinical disease, actual initiation of SS appears to be fully dependent on B cells [16,57], specifically the MZB cell population per se. This finding raises the question of which molecular bioprocesses activated in MZB cells are relevant to the early underlying stage(s) of SS autoimmunity. The current study addresses two important issues in this regard: (1) is there evidence that MZB cells are involved in the initial immune response(s) that result ultimately in SS disease of the eye, and (2) is the early-stage gene expression profile of lacrimal glands in SSS C57BL/6.NOD-Aec1Aec2 mice identical to our previously published data for salivary glands [36,37]. Results could clearly impact how future intervention therapies may need to be designed.
MZB cells possess the capability for mounting immune responses to both T-dependent and T-independent antigens, as well as for regulating both innate and adaptive immunity [62]. This capacity results from an inherent interaction between polyreactive BCRs, TLRs, S1P, chemokine receptors, and complement receptors with integrin LFA1 [48,63,64,65]. High LFA1 expression in splenic MZs prolongs retention of MZB cells and reduces their circulation through the blood and lymph systems This retention permits the BCRs of MZB cells direct access to blood-borne antigens presented by dendritic cells (DCs) and granulocytes (GCs). In addition, complement-decorated antigens bind to complement receptors, while an array of antigens can also bind to TLR and S1P receptors. In turn, antigen-activated MZB cells possess the capability to present antigens to T lymphocytes or, with costimulatory signals, differentiate into IgM-secreting plasmablasts that can undergo class switching in the correct environment. Lastly, MZB cells express several chemokine receptors that direct immune cell migrations along chemokine ligand gradients. An important chemokine receptor is Cxcr5 whose ligand is chemokine Cxcl13. At high concentrations, Cxcl13 dislodges MZB cells from MZs, permitting their migration to sites of injury [66]. Transcriptome data presented here indicate a marked upregulation of several chemokine receptor and chemokine ligand, especially Cxcr5 and Cxcl13, proximate with the appearance of infiltrating lymphocytes in the lacrimal glands [38].
MZB cell activation against T cell-independent antigens generally occurs in MZs through presentation of antigens to their TLRs via macrophages, or to their BCRs via reticular cells, neutrophils, and/or dendritic cells. Neutrophils and dendritic cells release both BAFF and APRIL (which in turn activates TACI on MZB cells), while neutrophils also secrete IL-21 which is involved in class-switch recombination and somatic hypermutation that can lead to IgA and IgG synthesis by both MZB cells and T follicular helper cells of germinal centers [67,68,69,70]. Dendritic cells also release chemokine ligands Cxcl9, Cxcl10, and Cxcl11 which bind to chemokine receptor Cxcr3. However, no evidence was seen for upregulated Il21 or Cxcr3 gene expressions out to 20 weeks of age in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice, possibly suggesting an absence of B cell class switching per se within the lacrimal gland itself. In contrast, Cxcr4 and its ligand Cxcl2, as well as Ccr7 and its ligands Ccl19 and Ccl21, are each upregulated, which may point to the initial recruitment, appearance, and activation of the T cell-mediated response.
In contrast to murine MZB cell responses against T cell-independent antigens, murine MZB cells responding towards T cell-dependent antigens can utilize several uniquely different molecular processes. First, Dcir2+ MZ dendritic cells capture and present antigens to both MZB cells and CD4+ T cells. This event promotes the T cell population to differentiate into Th2 cells that secrete Cd40 ligand and IL4, providing an environment for reactive MZB cells to transition to an antigen-presenting cell and differentiate to IgG1-producing plasmablasts. However, neither the Cd40 nor Cd40 ligand gene exhibited upregulated expression within the lacrimal glands. Second, MZB cells that are lipid-reactive can present various glycolipids via Cd1d molecules to invariant natural killer T (iNKT) cells which, with a concomitant production of IL4-, Cd40 ligand-, and Ifnγ-activated iNKT cells, can induce differentiation of the MZB cells to IgM/IgG plasmablasts. Third, MZB cells that capture complement-opsonized antigens downregulate their S1P receptors, which enhances Ccr5+ MZB cell recruitment by chemokine Cxcl13 to splenic and/or nodal follicles where they provide captured antigen presentation to follicular DCs for activation of follicular T and B cells. While migration of MZB cells is reportedly dependent on LSC (encoded by Arhgef1), more recent studies have indicated that Arhgef6 can perform this function as well [71]. Our transcriptomic profiling of lacrimal glands indicates that both Arhgef1 and Arhgef6 genes are upregulated, different from salivary glands where only Arhgef6 is upregulated [37], and this upregulation occurred with highest gene expressions at 12 weeks of age.
Although the microvasculature of mouse and human splenic marginal zones (MZs) are anatomically different, with human spleens apparently lacking a marginal sinus influencing migratory pathways, both species provide environments for MZB cells to interact with APCs in their respective stromal reticular cell networks [72], thereby permitting efficient immune surveillance of blood-borne antigens. The precise cellular functions attributed to MZB cells in autoimmunity and SS clearly require further definition, yet both species separate MZB and follicular B cells via the periarteriolar lymphoid sheaths (PALS). Furthermore, both mouse and human MZB cells tend to utilize similar emigration and activation signaling. For the present study, an overriding question is whether MZB cells function as the inducers of innate autoimmunity [73,74,75]. This requires a correct stromal cell environment within MZ areas and, most likely, the immune-targeted autoantigens to which an extra-nodal lymphogenesis occurs. Ontological development of MZs requires induced expression of ICAM1 (intercellular adhesion molecule-1), VCAM1 (vascular cell adhesion-1), and MADCAM1 (mucosal addressin cell adhesion molecule-1) that, in the presence of IL-7, recruits lymphoid tissue inducer cells (LTIs) that express RXRγt to sites surrounding blood vessels [76]. LTIs secrete lymphotoxin that stimulates secretion of chemokines, especially CCL19 and CCL21, that attract dendritic and lymphoid cells, plus regulate endothelial cells delineating marginal sinuses [77]. Three other stromal cell populations that dictate ontological effects on MZs are the marginal reticular cells (MRCs) which express RANKL (receptor activator of NF-kβ ligand), fibroblast reticular cells (FRCs) which along with MRCs regulate immune responses via production of reticular fibers and chemokines that guide lymphocyte trafficking, plus follicular dendritic cells (FDCs) which secrete CXCL13 and BAFF to enhance B cell maturation and survival [78]. Importantly, these stromal cell populations secrete type 1 interferon(s) in response to B cell secretion of lymphotoxin, promoting rise of IFN-signatures for autoimmune diseases such as rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), and Sjögren’s syndrome (SS) [6,79]. The current study now provides a possible molecular basis for the prolonged IFN1 production since our study reveals a failure in suppression of Tbk1 gene expression via the Nlrp4 > Dtx4 pathway thereby prolonging the IFN signal transduction pathway [80].
As stated above, MZB cells are activated through BCR and TLR antigen binding plus TACI binding of BAFF and/or APRIL. While TACI interacts with TLRs to stimulate the MYD88 > TRAF signal transduction pathway that subsequently activates Nf-kβ, it can also interact with BCRs to activate class switch recombination (CSR) in the presence of CD40 to initiate rapid innate antibody responses [81]. In this regard, murine MZB cells possess strong antigen-presentation capability for CD4+ T cells due in part to elevated expressions on MZB cells of MHC class II, CD80 and CD86, while lipid-activated MZB cells activate invariant natural killer T (iNKT) cells [82,83]. Thus, while MZB cells are strong producers of IgM antibodies, under the correct environment, MZB plasmablasts per se can be highly capable of producing both IgA and IgG antibodies that enhance targeted phagocytosis and killing [84]. Class switch recombination in MZB cells also appears to be regulated by a distinct neutrophil population present in the peri-MZs and characterized by high productions of BAFF, APRIL, CD40L, IL-6, IL21, and chemokines, especially CXCL12 [46]. Although these factors have been reported to be upregulated in various SSS mouse models, the Il21, Cd40, and Cd40l genes were not found to be upregulated in the lacrimal glands of the C57BL/6.NOD-Aec1Aec2 mice. Overall, these data appear to explain the observations that individuals with neutropenia, defective TACI and/or an impaired STAT3 have reduced MZB cells and exhibit reduced IgA and IgG antibody production toward T-dependent antigens.
Results of the present transcriptomic analysis also reveal distinct temporal gene profiles for Rho-GTPase family and subfamily proteins in the lacrimal glands of our SSS-susceptible mice. Rho-GTPases are known to play critical roles in multiple biological processes, especially cell migrations. While our previously published genome analyses [85] identified the Rac > Ras > Raf > Erk > AP pathway in development of glandular pathology, the current in-depth analysis of the Rho-GTPases suggests a more complex biological process involving multiple Rho-GTPase and Rho-GTPase-associated gene sets. For example, Tedford et al. [86] reported that Vav1/Vav2-negative B cells are unresponsive to thymus-independent antigens in vivo and indicated a role for Vav-2 in BCR calcium signaling critical to B cell development and function. Similarly, Wang et al. [87] have reported that membrane-proximal BCR signaling molecules (including Vav3, Lyn, Syk, Btk, PLC-γ2, and Blnk) together with adaptor molecules Grb2, Cbl, and Dok3 link BCR micro-clusters and motor proteins. Our transcriptome data indicate that Tiam1 and Vav2, but neither Vav1 nor Vav3 are temporally upregulated in the lacrimal glands of C57BL/6.NOD-Aec1Aec2 mice, while Pard3, known for its ability to interact with Tiam to suppress Tiam1-Vav2-Rac1complex function [88], is upregulated concomitantly with the downregulation of Tiam1 and Vav2, thereby establishing a profile suggesting an interesting activation versus regulation of cell differentiation, polarity, division and emigration.
Lastly, a special set of Rho-GEF proteins is the subfamily of DOCK molecules. A direct correlation exists between upregulated DOCK subfamily molecules Dock2, 8, 10, and 11 with cellular homeostasis, development, and migration [89,90,91]. Interactions of DOCK molecules with the Rho-family CDC42 proteins are also known to activate conformational changes in p21-activated PAK family molecules that regulate actin reorganization critical to cell adhesion and invasiveness, as well as upregulation of RAF and RAS pathways leading to activations of ERK, NF-kB, and AP1 (c-Jun, c-Fos and Atf2). As first reported by Namekata et al. [51], expressions of DOCK subgroup D molecules (i.e., Dock9, 10, and 11) are primarily in peripheral blood lymphocytes. However, more recently, evidence suggests that Dock10 expression is uniquely upregulated in B cells by IL-4 and that Dock10.1 isoform regulates T cell activities, while Dock10.2 isoform regulates CD23 expression on B cells, sustains B cell lymphopoiesis in secondary tissues, and up-regulates the IL4 > Stat6 pathway [89]. Our previously published studies have shown that IL4 and Stat6 gene knockout SSS mice prevents SS disease development [24,25]. In addition, Dock2-Pi3kdelta activation regulates B cell migration and proliferation, while Dock2-Pi3kgamma activation regulates T cell migration and proliferation [90,91]. While the importance of Dock10 cannot be ignored in B cell activation, Dock2 gene knockout mice are deficient in MZB cells [90], as are mice with deficiencies in Rac2, Pyk2 or Rho-GEFs. Nevertheless, both B and T cell emigrations into the exocrine glands of C57BL/6.NOD-Aec1Aec2 mice occur concomitantly with the upregulation of Dock2 and Dock10.
In summary, data presented from this temporal global transcriptome analysis provide new insights into the role of B lymphocytes, specifically MZB cells, play in development of SS autoimmunity in the lacrimal glands. It identifies probable molecular mechanisms and bioprocesses involving MZB cell emigration to the lacrimal glands during the covert phase of disease development, and the genetic basis for several observed pathophysiologic characteristics of the disease previously identified histologically [38,92,93]. The transcriptome profiles support the presence of a MZB cell population in lacrimal glands of our SSS C57BL/6.NOD-Aec1Aec2 mouse model beginning as early as 8 weeks of age, a time when autoantibodies, e.g., anti-nuclear autoantibodies (ANA) and anti-muscarinic receptor (M3R) antibodies, are first detected in sera and small peri-ductal lymphocytic aggregates are seen in the lacrimal glands [38]. A Notch2 signal transduction profile in the lacrimal glands suggests a MZB cell population that is undergoing both ontogenesis and activation, with the latter persisting through the early disease stage. Considering the importance of MZB cells in rapid immune responses to disease, it is not surprising to see a transcriptomic profile defining MZB cells and one that changes temporally. In conjunction with our previously published studies indicating absolute requirements for Ig BCRs [57], C’3 [26], IL4 [24], STAT6 [25], TH17 cells [27], and IFN [23] in the development of an overt clinical disease, we propose that the transition to a T cell-mediated cytotoxic response most likely occurs in the follicular areas of the spleen and lymph nodes induced by MZB cells emigrating from the exocrine glands back to secondary lymphoid tissues. These 8- and 16-week timepoints, therefore, represent potential focal points for intervention therapy if diagnosed in time. Nevertheless, it is important to note that our transcriptome analyses strictly reflect the ongoing bioprocesses in the lacrimal glands and fails to address any active bioprocesses occurring in the follicles of the draining lymph nodes or spleen [94]. Nevertheless, the current study shows how the gene profile described here for lacrimal glands mimics closely our published gene profile for salivary glands, at the same time reveals important differences. Furthermore, the study also demonstrates the power of temporal transcriptomic analyses to establish a foundation for further in-depth investigations that are expected to identify uniquely novel targets for possible testing of intervention therapies.
4. Materials and Methods
4.1. Animal Model
C57BL/6.NOD-Aec1Aec2 mice, a diabetes-free model of primary SS-like disease, were bred and maintained under specific pathogen free (SPF) conditions within the College of Medicine’s Department of Pathology’s Mouse Facility with oversight by Animal Care Services at the University of Florida, Gainesville. The C57BL/6.NOD-Aec1Aec2 mouse is a well-studied and well-characterized model of SS that spontaneously develops features of SS observed in patients. Mice were maintained on a 12-hr light–dark schedule and provided food and acidified water ad libitum. Any mice showing signs of eye dryness were treated daily with salve ointment. For the present study, breeding pairs (one male and one female) were paired per cage and offspring were weaned at 3 and 5 weeks of age. Weaned mice were caged maximum of n = 5 per cage per sex. The experimental mice were raised to a maximum of 20 weeks of age, a time prior to signs of any overt or covert disease. Euthanization was carried out at the appropriate timepoints by cervical dislocation after deep anesthetization as stipulated by the Panel on Euthanasia of the American Veterinary Association. Lacrimal and salivary glands were explanted simultaneously from the same animals. There were no indications that this procedure affected subsequent preparation of RNA specimens. Both the breeding and use of these animals for the present studies were approved by the University of Florida’s Institutional Animal Care and Use Committee (IACUC) under protocols 2008011756 and 201004828.
4.2. RNA Preparations
Procedures for the isolation, preparation, and quality testing of RNA samples are described in detail elsewhere [92,93]. In brief, extra-orbital lacrimal glands, free of lymph nodes, were excised in parallel from male C57BL/6.NOD-Aec1Aec2 mice euthanized at 4-, 8-, 12-, 16-, or 20 weeks of age (n = 4–6 per age group), snap-frozen in liquid nitrogen and stored at −80 ˚C until all samples were obtained. Using one lacrimal gland from each mouse, total RNA from each specimen was isolated concurrently using the RNeasy Mini-Kit (Qiagen, Valencia, CA, USA), as per the manufacturer’s recommended protocol. Each RNA sample was hybridized on an Affymetrix 3′ Expression Array GeneChip Mouse Genome 430 2.0 array and annotated (build 32; 06.09.2011). Each GeneChip contained 45,102 DNA sequences, thus the resulting data set consisted of 1,127,550 data points. A heatmap of differentially expressed genes is published elsewhere [92]. To verify relative microarray gene expressions, numerous genes were randomly selected for relative comparative expression by real-time polymerase chain reaction (rt-PCR) analyses (see [92]. The rt-PCR data confirmed both differential and temporal gene expressions. Full microarray data libraries are deposited with Gene Expression Omnibus, Accession number GSE15640.
4.3. Microarray Data Analyses
Gene expression analyses have been detailed elsewhere [92,93]. In brief, microarray data were normalized using the robust multiarray average (GCRMA) algorithm, followed by Linear Models for Microarray Analysis (LIMMA) (http://www.r-project.org) (accseesed on 6 April 2022) for differential expression determinations. The fdr (false discovery rate) method of Benjamini and Hochberg [95] was used to adjust the p-values for multiple testing. The original data represent 5 equally spaced timepoints; thus, multiple models were used to identify temporal patterns of gene expression, i.e., the linear fit (degree = 1), quadratic fit (degree = 2), cubic fit (degree = 3), and quartic fit (degree = 4) regression models. At this point, each gene has a single statistical value based on n = 5 data points at each of the 5 timepoints. B-statistics were calculated for each gene providing odds that a gene shows either positive or negative trends over time. In addition, these temporal changes in an individual gene’s expression relative to its value at the 4 weeks timepoint were considered; thus, data presented show the differential expression of an individual gene’s value at the 8, 12, 16 or 20 weeks timepoints relative to that gene’s value at the 4 weeks timepoint set as 1.0. The 4 weeks’ time point is a time when the exocrine glands have reached functional maturity and considered in a pre-diseased state. In the current study, the gene selection is not based on a SS-non-susceptible (SSNS) mouse strain, but on data generated in similar studies carried out using the salivary glands from the C57BL/6.NOD-Aec1Aec2 SS-susceptible (SSS) mouse strain; thus, our previously published gene expression data from the salivary glands represent the comparative data [69].
5. Conclusions: Summary and Contribution to the Field
Sjögren’s syndrome (SS), a human systemic rheumatoid autoimmune disease affecting lacrimal and salivary gland functions that lead respectively to dry eye and dry mouth pathologies, is considered an under-studied disease appearing most frequently in middle-aged women. Patients generally present in clinic only after the adaptive autoimmune phase is active and irreversible pathology has occurred. Furthermore, the time between disease onset and a correct diagnosis can range from 4 to 10 or more years, permitting the pathology to progress unchecked without intervention therapy, plus seriously slowing generation of effective therapy development. To address these issues, numerous mouse models exhibiting SS-like disease have been developed that are useful in identification of pre-disease phases of the autoimmune processes. While studies in these rodent models have addressed multiple cellular and pathological events, many details of the underlying cellular and molecular events remain unidentified. One observation that has recently come to light is an early-stage appearance of MZB cells within the salivary glands occurring concomitantly with the onset of SS pathology. Here, we provide evidence that indicates MZB cells are also infiltrating the lacrimal glands of the C57BL/6.NON-Aec1Aec2 mouse model early in the development of its SS-like disease. Furthermore, we have begun to define molecular processes involved with the cellular emigration to the lacrimal gland, specifically temporal activations of genes representing chemokines, focal adhesions, and Rho-GTPases. Comparisons of the present lacrimal gland profiles with those previously published for the salivary gland [36,37] indicate both similarities and differences in molecular processes occurring in the two glands. In summary, using temporal global transcription analyses, we have begun identifying molecular profiles for those critical bioprocesses involved in SS development and onset; however, the different profiles emerging for salivary and lacrimal glands are suggesting potential autoimmune processes that could seriously impact how one approaches design and development of potential therapies.
Author Contributions
Conceptualization, writing and editing (A.B.P., C.Q.N. and J.L.A.J.); Methodology, validation, data analyses, and curation (A.B.P. and C.Q.N.). The authors have no conflicts of interest with regards to this publication. All authors have read and agreed to the published version of the manuscript.
Funding
Creation of the microarray data was supported in part by the National Institute of Dental and Craniofacial Research (NIDCR) grant R01 DE-014344 and the National Institute of Allergy and Infectious Diseases (NIAID) grant AI-0819529. Publication of the manuscript supported by author’s personal funds.
Data Availability Statement
Gene Expression Omnibus, Accession number GSE15640.
Conflicts of Interest
The authors declare no conflict of interest. In addition, the NIH had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| APC | Antigen-presenting Cell |
| APRIL | Tnfsf13 (Tumor necrosis factor superfamily, member 13 |
| ARHGAP | Rho GTPase activating protein |
| ARHGEF | Rho GTPase exchange factor |
| BAFF | Tnfsf13b |
| BCR | B cell receptor |
| B220 | CD45-receptor |
| CCR | Chemokine (C-C motif) receptor |
| CCL | Chemokine (C-C motif) ligand |
| CD | Cell differentiation antigen |
| CDC42 | Cell division cycle 42 |
| CXCL | Chemokine (C-X-C motif) ligand |
| CXCR | Chemokine (C-X-C motif) receptor |
| DC | Dendritic cell |
| DOCK | Dedicator of cytokinesis |
| FACS | Fluoresec-activated cell sorting |
| GC | Granulocyte |
| GCRMA | Guanine-cytosine robust multi-array average |
| GTP-GAP | G protein bound-guanine nucleotide activating protein |
| GTP-GEF | G protein bound-guanine nucleotide exchange factor |
| HDAC | Histone deacetylase complex |
| ICAM1 | Intercellular adhesion molecule 1 |
| IFN | Interferon |
| IL | Interleukin |
| KO | Knock-out (gene) |
| LIMMA | Linear models for microarray analysis |
| LF | Lymphocytic foci |
| LFA1 | Lymphocyte function-association antigen 1 |
| LTI | Lymphoid tissue inducerr cell |
| MADCAM1 | Mucosal cell adhesion molecule-1 |
| MZ | Marginal zone |
| Mapk | Mitogen-activated protein kinase |
| MRC | Marginal reticular cell |
| MZB | Marginal zone B cell |
| NF-kb | Nuclear factor-kb |
| NK | Natural killer (cell) |
| NOTCH | Notch gene homolog |
| PALS | Periarteriolar lymphoid sheaths |
| PTK2 | Protein tyrosine kinase |
| RAC | Ras-related C3 botulinum toxin substrate |
| RANKL | Receptor activator of NF-kb |
| rt-PCR | Real-time polymerase chain reaction |
| RA | Rheumatoid arthritis |
| RHO-GTP | Ras homolog member |
| RNA | Ribonucleic acid |
| RND | Rho-family GTPase |
| RSF | Remodeling and spacing factor |
| SLE | Systemic lupus erythematosus |
| SS | Sjögren’s syndrome |
| S1P | Sphingosin-1-phosphate |
| S1PR | Sphingosine-1-phosphate receptor |
| TG | Transgenic |
| TLR | Toll-like receptor |
| VCAM1 | Vascular cell adhesion molecule 1 |
| VLA4 | Very late antigen 4 |
References
- Vivino, F.B.; Bunya, V.Y.; Massaro-Giordano, G.; Johr, C.R.; Giattino, S.L.; Schorpion, A.; Shafer, B.; Peck, A.; Sivils, K.; Rasmussen, A.; et al. Sjogren’s syndrome: An update on disease pathogenesis, clinical manifestations and treatment. Clin. Immunol. 2019, 203, 81–121. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.E.; Grimbacher, B.; Witte, T. Autoimmunity and primary immunodeficiency: Two sides of the same coin? Nat. Rev. Rheumatol. 2018, 14, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, Y.; Zhu, H.; Zhao, M.; Lu, Q. The Pathogenic Role of Dysregulated Epigenetic Modifications in Autoimmune Diseases. Front. Immunol. 2019, 10, 2305. [Google Scholar] [CrossRef]
- Slight-Webb, S.; Bourn, R.L.; Holers, V.M.; James, J.A. Shared and unique immune alterations in pre-clinical autoimmunity. Curr. Opin. Immunol. 2019, 61, 60–68. [Google Scholar] [CrossRef]
- Teng, X.; Cornaby, C.; Li, W.; Morel, L. Metabolic regulation of pathogenic autoimmunity: Therapeutic targeting. Curr. Opin. Immunol. 2019, 61, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Marketos, N.; Cinoku, I.; Rapti, A.; Mavragani, C.P. Type I interferon signature in Sjogren’s syndrome: Pathophysiological and clinical implications. Clin. Exp. Rheumatol. 2019, 37, S185–S191. [Google Scholar]
- Brito-Zeron, P.; Theander, E.; Baldini, C.; Seror, R.; Retamozo, S.; Quartuccio, L.; Bootsma, H.; Bowman, S.J.; Dorner, T.; Gottenberg, J.E.; et al. Early diagnosis of primary Sjogren’s syndrome: EULAR-SS task force clinical recommendations. Expert Rev. Clin. Immunol. 2016, 12, 137–156. [Google Scholar] [CrossRef]
- Fisher, B.A.; Jonsson, R.; Daniels, T.; Bombardieri, M.; Brown, R.M.; Morgan, P.; Bombardieri, S.; Ng, W.F.; Tzioufas, A.G.; Vitali, C.; et al. Sjogren’s histopathology workshop group from, E. Standardisation of labial salivary gland histopathology in clinical trials in primary Sjogren’s syndrome. Ann. Rheum. Dis. 2017, 76, 1161–1168. [Google Scholar] [CrossRef]
- Yin, H.; Wu, H.; Chen, Y.; Zhang, J.; Zheng, M.; Chen, G.; Li, L.; Lu, Q. The Therapeutic and Pathogenic Role of Autophagy in Autoimmune Diseases. Front. Immunol. 2018, 9, 1512. [Google Scholar] [CrossRef]
- Stojanov, S.; Kastner, D.L. Familial autoinflammatory diseases: Genetics, pathogenesis and treatment. Curr. Opin. Rheumatol. 2005, 17, 586–599. [Google Scholar] [CrossRef]
- Dorner, T. Crossroads of B cell activation in autoimmunity: Rationale of targeting B cells. J. Rheumatol. Suppl. 2006, 77, 3–11. [Google Scholar] [PubMed]
- Hofmann, K.; Clauder, A.K.; Manz, R.A. Targeting B Cells and Plasma Cells in Autoimmune Diseases. Front. Immunol. 2018, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, L.I.; Daoussis, D.; Mavropoulos, A.; Liossis, S.N.; Bogdanos, D.P. Regulatory B cells: New players in inflammatory and autoimmune rheumatic diseases. Semin. Arthritis Rheum. 2019, 48, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Kroese, F.G.M.; Abdulahad, W.H.; Haacke, E.; Bos, N.A.; Vissink, A.; Bootsma, H. B-cell hyperactivity in primary Sjogren’s syndrome. Expert Rev. Clin. Immunol. 2014, 10, 483–499. [Google Scholar] [CrossRef]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjogren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef]
- Nguyen, C.Q.; Cha, S.R.; Peck, A.B. Sjogren’s syndrome (SjS)-like disease of mice: The importance of B lymphocytes and autoantibodies. Front. Biosci. 2007, 12, 1767–1789. [Google Scholar] [CrossRef][Green Version]
- Lee, B.H.; Gauna, A.E.; Pauley, K.M.; Park, Y.J.; Cha, S. Animal Models in Autoimmune Diseases: Lessons Learned from Mouse Models for Sjogren’s Syndrome. Clin. Rev. Allergy Immunol. 2012, 42, 35–44. [Google Scholar] [CrossRef]
- Delaleu, N.; Nguyen, C.Q.; Peck, A.B.; Jonsson, R. Sjogren’s syndrome: Studying the disease in mice. Arthritis Res. Ther. 2011, 13, 217. [Google Scholar] [CrossRef]
- Cha, S.; Peck, A.B.; Humphreys-Beher, M.G. Progress in understanding autoimmune exocrinopathy using the non-obese diabetic mouse: An update. Crit. Rev. Oral Biol. Med. 2002, 13, 5–16. [Google Scholar] [CrossRef]
- Nguyen, C.Q.; Kim, H.; Cornelius, J.G.; Peck, A.B. Development of Sjogren’s syndrome in nonobese diabetic-derived autoimmune-prone C57BL/6.NOD-Aec1Aec2 mice is dependent on complement component-3. J. Immunol. 2007, 179, 2318–2329. [Google Scholar] [CrossRef]
- Robinson, C.P.; Yamachika, S.; Bounous, D.I.; Brayer, J.; Jonsson, R.; Holmdahl, R.; Peck, A.B.; Humphreys-Beher, M.G. A novel NOD-derived murine model of primary Sjogren’s syndrome. Arthritis Rheum. 1998, 41, 150–156. [Google Scholar] [CrossRef]
- Kong, L.; Robinson, C.P.; Peck, A.B.; Vela-Roch, N.; Sakata, K.M.; Dang, H.; Talal, N.; Humphreys-Beher, M.G. Inappropriate apoptosis of salivary and lacrimal gland epithelium of immunodeficient NOD-scid mice. Clin. Exp. Rheumatol. 1998, 16, 675–681. [Google Scholar] [PubMed]
- Cha, S.; Brayer, J.; Gao, J.; Brown, V.; Killedar, S.; Yasunari, U.; Peck, A.B. A dual role for interferon-gamma in the pathogenesis of Sjogren’s syndrome-like autoimmune exocrinopathy in the nonobese diabetic mouse. Scand. J. Immunol. 2004, 60, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.H.; Killedar, S.; Cornelius, J.G.; Nguyen, C.; Cha, S.H.; Peck, A.B. Sjogren’s syndrome in the NOD mouse model is an interleukin-4 time-dependent, antibody isotype-specific autoimmune disease. J. Autoimmun. 2006, 26, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.Q.; Gao, J.H.; Kim, H.; Saban, D.R.; Cornelius, J.G.; Peck, A.B. IL-4-STAT6 signal transduction-dependent induction of the clinical phase of Sjogren’s Syndrome-like disease of the nonobese diabetic mouse. J. Immunol. 2007, 179, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.; Cornelius, J.; Singson, E.; Killedar, S.; Cha, S.H.; Peck, A.B. Role of complement and B lymphocytes in Sjogren’s syndrome-like autoimmune exocrinopathy of NOD.B10-H2(B) mice. Mol. Immunol. 2006, 43, 1332–1339. [Google Scholar] [CrossRef]
- Nguyen, C. Pathogenic effect of interleukin-17A in induction of Sjogren syndrome-like disease using adenovirus-mediated gene transfer. Arthritis Res. Ther. 2010, 12, R220. [Google Scholar] [CrossRef]
- Shen, L.; Zhang, C.; Wang, T.; Brooks, S.; Ford, R.J.; Lin-Lee, Y.C.; Kasianowicz, A.; Kumar, V.; Martin, L.; Liang, P.; et al. Development of autoimmunity in IL-14α-transgenic mice. J. Immunol. 2006, 177, 5676–5686. [Google Scholar] [CrossRef]
- Shen, L.; Suresh, L.; Li, H.; Zhang, C.J.; Kumar, V.; Pankewycz, O.; Ambrus, J.L. IL-14 alpha, the nexus for primary Sjogren’s disease in mice and humans. Clin. Immunol. 2009, 130, 304–312. [Google Scholar] [CrossRef]
- Cha, S.; Nagashima, H.; Brown, V.B.; Peck, A.B.; Humphreys-Beher, M.G. Two NOD Idd-associated intervals contribute synergistically to the development of autoimmune exocrinopathy (Sjogren’s syndrome) on a healthy murine background. Arthritis Rheum. 2002, 46, 1390–1398. [Google Scholar] [CrossRef]
- Delaleu, N.; Nguyen, C.Q.; Tekle, K.M.; Jonsson, R.; Peck, A.B. Transcriptional landscapes of emerging autoimmunity: Transient aberrations in the targeted tissue’s extracellular milieu precede immune responses in Sjogren’s syndrome. Arthritis Res. Ther. 2013, 15, R174. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.B.; Saulep-Easton, D.; Figgett, W.A.; Fairfax, K.A.; Mackay, F. The BAFF/APRIL system: Emerging functions beyond B cell biology and autoimmunity. Cytokine Growth Factor Rev. 2013, 24, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhang, W.; Haskett, S.; Pellerin, A.; Xu, S.Q.; Petersen, B.; Jandreski, L.; Hamann, S.; Reynolds, T.L.; Zheng, T.S.; et al. BAFF overexpression increases lymphocytic infiltration in Sjogren’s target tissue, but only inefficiently promotes ectopic B-cell differentiation. Clin. Immunol. 2016, 169, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kiripolsky, J.; Klimatcheva, E.; Howell, A.; Fereidouni, F.; Levenson, R.; Rothstein, T.L.; Kramer, J.M. Early BAFF receptor blockade mitigates murine Sjogren’s syndrome: Concomitant targeting of CXCL13 and the BAFF receptor prevents salivary hypofunction. Clin. Immunol. 2016, 164, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Suresh, L.; Wu, J.; Xuan, J.; Li, H.; Zhang, C.; Pankewycz, O.; Ambrus, J.L. A Role for Lymphotoxin in Primary Sjogren’s Disease. J. Immunol. 2010, 185, 6355–6363. [Google Scholar] [CrossRef] [PubMed]
- Peck, A.B.; Nguyen, C.Q.; Ambrus, J. Early Covert Appearance of Marginal Zone B Cells in Salivary Glands of Sjogren’s Syndrome-Susceptible Mice: Initiators of Subsequent Overt Clinical Disease. Int. J. Mol. Sci. 2021, 22, 1919. [Google Scholar] [CrossRef] [PubMed]
- Peck, A.B.; Nguyen, C.Q.; Ambrus, J.L. Upregulated Chemokine and Rho-GTPase Genes Define Immune Cell Emigration into Salivary Glands of Sjogren’s Syndrome-Susceptible C57BL/6.NOD-Aec1Aec2 Mice. Int. J. Mol. Sci. 2021, 22, 16. [Google Scholar] [CrossRef]
- Doyle, M.E.; Boggs, L.; Attia, R.; Cooper, L.R.; Saban, D.R.; Nguyen, C.Q.; Peck, A.B. Autoimmune dacryoadenitis of NOD/Lt mice and its subsequent effects on tear protein composition. Am. J. Pathol. 2007, 171, 1224–1236. [Google Scholar] [CrossRef]
- Hammad, H.; Vanderkerken, M.; Pouliot, P.; Deswarte, K.; Toussaint, W.; Vergote, K.; Vandersarren, L.; Janssens, S.; Ramou, I.; Savvides, S.N.; et al. Transitional B cells commit to marginal zone B cell fate by Taok3-mediated surface expression of ADAM10. Nat. Immunol. 2017, 18, 313–320. [Google Scholar] [CrossRef]
- Arruga, F.; Vaisitti, T.; Deaglio, S. The NOTCH Pathway and Its Mutations in Mature B Cell Malignancies. Front. Oncol. 2018, 8, 550. [Google Scholar] [CrossRef]
- Pillai, S.; Cariappa, A. The follicular versus marginal zone B lymphocyte cell fate decision. Nat. Rev. Immunol. 2009, 9, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Kearney, J.F. Positive selection from newly formed to marginal zone B cells depends on the rate of clonal production, CD19, and btk. Immunity 2000, 12, 39–49. [Google Scholar] [CrossRef]
- Lopes-Carvalho, T.; Foote, J.; Kearney, J.F. Marginal zone B cells in lymphocyte activation and regulation. Curr. Opin. Immunol. 2005, 17, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Batista, F.D.; Harwood, N.E. The who, how and where of antigen presentation to B cells. Nat. Rev. Immunol. 2009, 9, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Cols, M.; Puga, I. Marginal zone B cells: Virtues of innate-like antibody-producing lymphocytes. Nat. Rev. Immunol. 2013, 13, 118–132. [Google Scholar] [CrossRef]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Bombardieri, M.; Raimondo, S.; Carubbi, F.; Cannizzaro, A.; Sireci, G.; Dieli, F.; Campisi, G.; et al. Interleukin (IL)-22 receptoris over-expressed in primary Sjogren’s syndrome and Sjogren-associated non-Hodgkin lymphomas and is regulated by IL-18. Clin. Exp. Immunol. 2015, 181, 219–229. [Google Scholar] [CrossRef]
- Chen, Y.; Pikkarainen, T.; Elomaa, O.; Soininen, R.; Kodama, T.; Kraal, G.; Tryggvason, K. Defective microarchitecture of the spleen marginal zone and impaired response to a thymus-independent type 2 antigen in mice lacking scavenger receptors MARCO and SR-A. J. Immunol. 2005, 175, 8173–8180. [Google Scholar] [CrossRef]
- Lu, T.T.; Cyster, J.G. Integrin-mediated long-term B cell retention in the splenic marginal zone. Science 2002, 297, 409–412. [Google Scholar] [CrossRef]
- Peck, A.B.; Saylor, B.T.; Nguyen, L.; Sharma, A.; She, J.-X.; Nguyen, C.Q.; McIndoe, R.A. Gene expression profiling of early-phase Sjogren’s syndrome in C57BL/6.NOD-Aec1Aec2 mice identifies focal adhesion maturation associated with infiltrating leukocytes. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5647–5655. [Google Scholar] [CrossRef]
- Tybulewicz, V.L.; Henderson, R.B. Rho family GTPases and their regulators in lymphocytes. Nat. Rev. Immunol. 2009, 9, 630–644. [Google Scholar] [CrossRef]
- Namekata, K.; Kimura, A.; Kawamura, K.; Harada, C.; Harada, T. Dock GEFs and their therapeutic potential: Neuroprotection and axon regeneration. Prog. Retin. Eye Res. 2014, 43, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gadea, G.; Blangy, A. Dock-family exchange factors in cell migration and disease. Eur. J. Cell Biol. 2014, 93, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.D.; Ridley, A.J. Rho GTPase signaling complexes in cell migration and invasion. J. Cell Biol. 2018, 217, 447–457. [Google Scholar] [CrossRef]
- Gotoh, K.; Tanaka, Y.; Nishikimi, A.; Inayoshi, A.; Enjoji, M.; Takayanagi, R.; Sasazuki, T.; Fukui, Y. Differential requirement for DOCK2 in migration of plasmacytoid dendritic cells versus myeloid dendritic cells. Blood 2008, 111, 2973–2976. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.P.; Yamamoto, H.; Peck, A.B.; Humphreys-Beher, M.G. Genetically programmed development of salivary gland abnormalities in the NOD (nonobese diabetic)-scid mouse in the absence of detectable lymphocytic infiltration: A potential trigger for sialoadenitis of NOD mice. Clin. Immunol. Immunopathol. 1996, 79, 50–59. [Google Scholar] [CrossRef]
- Schenke-Layland, K.; Xie, J.; Magnusson, M.; Angelis, E.; Li, X.; Wu, K.; Reinhardt, D.P.; MacLellan, W.R.; Hamm-Alvarez, S.F. Lymphocytic infiltration leads to degradation of lacrimal gland extracellular matrix structures in NOD mice exhibiting a Sjogren’s syndrome exocrinopathy. Exp. Eye Res. 2010, 90, 223–237. [Google Scholar] [CrossRef]
- Robinson, C.P.; Brayer, J.; Yamachika, S.; Esch, T.R.; Peck, A.B.; Stewart, C.A.; Peen, E.; Jonsson, R.; Humphreys-Beher, M.G. Transfer of human serum IgG to nonobese diabetic Igmu null mice reveals a role for autoantibodies in the loss of secretory function of exocrine tissues in Sjogren’s syndrome. Proc. Natl. Acad. Sci. USA 1998, 95, 7538–7543. [Google Scholar] [CrossRef]
- Tan, J.B.; Xu, K.; Cretegny, K.; Visan, I.; Yuan, J.S.; Egan, S.E.; Guidos, C.J. Lunatic and Manic Fringe Cooperatively Marginal Zone B Cell Precursor Competition for Delta-like 1 in Splenic Endothelial Niches. Immunity 2009, 30, 254–263. [Google Scholar] [CrossRef]
- Baker, N.; Ehrenstein, M.R. Cutting edge: Selection of B lymphocyte subsets is regulated by natural IgM. J. Immunol. 2002, 169, 6686–6690. [Google Scholar] [CrossRef]
- Shen, L.; Gao, C.; Suresh, L.; Xian, Z.; Song, N.; Chaves, L.D.; Yu, M.; Ambrus, J.L., Jr. Central role for marginal zone B cells in an animal model of Sjogren’s syndrome. Clin. Immunol. 2016, 168, 30–36. [Google Scholar] [CrossRef]
- Karlsen, M.; Jakobsen, K.; Jonsson, R.; Hammenfors, D.; Hansen, T.; Appel, S. Expression of Toll-Like Receptors in Peripheral Blood Mononuclear Cells of Patients with Primary Sjogren’s Syndrome. Scand. J. Immunol. 2017, 85, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Hillion, S.; Arleevskaya, M.I.; Blanco, P.; Bordron, A.; Brooks, W.H.; Cesbron, J.Y.; Bordron, A.; Vivier, E.; Brooks, W. The Innate Part of the Adaptive Immune System. Clin. Rev. Allergy Immunol. 2020, 58, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.C.; Guinamard, R.; Bolland, S.; Sankala, M.; Steinman, R.M.; Ravetch, J.V. Macrophages control the retention and trafficking of B lymphocytes in the splenic marginal zone. J. Exp. Med. 2003, 198, 333–340. [Google Scholar] [CrossRef]
- Cinamon, G.; Matloubian, M.; Lesneski, M.J.; Xu, Y.; Low, C.; Lu, T.; Proia, R.L.; Cyster, J.G. Sphingosine 1-phosphate receptor 1 promotes B cell localization in the splenic marginal zone. Nat. Immunol. 2004, 5, 713–720. [Google Scholar] [CrossRef]
- Cinamon, G.; Zachariah, M.A.; Lam, O.M.; Foss, F.W., Jr.; Cyster, J.G. Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat. Immunol. 2008, 9, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Girkontaite, L.; Missy, K.; Sakk, V.; Harenberg, A.; Tedford, K.; Potzel, T.; Pfeffer, K.; Fischer, H.D. Lsc is required for marginal zone B cells, regulation of lymphocyte motility and immune responses. Nat. Immunol. 2001, 2, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, I.; Tsubota, K.; Satake, Y.; Kita, Y.; Matsumura, R.; Murata, H.; Namekawa, T.; Nishioka, K.; Iwamoto, I.; Saitoh, Y.; et al. Common T cell receptor clonotype in lacrimal glands and labial salivary glands from patients with Sjogren’s syndrome. J. Clin. Investig. 1996, 97, 1969–1977. [Google Scholar] [CrossRef]
- Katsifis, G.E.; Moutsopoulos, N.M.; Wahl, S.M. T lymphocytes in Sjogren’s syndrome: Contributors to and regulators of pathophysiology. Clin. Rev. Allergy Immunol. 2007, 32, 252–264. [Google Scholar] [CrossRef]
- Fujihara, T.; Fujita, H.; Tsubota, K.; Saito, K.; Tsuzaka, K.; Abe, T.; Takeuchi, T. Preferential localization of CD8+ αEβ7+ T cells around acinar epithelial cells with apoptosis in patients with Sjogren’s syndrome. J. Immunol. 1999, 163, 2226–2235. [Google Scholar]
- Hayashi, T.; Shimoyama, N.; Mizuno, T. Destruction of Salivary and Lacrimal Glands by Th1-Polarized Reaction in a Model of Secondary Sjogren’s Syndrome in Lupus-Prone Female NZB × NZWF1 Mice. Inflammation 2012, 35, 638–646. [Google Scholar] [CrossRef]
- Tedford, K.; Steiner, M.; Koshutin, S.; Richter, K.; Tech, L.; Eggers, Y.; Jansing, I.; Schilling, K.; Hauser, A.E.; Korthals, M.; et al. The opposing forces of shear flow and sphingosine-1-phosphate control marginal zone B cell shuttling. Nat. Commun. 2017, 8, 2261. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Carvalho, T.; Kearney, J.F. Development and selection of marginal zone B cells. Immunol. Rev. 2004, 197, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Allam, R.; Anders, H.J. The role of innate immunity in autoimmune tissue injury. Curr. Opin. Rheumatol. 2008, 20, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Theofilopoulos, A.N.; Kono, D.H.; Baccala, R. The multiple pathways to autoimmunity. Nat. Immunol. 2017, 18, 716–724. [Google Scholar] [CrossRef]
- Weidenbusch, M.; Kulkarni, O.P.; Anders, H.J. The innate immune system in human systemic lupus erythematosus. Clin. Sci. 2017, 131, 625–634. [Google Scholar] [CrossRef]
- Oliver, A.M.; Martin, F.; Gartland, G.L.; Carter, R.H.; Kearney, J.F. Marginal Zone B Cells Exhibit Unique Activation, Proliferative and Immunoglobulin Secretory Responses. Eur. J. Immunol. 1997, 27, 2366–2374. [Google Scholar] [CrossRef]
- Colonna, M. Innate Lymphoid Cells: Diversity, Plasticity, and Unique Functions in Immunity. Immunity 2018, 48, 1104–1117. [Google Scholar] [CrossRef]
- Magri, G.; Miyajima, M.; Bascones, S.; Mortha, A.; Puga, I.; Cassis, L.; Barra, C.M.; Comerma, L.; Chudnovskiy, A.; Gentile, M.; et al. Innate lymphoid cells integrate stromal and immunological signals to enhance antibody production by splenic marginal zone B cells. Nat. Immunol. 2014, 15, 354–364. [Google Scholar] [CrossRef]
- Sozzani, S.; Bosisio, D.; Scarsi, M.; Tincani, A. Type I interferons in systemic autoimmunity. Autoimmunity 2010, 43, 196–203. [Google Scholar] [CrossRef]
- Cui, J.; Li, Y.; Zhu, L.; Liu, D.; Zhou, S.; Wang, H.Y.; Wang, R.-F. NLRP4 negatively regulates type I interferon signaling by targeting the kinase TBK1 for degradation via the ubiquitin ligase DTX4. Nat. Immunol. 2012, 13, 387–395. [Google Scholar] [CrossRef]
- He, B.; Santamaria, R.; Xu, W.; Cols, M.; Chen, K.; Puga, I.; Shan, M.; Xiong, H.; Bussel, J.B.; Chiu, A.; et al. The transmembrane activator TACI triggers immunoglobulin class switching by activating B cells through the adaptor MyD88. Nat. Immunol. 2010, 11, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Brigl, M.; Brenner, M.B. How invariant natural killer T cells respond to infection by recognizing microbial or endogenous lipid antigens. Semin. Immunol. 2010, 22, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Marino, E.; Batten, M.; Groom, J.; Walters, S.; Liuwantara, D.; Mackay, F.; Grey, S.T. Marginal-zone B-Cells of Nonobese diabetic mice expand with diabetes onset, invade the pancreatic lymph nodes, and present autoantigen to diabetogenic T-Cells. Diabetes 2008, 57, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, A.; Strauch, P.; DiGiacomo, A.; Hu, J.; Pelanda, R.; Torres, R.M. Lsc regulates marginal-zone B cell migration and adhesion and is required for the IgM T-dependent antibody response. Immunity 2005, 23, 527–538. [Google Scholar] [CrossRef]
- Nguyen, C.Q.; Cornelius, J.G.; Cooper, L.; Neff, J.; Tao, J.; Lee, B.H.; Peck, A.B. Identification of possible candidate genes regulating Sjogren’s syndrome-associated autoimmunity: A potential role for TNFSF4 in autoimmune exocrinopathy. Arthritis Res. Ther. 2008, 10, R137. [Google Scholar] [CrossRef]
- Tedford, K.; Nitschke, L.; Girkontaite, I.; Charlesworth, A.; Chan, G.; Sakk, V.; Barbacid, M.; Fischer, K.D. Compensation between Vav-1 and Vav-2 in B cell development and antigen receptor signaling. Nat. Immunol. 2001, 2, 548–555. [Google Scholar] [CrossRef]
- Wang, J.; Lin, F.; Wan, Z.; Sun, X.; Lu, Y.; Huang, J.; Wang, F.; Zeng, Y.; Chen, Y.-H.; Shi, Y.; et al. Profiling the origin, dynamics, and function of traction force in B cell activation. Sci. Signal. 2018, 11, 542. [Google Scholar] [CrossRef]
- Matsuzawa, K.; Akita, H.; Watanabe, T.; Kakeno, M.; Matsui, T.; Wang, S.; Kaibuchi, K. PAR3-aPKC regulates Tiam1 by modulating suppressive internal interactions. Mol. Biol. Cell 2016, 27, 1511–1523. [Google Scholar] [CrossRef]
- Garcia-Serna, A.M.; Alcaraz-Garcia, M.J.; Ruiz-Lafuente, N.; Sebastian-Ruiz, S.; Martinez, C.M.; Moya-Quiles, M.R.; Minguela, A.; Garcia-Alonso, A.M.; Martin-Orozco, E.; Parrado, A. Dock10 regulates CD23 expression and sustains B-cell lymphopoiesis in secondary lymphoid tissue. Immunobiology 2016, 221, 1343–1350. [Google Scholar] [CrossRef]
- Nombela-Arrieta, C.; Lacalle, R.A.; Montoya, M.C.; Kunisaki, Y.; Megias, D.; Marques, M.; Carrera, A.C.; Manes, S.; Fukui, Y.; Martinez-A, C.; et al. Differential requirements for DOCK2 and phosphoinositide-3-kinase γ during T and B lymphocyte homing. Immunity 2004, 21, 429–441. [Google Scholar] [CrossRef]
- Ushijima, M.; Uruno, T.; Nishikimi, A.; Sanematsu, F.; Kamikaseda, Y.; Kunimura, K.; Sakata, D.; Okada, T.; Fukui, Y. The Rac Activator DOCK2 Mediates Plasma Cell Differentiation and IgG Antibody Production. Front. Immunol. 2018, 9, 243. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.Q.; Sharma, A.; She, J.X.; McIndoe, R.A.; Peck, A.B. Differential gene expressions in the lacrimal gland during development and onset of keratoconjunctivitis sicca in Sjogren’s syndrome (SJS)-like disease of the C57BL/6.NOD-Aec1Aec2 mouse. Exp. Eye Res. 2009, 88, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.Q.; Sharma, A.; Lee, B.H.; She, J.X.; McIndoe, R.A.; Peck, A.B. Differential gene expression in the salivary gland during development and onset of xerostomia in Sjogren’s syndrome-like disease of the C57BL/6.NOD-Aec1Aec2 mouse. Arthritis Res. Ther. 2009, 11, R56. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Brill-Dashoff, J.; Shinton, S.A.; Asano, M.; Hardy, R.R.; Hayakawa, K. Evidence of marginal-zone B cell-positive selection in spleen. Immunity 2005, 23, 297–308. [Google Scholar] [CrossRef]
- Benjanini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).