
Associative and Physical Mapping of Markers Related to Fusarium in Maize Resistance, Obtained by Next-Generation Sequencing (NGS)

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
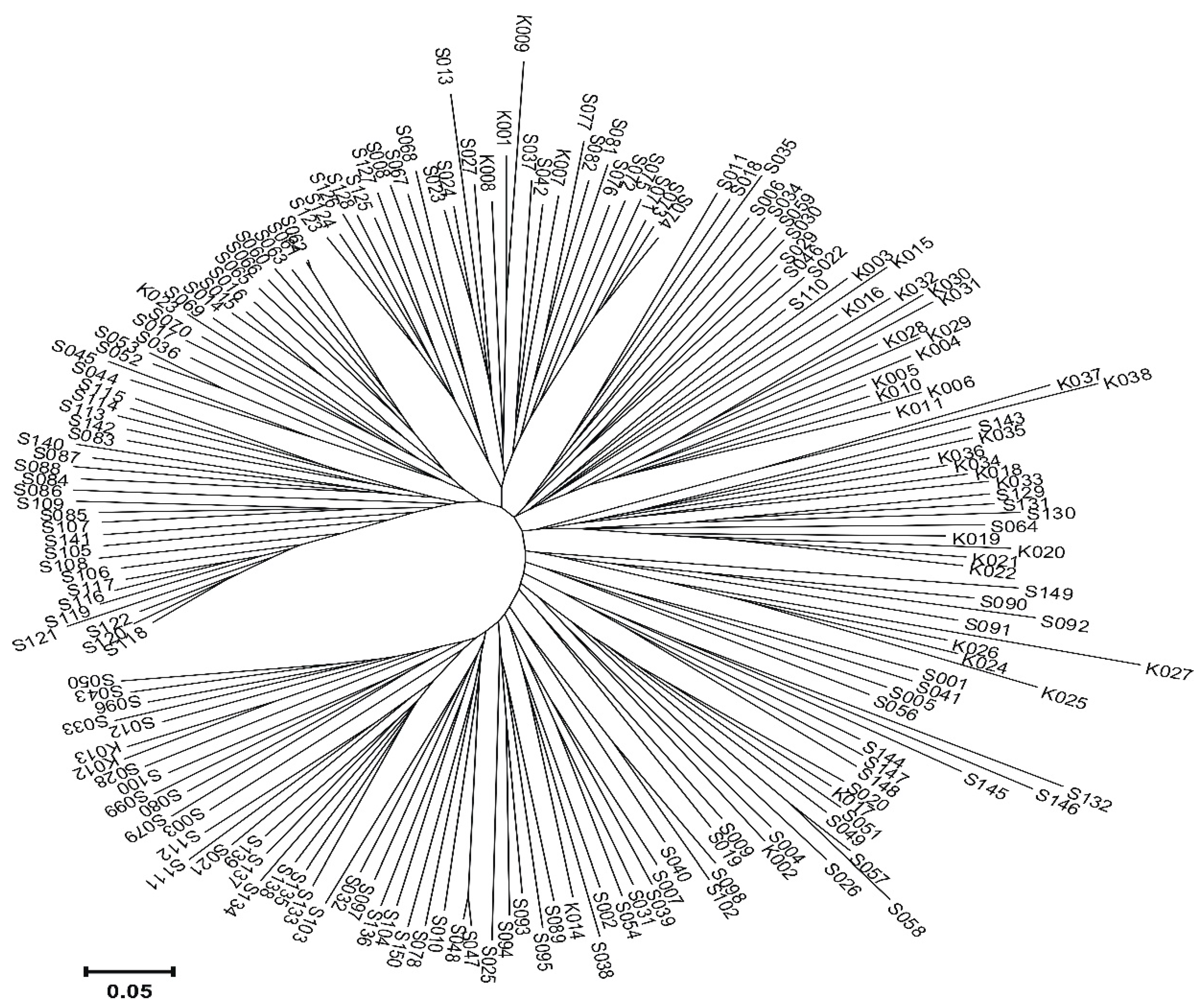
2. Results
2.1. Phenotyping
2.2. DNA Isolation
2.3. Genotyping
2.4. Associative Mapping Using GWAS Analysis
2.5. Physical Mapping and Functional Analysis of Gene Sequences
2.6. Design of Primers for Identified SilicoDArT and SNP Polymorphisms Associated with Fusarium Resistance of Maize Plants
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Methods
4.2.1. Phenotyping
4.2.2. DNA Isolation
4.2.3. Genotyping
4.2.4. Statistical Analysis
4.2.5. Associative Mapping Using GWAS Analysis
4.2.6. Physical Mapping
4.2.7. Functional Analysis of Gene Sequences
4.2.8. Designing Primers for Identified SilicoDArT and SNP Polymorphisms Related to Fusarium Resistance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yadav, M.K.; Lipsa, D.; Sandeep, R. Evaluation of Host Plant Resistance Traits in Certain Genotypes of Maize (Zea mays L.) against Maize Spotted Stem Borer (Chilo Partellus Swinhoe, Crambidae). Int. J. Entomol. Res. 2021, 6, 64–69. Available online: www.entomologyjournals.com (accessed on 1 April 2022).
- Zijlstra, C.; Lund, I.; Justesen, A.; Nicolaisen, M.; Bianciotto, V.; Posta, K.; Balestrini, R.; Przetakiewicz, A.; Czembor, E.; van de Zande, J. Combining novel monitoring tools and precision application technologies for integrated high-tech crop protection in the future (a discussion document). Pest Manag. Sci. 2011, 67, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Vasileiadis, V.P.; Otto, S.; Sattin, M.; Palinkás, Z.; Veres, A.; Bán, R.; Kiss, J.; Pons, X.; Kudsk, P.; Weide, R.; et al. Crop protection in European maize-based cropping systems: Current practices and recommendations for innovative Integrated Pest Management. Agric. Syst. 2011, 104, 533–540. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Lemmens, M.; Reid, L.M. Breeding for resistance to ear rot caused by Fusarium spp. in maize—A review. Plant Breed. 2012, 131, 1–19. [Google Scholar] [CrossRef]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Predicting mycotoxins in foods: A review. Food Microbiol. 2009, 26, 757–769. [Google Scholar] [CrossRef]
- Garcia-Ceron, D.; Lowe, R.G.T.; McKenna, J.A.; Brain, L.M.; Dawson, C.S.; Clark, B.; Berkowitz, O.P.; Whelan, J.; Bleackley, M.R.; Anderson, M.A. Extracellular Vesicles from Fusarium graminearum Contain Protein Effectors Expressed during Infection of Corn. J. Fungi 2021, 7, 977. [Google Scholar] [CrossRef]
- Meissler, M.; Mouron, P.; Musa, T.; Bigler, F.; Pons, X.; Vasileiadis, V.P.; Otto, S.; Antichi, D.; Kiss, J.; Pálinkás, Z.; et al. Pests, pesticide use and alternative options in European maize production: Current status and future prospects. J. Appl. Entomol. 2010, 34, 357–375. [Google Scholar]
- Uwe, L.; Miedaner, T.; Bürstmayr, H.; Vögele, R.T. Breeding for Resistance to Fusarium Ear Diseases in Maize and Small-Grain Cereals Using Genomic Tools. Ph.D. Thesis, Faculty of Agricultural Sciences University of Hohenheim, Stuttgart, Germany, 2020. [Google Scholar]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. In Epidemiology of Mycotoxin Producing Fungi; Xu, X., Bailey, J.A., Cooke, B.M., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 705–713. [Google Scholar] [CrossRef]
- Munkvold, G.P. Cultural and genetic approaches to managing mycotoxins in maize. Annu. Rev. Phytopathol. 2003, 41, 99–116. [Google Scholar] [CrossRef]
- Bush, B.J.; Carson, M.L.; Cubeta, M.A.; Hagler, W.M.; Payne, G.A. Infection and Fumonisin Production by Fusarium Verticillioide s in Developing Maize Kernels. Genet. Resist. Infect. Am. Phytopathol. Soc. 2003, 94, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Maschietto, V.; Cinzia, C.; Pirona, R.; Pea, G.; Strozzi, F.; Marocco, A.; Rossini, L.; Lanubile, A. QTL Mapping and Candidate Genes for Resistance to Fusarium Ear Rot and Fumonisin Contamination in Maize. BMC Plant Biol. 2017, 17, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, R.; Cao, A.; Malvar, R.A.; Butrón, A. Genomics of Maize Resistance to Fusarium Ear Rot and Fumonisin Contamination. Toxins 2020, 12, 431. [Google Scholar] [CrossRef] [PubMed]
- Voss-Fels, K.; Snowdon, R.J. Understanding and Utilizing Crop Genome Diversity via High-Resolution Genotyping. Plant Biotechnol. J. 2016, 14, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Maschietto, V.; Borrelli, V.M.; Stagnat, L.; Logrieco, A.; Marocco, F.A. Physiological and Molecular Plant Pathology Resistant and Susceptible Maize Genotypes Activate Different Transcriptional Responses against Fusarium Verticillioides. Physiol. Mol. Plant Pathol. 2012, 77, 52–59. [Google Scholar] [CrossRef]
- Campos-Bermudez, V.A.; Fauguel, C.M.; Tronconi, M.A.; Casati, P.; Presello, D.A.; Andreo, C.S. Transcriptional and Metabolic Changes Associated to the Infection by Fusarium verticillioides in Maize Inbreds with Contrasting Ear Rot Resistance. PLoS ONE 2013, 8, e61580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanubile, A.; Maschietto, V.; Borrelli, V.M.; Stagnat, L.; Logrieco, A.; Marocco, F.A. Molecular Basis of Resistance to Fusarium Ear Rot in Maize. Front. Plant Sci. Mol. 2017, 8, 1774. [Google Scholar] [CrossRef]
- Cruz Von, M.V.; Kilian, A.; Dierig, D.A. Development of DArT Marker Platforms and Genetic Diversity Assessment of the U.S. Collection of the New Oilseed Crop Lesquerella and Related Species. PLoS ONE 2013, 8, e64062. [Google Scholar] [CrossRef] [Green Version]
- Wittenberg, A.H.J.; Lee, T.; Cayla, C.; Kilian, A.; Visser, R.G.F.; Schouten, H. Validation of the high-throughput marker technology DArT using the model plant Arabidopsis thaliana. Mol. Genet. Genom. 2005, 274, 30–39. [Google Scholar] [CrossRef]
- Wenzl, P.; Carling, J.; Kudrna, D.; Jaccoud, D.; Huttner, E.; Kleinhofs, A.; Kilian, A. Diversity Arrays Technology (DArT) for Whole-Genome Profiling of Barley. Proc. Natl. Acad. Sci. USA 2004, 101, 9915–9920. [Google Scholar] [CrossRef] [Green Version]
- Zila, C.T.; Samayoa, L.F.; Santiago, R.; Butrón, A.; Holland, J.B. A genome-wide association study reveals genes associated with fusarium ear rot resistance in a maize core diversity panel. G3-Genes Genom. Genet. 2013, 3, 2095–2104. [Google Scholar] [CrossRef] [Green Version]
- Zila, C.T.; Ogut, F.; Romay, M.C.; Gardner, C.A.; Buckler, E.S. Genome wide association study of Fusarium ear rot disease in the U.S.A. maize inbred line collection. BMC Plant Biol. 2014, 14, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yubo, L.; Zhang, G.; Loladze, A.; Hu, A.; Wang, Y.; Zhang, H.Q.J.; Olsen, X.; San Vicente, M.; Crossa, F.; Lin, J.; et al. Genome-Wide Association Study and Genomic Prediction of Fusarium Ear Rot Resistance in Tropical Maize Germplasm. Crop J. 2021, 9, 325–341. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Oldenburg, E.; Höppner, F.; Ellner, F.; Weinert, J. Fusarium diseases of maize associated with mycotoxin contamination of agricultural products intended to be used for food and feed. Mycotoxin Res. 2017, 33, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Eli, K.; Schaafsma, A.W.; Limay-Rios, V.; Hooker, D.C. Effect of pydiflumetofen on Gibberella ear rot and Fusarium mycotoxin accumulation in maize grain. World Mycotoxin J. 2021, 14, 495–512. [Google Scholar] [CrossRef]
- Gxasheka, M.; Wang, J.; Gunya, B.; Mbanjwa, V.; Louis, T.; Dlamini, T.; Gao, J. In vitro effect of some commercial fungicides on mycelial growth of Fusarium species causing maize ear rot disease in China. Arch. Phytopathol. Plant Prot. 2021, 54, 557–569. [Google Scholar] [CrossRef]
- Champeil, A.; Doré, T.; Fourbet, J.F. Fusarium head blight: Epidemiological origin of the effects of cultural practices on head blight attacks and the production of mycotoxins by Fusarium in wheat grains. Plant Sci. 2004, 166, 1389–1415. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Samardžija, M.; Jeličić, A.; Mitak, M.; Pleadin, J. Oestrogen effects of zearelenon in farm animals and risks for human and animal health. Vet. Stanica 2017, 48, 109–118. Available online: https://hrcak.srce.hr/222017 (accessed on 1 April 2022).
- Ghareeb, K.; Awad, W.A.; Boehm, J.; Zebeli, Q. Impacts of the feed contaminant deoxynivalenol on the intestine of monogastric animals: Poultry and swine. J. Appl. Toxicol. 2015, 35, 327–337. [Google Scholar] [CrossRef]
- Guerre, P. Fusariotoxins in avian species: Toxicokinetics, metabolism and persistence in tissues. Toxins 2015, 7, 2289–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, S.T.; Auvergne, A.; Bernard, G.; Bailly, J.D.; Tardieu, D.; Babile, R.; Guerre, P. Chronic effects of fumonisin B1 on ducks. Poultry Sci. 2005, 84, 22–28. [Google Scholar] [CrossRef]
- Roche 454 Sequencing. Multiplex Identifier (MID) Adaptors for Rapid Library Preparations 454. Life Sci. 2010, 006-2009, 1–7. [Google Scholar]
- Mardis, E.R. The impact of next-generation sequencing technology on genetics. Trends Genet. 2008, 24, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Curey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP Discovery and Genetic Mapping Using Sequenced RAD Markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef] [PubMed]
- Adolfatto, P.; Davison, D.; Erezyilmaz, D.; Hu, T.T.; Mast, J.; Sunayama-Morita, T.; Stern, D.L. Multiplexed shotgun genotyping for rapid and efficient genetic mapping. Genome Res. 2011, 21, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yeh, C.-T.; Tang, H.M.; Nettleton, D.; Schnable, P.S. Gene mapping via bulked segregant RNASeq (BSR-Seq). PLoS ONE 2012, 7, e36406. [Google Scholar] [CrossRef] [Green Version]
- Sansaloni, C.; Petroli, C.; Jaccoud, D.; Carling, J.; Detering, F.; Grattapaglia, D.; Kilian, A. Diversity Arrays Technology (DArT) and next-generation sequencing combined: Genome-wide, high throughput, highly informative genotyping for molecular breeding of Eucalyptus. BMC Proc. 2011, 5, P54. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Gul Arabzai, M.; Gul, H. Application Techniques of Molecular Marker and Achievement of Marker Assisted Selection (MAS) in Three Major Crops Rice, Wheat and Maize. Int. J. Res. Appl. Sci. Biotechnol. 2021, 8, 82–93. [Google Scholar] [CrossRef]
- Salah, N.; Milad, S.I.; El- Rouby, M.M.; Barakat, M.N. Identification of New Molecular Markers Linked to Maize Stalk Rot Disease Resistance (Fusarium Moniliforme) in Maize. Plant Omics J. 2016, 9, 12–18. [Google Scholar]
- Michel, S.; Wagner, C.; Nosenko, T.; Steiner, B.; Samad-Zamini, M.; Buerstmayr, M.; Mayer, K.; Buerstmayr, H. Merging, Genomics and Transcriptomics for Predicting Fusarium Head Blight Resistance in Wheat. Genes 2021, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Snooks, H.D.; Sang, S. The Chemistry and Health Benefits of Dietary Phenolamides. J. Agric. Food Chem. 2020, 68, 6248–6267. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Zhang, M.; Apea-Bah, F.B.; Beta, T. Hydroxycinnamic acid amide (HCAA) derivatives, flavonoid C-glycosides, phenolic acids and antioxidant properties of foxtail millet. Food Chem. 2019, 295, 214–223. [Google Scholar] [CrossRef]
- Tanabe, K.; Hojo, Y.; Shinya, T.; Galis, I. Molecular evidence for biochemical diversification of phenolamide biosynthesis in rice plants. J. Integr. Plant Biol. 2016, 58, 903–913. [Google Scholar] [CrossRef]
- Chen, H.; Jones, A.D.; Howe, G.A. Constitutive activation of the jasmonate signaling pathway enhances the production of secondary metabolites in tomato. FEBS Lett. 2006, 580, 2540–2546. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.F.; He, Y.; Strauch, R.; Olukolu, B.A.; Nielsen, D.; Li, X.; Balint-Kurti, P.J. Maize Homologs of Hydroxycinnamoyltransferase, a Key Enzyme in Lignin Biosynthesis, Bind the Nucleotide Binding Leucine-Rich Repeat Rp1 Proteins to Modulate the Defense Response. Plant Physiol. 2015, 169, 2230–2243. [Google Scholar] [CrossRef]
- Herrero, J.; Fernández-Pérez, F.; Yebra, T.; Novo-Uzal, E.; Pomar, F.; Pedreño, M.; Cuello, J.; Guéra, A.; Esteban-Carrasco, A.; Zapata, J.M. Bioinformatic and functional characterization of the basic peroxidase 72 from Arabidopsis thaliana involved in lignin biosynthesis. Planta 2013, 237, 1599–1612. [Google Scholar] [CrossRef]
- Fernández-Pérez, F.; Pomar, F.; Pedreño, M.A.; Novo-Uzal, E. Suppression of Arabidopsis peroxidase 72 alters cell wall and phenylpropanoid metabolism. Plant Sci. 2015, 239, 192–199. [Google Scholar] [CrossRef]
- Lanubile, A.; Bernardi, J.; Marocco, A.; Logrieco, A.; Paciolla, C. Differential activation of defense genes and enzymes in maize genotypes with contrasting levels of resistance to Fusarium verticillioides. Environ. Exp. Bot. 2012, 78, 39–46. [Google Scholar] [CrossRef]
- Tomkowiak, A.; Bocianowski, J.; Wolko, Ł.; Adamczyk, J.; Mikołajczyk, S.; Kowalczewski, P.Ł. Identification of Markers Associated with Yield Traits and Morphological Features in Maize (Zea mays L.). Plants 2019, 8, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilian, B.; Graner, A. NGS technologies for analyzing germplasm diversity in genebanks. Brief. Funct. Genom. 2012, 11, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starzycka-Korbas, E.; Weber, Z.; Matuszczak, M.; Bocianowski, J.; Budzianowski, G.; Stefanowicz, M.; Starzycki, M. The diversity of Sclerotinia sclerotiorum (Lib.) de Bary isolates from western Poland. J. Plant Pathol. 2021, 103, 185–195. [Google Scholar] [CrossRef]
- Wrońska-Pilarek, D.; Dering, M.; Bocianowski, J.; Lechowicz, K.; Kowalkowski, W.; Barzdajn, W.; Hauke-Kowalska, M. Pollen Morphology and Variability of Abies alba Mill. Genotypes from South-Western Poland. Forests 2020, 11, 1125. [Google Scholar] [CrossRef]
- Wrońska-Pilarek, D.; Jagodziński, A.M.; Bocianowski, J.; Marecik, M.; Janyszek-Sołtysiak, M. Pollen morphology and variability of Sambucus nigra L.–Adoxaceae. Biologia 2020, 75, 481–493. [Google Scholar] [CrossRef]
- Bocianowski, J.; Kozak, M.; Liersch, A.; Bartkowiak-Broda, I. A heuristic method of searching for interesting markers in terms of quantitative traits. Euphytica 2011, 181, 89–100. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Line Number | Line Name | The Degree of Infection (9-Point Scale) | Line Number | Line Name | The Degree of Infection (9-Point Scale) | Line Number | Line Name | The Degree of Infection (9-Point Scale) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Smolice | Kobierzyce | Smolice | Kobierzyce | Smolice | Kobierzyce | ||||||
| 1 | S001 | 7 | 7.7 | 63 | S063 | 9 | 9 | 125 | S125 | 9 | 8.7 |
| 2 | S002 | 8 | 8.3 | 64 | S064 | 9 | 9 | 126 | S126 | 9 | 8.7 |
| 3 | S003 | 9 | 9 | 65 | S065 | 7 | 8.7 | 127 | S127 | 9 | 9 |
| 4 | S004 | 9 | 9 | 66 | S066 | 9 | 9 | 128 | S128 | 7 | 7.3 |
| 5 | S005 | 9 | 9 | 67 | S067 | 9 | 9 | 129 | S129 | 8 | 8.3 |
| 6 | S006 | 9 | 9 | 68 | S068 | 9 | 9 | 130 | S130 | 9 | 8.7 |
| 7 | S007 | 9 | 8.7 | 69 | S069 | 9 | 9 | 131 | S131 | 8 | 8.7 |
| 8 | S008 | 8 | 8 | 70 | S070 | 9 | 9 | 132 | S132 | 7 | 7 |
| 9 | S009 | 9 | 9 | 71 | S071 | 9 | 9 | 133 | S133 | 8 | 8.3 |
| 10 | S010 | 9 | 9 | 72 | S072 | 9 | 9 | 134 | S134 | 8 | 7.5 |
| 11 | S011 | 9 | 9 | 73 | S073 | 9 | 8.7 | 135 | S135 | 9 | 8.7 |
| 12 | S012 | 9 | 9 | 74 | S074 | 9 | 8.7 | 136 | S136 | 8 | 9 |
| 13 | S013 | 7 | 7.7 | 75 | S075 | 9 | 9 | 137 | S137 | 8 | 8.3 |
| 14 | S014 | 9 | 9 | 76 | S076 | 9 | 8.7 | 138 | S138 | 9 | 9 |
| 15 | S015 | 9 | 9 | 77 | S077 | 9 | 8.7 | 139 | S139 | 9 | 8.7 |
| 16 | S016 | 8 | 8.3 | 78 | S078 | 9 | 9 | 140 | S140 | 7 | 6.7 |
| 17 | S017 | 9 | 9 | 79 | S079 | 9 | 9 | 141 | S141 | 9 | 9 |
| 17 | S018 | 8 | 9 | 80 | S080 | 9 | 9 | 142 | S142 | 9 | 9 |
| 18 | S019 | 9 | 9 | 81 | S081 | 9 | 9 | 143 | S143 | 9 | 9 |
| 20 | S020 | 9 | 8.7 | 82 | S082 | 9 | 9 | 144 | S144 | 9 | 8.7 |
| 21 | S021 | 9 | 9 | 83 | S083 | 9 | 9 | 145 | S145 | 9 | 9 |
| 22 | S022 | 8 | 8.3 | 84 | S084 | 9 | 9 | 146 | S146 | 9 | 9 |
| 23 | S023 | 9 | 9 | 85 | S085 | 9 | 8.7 | 147 | S147 | 9 | 9 |
| 24 | S024 | 8 | 9 | 86 | S086 | 8 | 9 | 148 | S148 | 9 | 8.7 |
| 25 | S025 | 9 | 9 | 87 | S087 | 8 | 9 | 149 | S149 | 8 | 8 |
| 26 | S026 | 9 | 9 | 88 | S088 | 9 | 9 | 150 | S150 | 9 | 9 |
| 27 | S027 | 9 | 9 | 89 | S089 | 9 | 9 | 151 | K001 | 9 | 9 |
| 28 | S028 | 9 | 9 | 90 | S090 | 9 | 9 | 152 | K002 | 9 | 8.7 |
| 29 | S029 | 9 | 9 | 91 | S091 | 8 | 8.3 | 153 | K003 | 9 | 8.7 |
| 30 | S030 | 9 | 9 | 92 | S092 | 9 | 9 | 154 | K004 | 9 | 9 |
| 31 | S031 | 9 | 8.7 | 93 | S093 | 8 | 8.7 | 155 | K005 | 8 | 8.3 |
| 32 | S032 | 9 | 9 | 94 | S094 | 9 | 8.7 | 156 | K006 | 8 | 8 |
| 33 | S033 | 9 | 9 | 95 | S095 | 9 | 8.7 | 157 | K007 | 9 | 8.7 |
| 34 | S034 | 9 | 8.7 | 96 | S096 | 9 | 9 | 158 | K008 | 9 | 9 |
| 35 | S035 | 7 | 8.3 | 97 | S097 | 9 | 9 | 159 | K009 | 8 | 8.3 |
| 36 | S036 | 9 | 9 | 98 | S098 | 9 | 9 | 160 | K010 | 8 | 8.7 |
| 37 | S037 | 9 | 8.7 | 99 | S099 | 9 | 9 | 161 | K011 | 9 | 9 |
| 38 | S038 | 9 | 9 | 100 | S100 | 9 | 8.7 | 162 | K012 | 9 | 9 |
| 39 | S039 | 8 | 9 | 101 | S101 | 9 | 9 | 163 | K013 | 9 | 9 |
| 40 | S040 | 9 | 9 | 102 | S102 | 9 | 9 | 164 | K014 | 9 | 9 |
| 41 | S041 | 9 | 9 | 103 | S103 | 9 | 9 | 165 | K015 | 9 | 9 |
| 42 | S042 | 9 | 9 | 104 | S104 | 9 | 9 | 166 | K016 | 8 | 8.3 |
| 43 | S043 | 9 | 9 | 105 | S105 | 9 | 8.7 | 167 | K017 | 9 | 8.7 |
| 44 | S044 | 9 | 9 | 106 | S106 | 8 | 8.3 | 168 | K018 | 9 | 9 |
| 45 | S045 | 9 | 8.7 | 107 | S107 | 8 | 7.7 | 169 | K019 | 9 | 9 |
| 46 | S046 | 9 | 9 | 108 | S108 | 9 | 8.7 | 170 | K020 | 8 | 8 |
| 47 | S047 | 9 | 9 | 109 | S109 | 9 | 9 | 171 | K021 | 9 | 9 |
| 48 | S048 | 8 | 9 | 110 | S110 | 9 | 8.7 | 172 | K022 | 8 | 8.3 |
| 49 | S049 | 7 | 7 | 111 | S111 | 8 | 8 | 173 | K023 | 9 | 9 |
| 50 | S050 | 9 | 9 | 112 | S112 | 9 | 8.7 | 174 | K024 | 9 | 9 |
| 51 | S051 | 7 | 7.3 | 113 | S113 | 9 | 8.7 | 175 | K025 | 9 | 9 |
| 52 | S052 | 9 | 9 | 114 | S114 | 9 | 8.7 | 176 | K026 | 9 | 9 |
| 53 | S053 | 8 | 8.3 | 115 | S115 | 9 | 9 | 177 | K027 | 9 | 9 |
| 54 | S054 | 9 | 9 | 116 | S116 | 9 | 9 | 178 | K028 | 7 | 7.7 |
| 55 | S055 | 9 | 9 | 117 | S117 | 9 | 9 | 179 | K029 | 9 | 9 |
| 56 | S056 | 9 | 9 | 118 | S118 | 9 | 9 | 180 | K030 | 8 | 8.3 |
| 57 | S057 | 9 | 9 | 119 | S119 | 9 | 9 | 181 | K031 | 9 | 9 |
| 58 | S058 | 9 | 8.7 | 120 | S120 | 8 | 8 | 182 | K032 | 8 | 8.3 |
| 59 | S059 | 9 | 9 | 121 | S121 | 9 | 9 | 183 | K033 | 9 | 9 |
| 60 | S060 | 9 | 9 | 122 | S122 | 9 | 8.7 | 184 | K034 | 9 | 9 |
| 61 | S061 | 9 | 8.7 | 123 | S123 | 8 | 8.3 | 185 | K035 | 9 | 9 |
| 62 | S062 | 9 | 9 | 124 | S124 | 6 | 5.7 | 186 | K036 | 9 | 9 |
| Source of Variation | The Number of Degrees of Freedom | F Statistic |
|---|---|---|
| Location | 1 | 0.18 |
| Lines | 251 | 16.22 *** |
| Location × line interaction | 251 | 25.73 *** |
| Location | Kobierzyce | Smolice | Total | |
|---|---|---|---|---|
| The number of significant markers | SilicoDArT | 136 | 185 | 321 |
| SNP | 1067 | 1574 | 2641 | |
| Total (Silico DArT and SNP) | 1203 | 1759 | 2962 | |
| Minimal effect | SilicoDArT | −1.234 | −0.279 | |
| SNP | −1.469 | −0.305 | ||
| Total (Silico DArT and SNP) | −1.469 | −0.305 | ||
| Maximal effect | Silico DArT | 1.381 | 0.269 | |
| SNP | 1.574 | 0.311 | ||
| Total (Silico DArT and SNP) | 1.574 | 0.311 | ||
| Average effect | Sicico DArT | 0.092 | 0.039 | |
| SNP | −0.043 | 0.008 | ||
| Total (Silico DArT and SNP) | −0.028 | 0.011 | ||
| Total effect | Silico DArT | 12.483 | 7.146 | |
| SNP | −46.064 | 12.99 | ||
| Total (Silico DArT and SNP) | −33.581 | 20.136 | ||
| Marker | Marker Type | Chromosome | Marker Location | Candidate Genes |
|---|---|---|---|---|
| 553 | DArT | Chr9 | 19345104 | A marker that is anchored in the gene GDSL esterase/lipase At4g01130 precursor uncharacterized precursor of the protein) (LOC100273960) |
| 10382 | DArT | Chr10 | 149495362 | 1182 bp at 5′ side: ubiquitin carboxyl-terminal hydrolase 3 (LOC100191221) 1718 bp at 3′ side: Heavy metal transport/detoxification superfamily protein (LOC100501931) |
| 13242 | DArT | Chr1 | 292840905 292841155 292841283 | Within the tRNA Cys, 66,700 bp at 5′ side: fasciclin-like arabinogalactan protein 16 precursor (LOC100191430) 90,541 bp at 3′ side: calcium dependent protein kinase 11 (LOC103644148) |
| 15097 | DArT | Chr2 | 203171066 | A marker that is anchored in putrescine hydroxycinnamyltransferase gene (LOC103649226) |
| 15156 | DArT | Chr5 | 215026162 | 50,422 bp at 5′ side: photosynthetic NDH subunit of subcomplex B 4 chloroplastic (Loc100276619) 408 bp at 3′ side: pseudogene (LOC103627720) and 40,871 bp: expansin alpha precursor 2 (LOC542648) |
| 58153 | SNP | Chr9 | 145274999 | 1499 bp at 5′ side: histon h2a (LOC103639303) 2328 bp at 3′ side: histon h2b.1-similar (LOC103639303) |
| 58771 | SNP | Chr3 | 40548812 | A marker that is anchored the peroxidase precursor gene 72 (LOC100282124) and pentatricopeptide repeat-containing protein At5g57250, mitochondrial (LOC103649988) |
| Marker | Primer Sequences | Annual Temperature (°C) | Product Size (bp) | |
|---|---|---|---|---|
| Forward | Reverse | |||
| 553 | TTGTCGACGTACACGACCG | TTCGGGTGCGTGAAAAGCTA | 60 | 116 |
| 10,382 | GCAGTGCGTCGTGCAGT | AAGCCGATCGAGTTTGTGTTT | 58 | 91 |
| 13,242 | ACCTGCAGATCAATAGTCAC | GGACCCTTTGTATCGAAAA | 52 | 122 |
| 15,097 | GGCTCACCTTCCCGTTCTAC | GTACGAAGGCACCAGGAACA | 59 | 107 |
| 15,156 | CCGACATCAAATGTCACAGCA | TGAGAAGACGACGACGAAGC | 59 | 151 |
| 58,153 | ACTGCAGTATGGGACCACAA | TGAAACATGCACCAAAATAAAATCC | 57 | 100 |
| 58,771 | TGCTAGCACAAGTGCATTTCAA | TGAAGGTGTTGCAAGCGAAT | 58 | 103 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobiech, A.; Tomkowiak, A.; Nowak, B.; Bocianowski, J.; Wolko, Ł.; Spychała, J. Associative and Physical Mapping of Markers Related to Fusarium in Maize Resistance, Obtained by Next-Generation Sequencing (NGS). Int. J. Mol. Sci. 2022, 23, 6105. https://doi.org/10.3390/ijms23116105
Sobiech A, Tomkowiak A, Nowak B, Bocianowski J, Wolko Ł, Spychała J. Associative and Physical Mapping of Markers Related to Fusarium in Maize Resistance, Obtained by Next-Generation Sequencing (NGS). International Journal of Molecular Sciences. 2022; 23(11):6105. https://doi.org/10.3390/ijms23116105
Chicago/Turabian StyleSobiech, Aleksandra, Agnieszka Tomkowiak, Bartosz Nowak, Jan Bocianowski, Łukasz Wolko, and Julia Spychała. 2022. "Associative and Physical Mapping of Markers Related to Fusarium in Maize Resistance, Obtained by Next-Generation Sequencing (NGS)" International Journal of Molecular Sciences 23, no. 11: 6105. https://doi.org/10.3390/ijms23116105
APA StyleSobiech, A., Tomkowiak, A., Nowak, B., Bocianowski, J., Wolko, Ł., & Spychała, J. (2022). Associative and Physical Mapping of Markers Related to Fusarium in Maize Resistance, Obtained by Next-Generation Sequencing (NGS). International Journal of Molecular Sciences, 23(11), 6105. https://doi.org/10.3390/ijms23116105