
Cellular Environment and Phenotypic Heterogeneity: How Data-Driven Modeling Finds the Smoking Gun


Abstract
:1. Introduction
2. Results
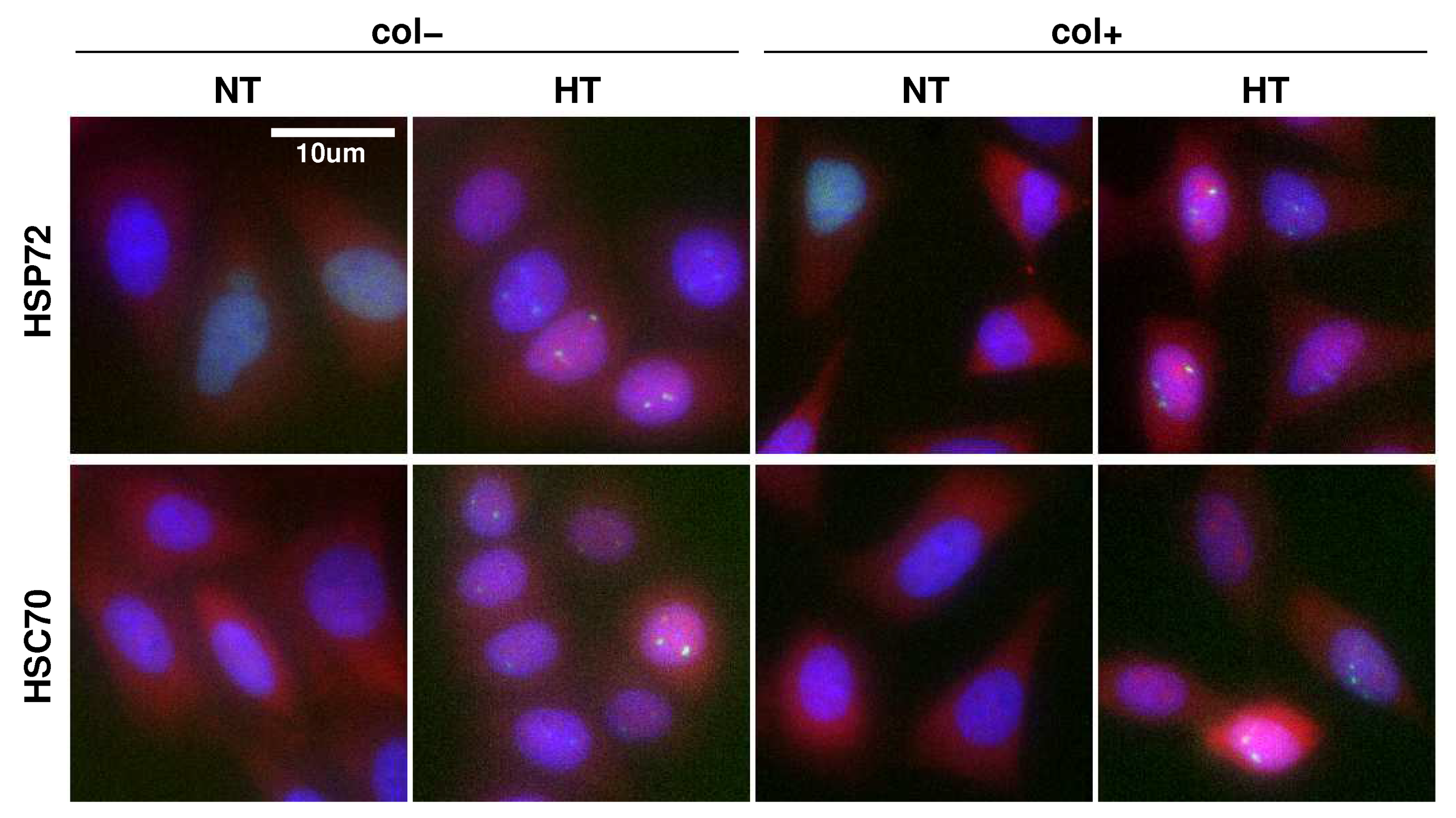
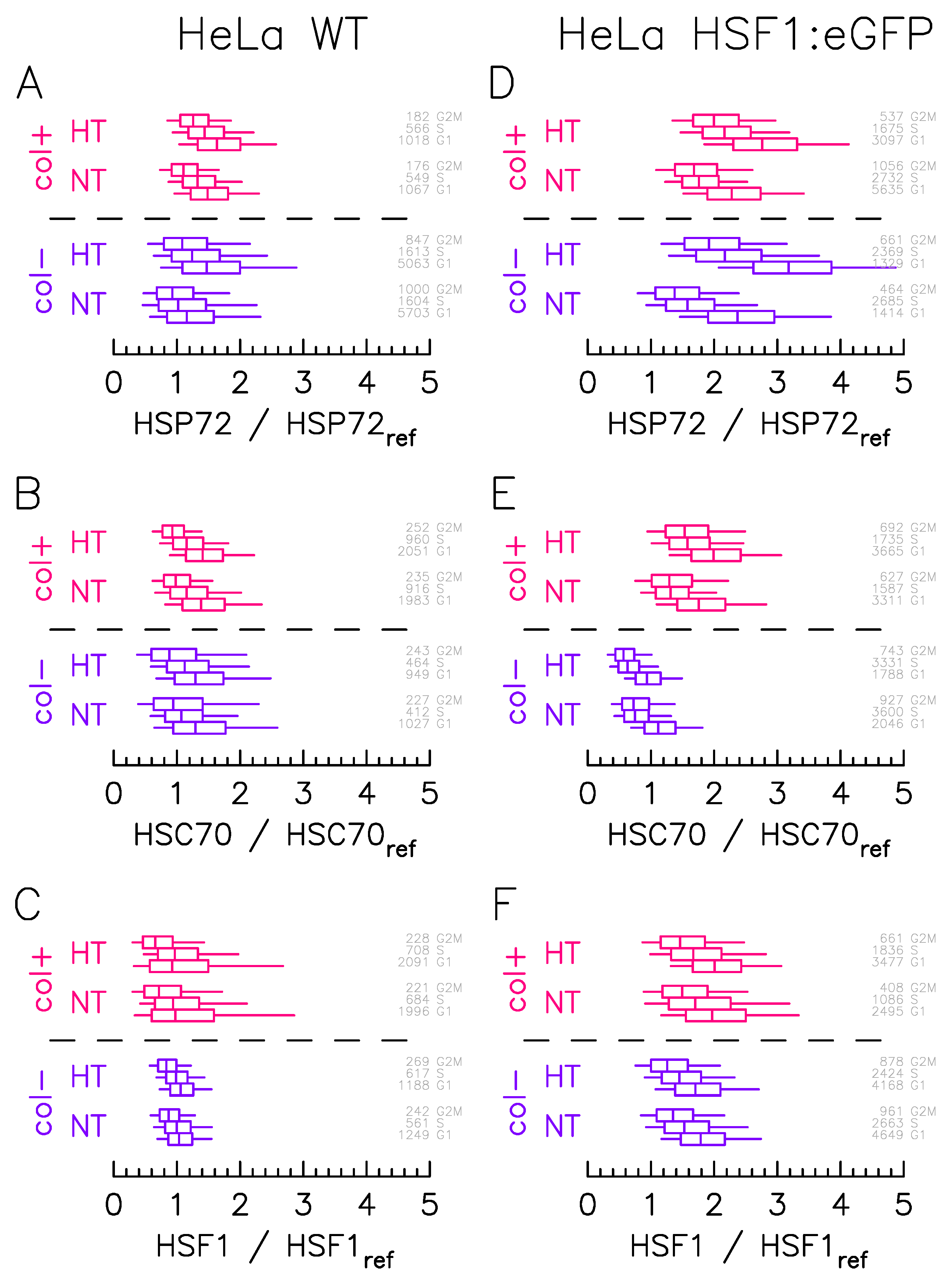
2.1. More Than Cell Cycle Phase, ECM Affects the Protein Expression Level and Variability
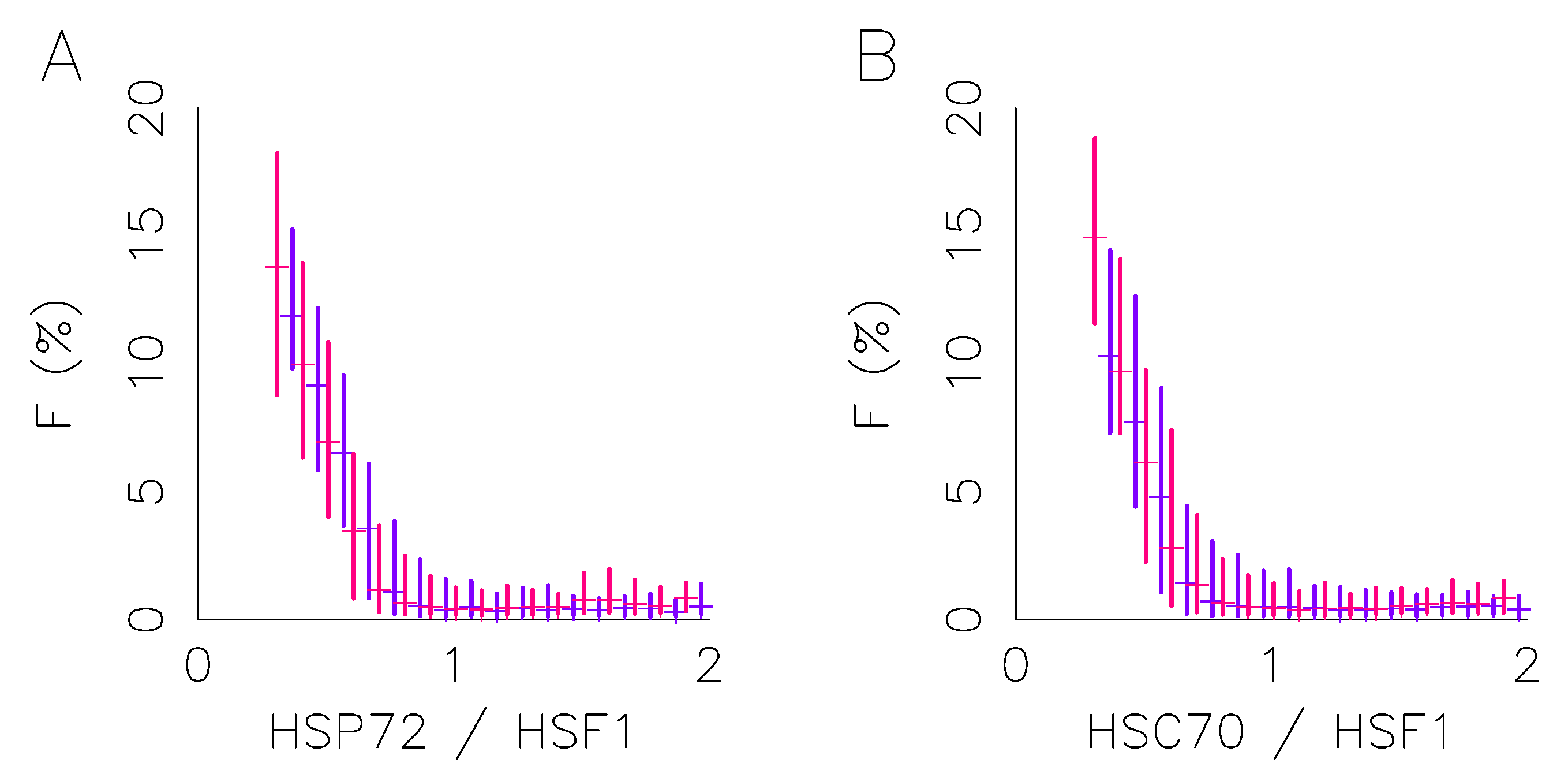
2.2. ECM Does Not Affect the Foci to Protein Expression Activation Function
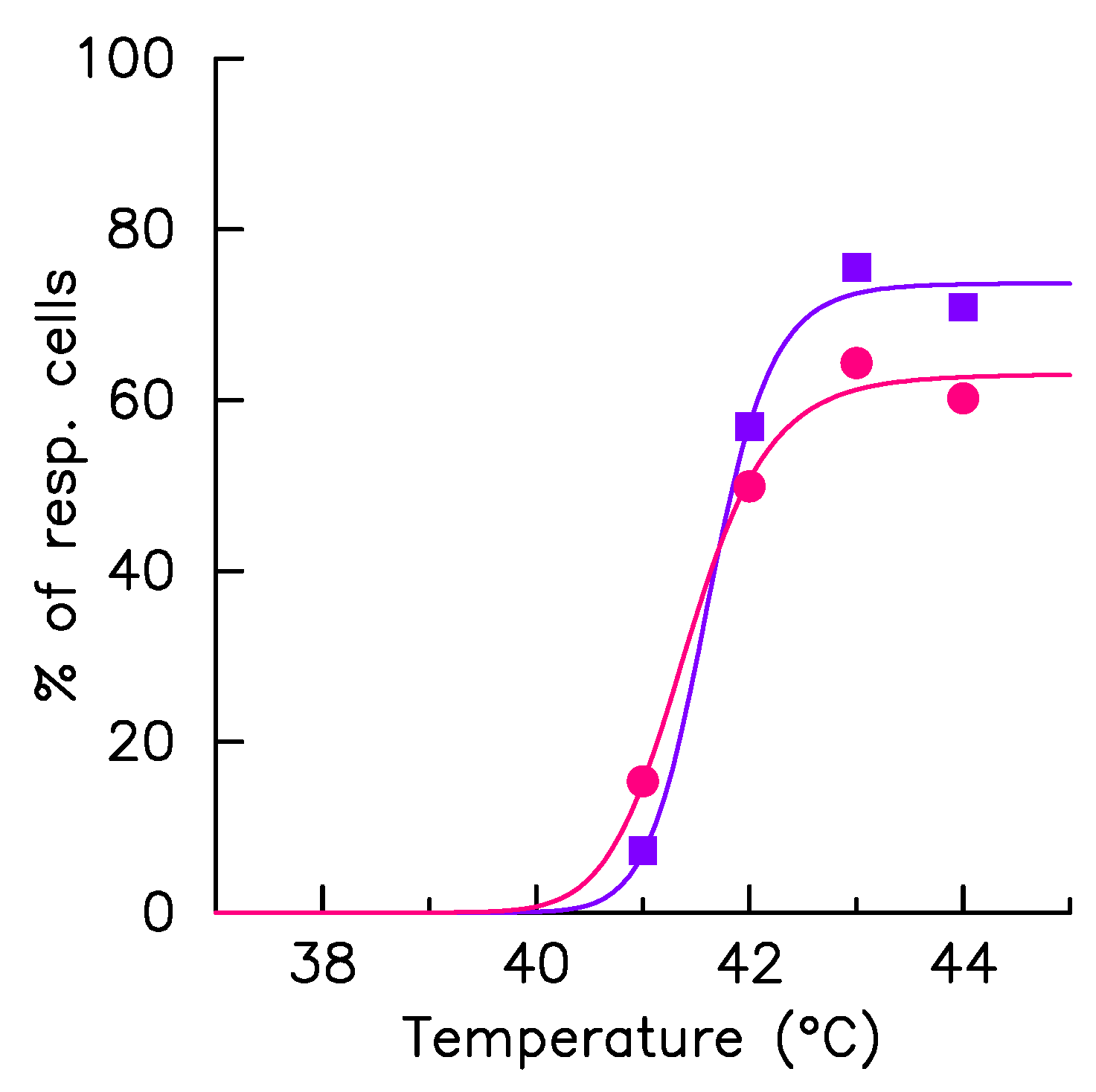
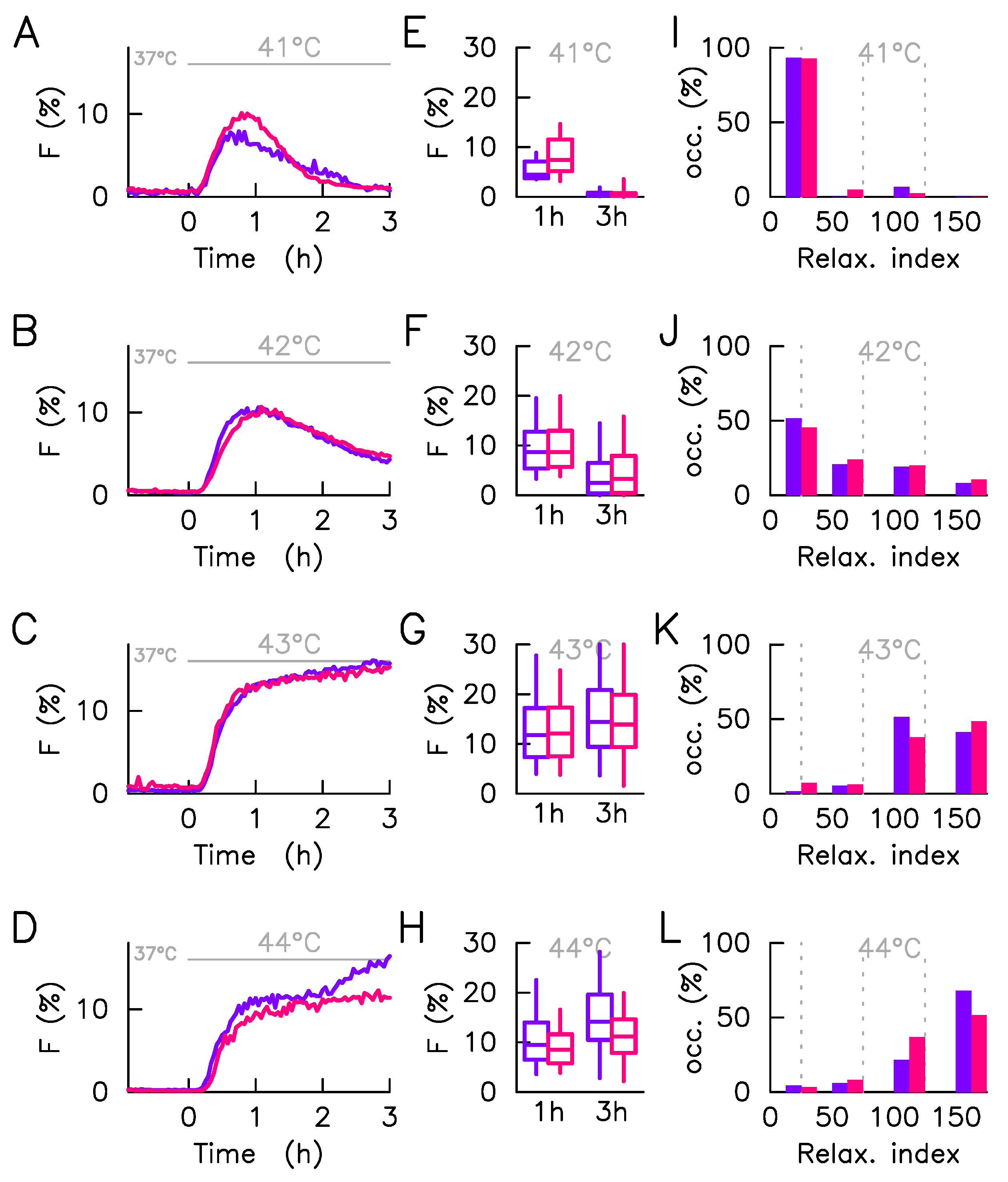
2.3. Monitoring Response Activation on HeLa Single Cell Level
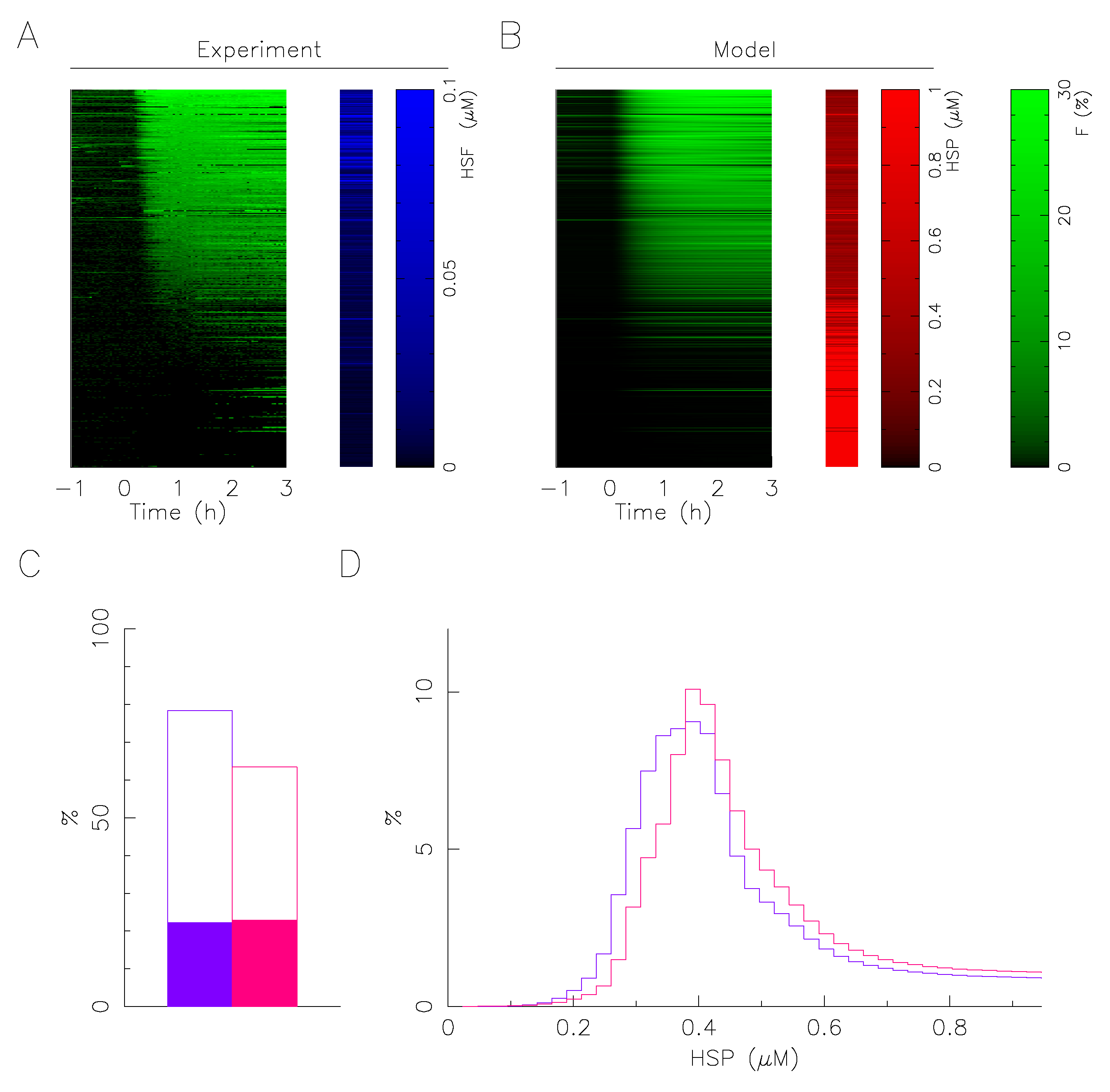
2.4. Mathematical Model Correlates Phenotypic Heterogeneity with Protein Expression Variability
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Cell Transfection
4.2. Collagen Coatings
4.3. Immunofluorescence Staining of HSPs and HSF1
4.4. Live/Fixed Cells Imaging
4.5. Image Processing and Analysis
4.6. Mathematical Model for HSRN
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | ExtraCellular Matrix |
| HSP70 | 70-kDa Heat Shock Protein family |
| HSP72 | Heat shock 70 kDa protein 1 (HSPA1A) |
| HSC70 | Heat shock 70 kDa protein 8 (HSPA8) |
| HSF1 | Heat shock factor 1 |
| NT | NormoThermia |
| HT | HyperThermia |
| F | Fraction of HSF1 fluorescence within foci |
References
- Hastings, J.F.; Skhinas, J.N.; Fey, D.; Croucher, D.R.; Cox, T.R. The extracellular matrix as a key regulator of intracellular signalling networks. Br. J. Pharmacol. 2019, 176, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Stamov, D.R.; Pompe, T. Structure and function of ECM-inspired composite collagen type I scaffolds. Soft Matter 2012, 8, 10200–10212. [Google Scholar] [CrossRef]
- Balázsi, G.; Van Oudenaarden, A.; Collins, J.J. Cellular decision making and biological noise: From microbes to mammals. Cell 2011, 144, 910–925. [Google Scholar] [CrossRef] [PubMed]
- Roux, J.; Hafner, M.; Bandara, S.; Sims, J.J.; Hudson, H.; Chai, D.; Sorger, P.K. Fractional killing arises from cell-to-cell variability in overcoming a caspase activity threshold. Mol. Syst. Biol. 2015, 11, 803. [Google Scholar] [CrossRef] [PubMed]
- Inde, Z.; Dixon, S.J. The impact of non-genetic heterogeneity on cancer cell death. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, K.S.; Charlebois, D.A.; Szenk, M.; Cohen, J.; Nevozhay, D.; Balázsi, G. Role of network-mediated stochasticity in mammalian drug resistance. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.C.; Kats, L.M. Non-genetic heterogeneity, altered cell fate and differentiation therapy. EMBO Mol. Med. 2021, 13, e12670. [Google Scholar] [CrossRef] [PubMed]
- Dayalan Naidu, S.; Dinkova-Kostova, A.T. Regulation of the mammalian heat shock factor 1. FEBS J. 2017, 284, 1606–1627. [Google Scholar] [CrossRef] [PubMed]
- Guilbert, M.; Anquez, F.; Pruvost, A.; Thommen, Q.; Courtade, E. Protein level variability determines phenotypic heterogeneity in proteotoxic stress response. FEBS J. 2020, 287, 5345–5361. [Google Scholar] [CrossRef] [PubMed]
- Biamonti, G.; Vourc’h, C. Nuclear stress bodies. Cold Spring Harb. Perspect. Biol. 2010, 95, a000695. [Google Scholar] [CrossRef] [PubMed]
- Sivéry, A.; Courtade, E.; Thommen, Q. A minimal titration model of the mammalian dynamical heat shock response. Phys. Biol. 2016, 13, 066008. [Google Scholar] [CrossRef] [PubMed]
- Tavaria, M.; Gabriele, T.; Kola, I.; Anderson, R.L. A hitchhiker’s guide to the human Hsp70 family. Cell Stress Chaperones 1996, 1, 23. [Google Scholar] [CrossRef]
- Walker, C.; Mojares, E.; del Río Hernández, A. Role of extracellular matrix in development and cancer progression. Int. J. Mol. Sci. 2018, 19, 3028. [Google Scholar] [CrossRef] [PubMed]
- Marusyk, A.; Almendro, V.; Polyak, K. Intra-tumour heterogeneity: A looking glass for cancer? Nat. Rev. Cancer 2012, 12, 323–334. [Google Scholar] [CrossRef]
- Haffner, M.C.; Zwart, W.; Roudier, M.P.; True, L.D.; Nelson, W.G.; Epstein, J.I.; De Marzo, A.M.; Nelson, P.S.; Yegnasubramanian, S. Genomic and phenotypic heterogeneity in prostate cancer. Nat. Rev. Urol. 2021, 18, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Sauk, J.J.; Van Kampen, C.L.; Norris, K.; Foster, R.; Somerman, M.J. Expression of constitutive and inducible HSP70 and HSP47 is enhanced in cells persistently spread on OPN1 or collagen. Biochem. Biophys. Res. Commun. 1990, 172, 135–142. [Google Scholar] [CrossRef]
- Foreman, R.; Wollman, R. Mammalian gene expression variability is explained by underlying cell state. Mol. Syst. Biol. 2020, 16, e9146. [Google Scholar] [CrossRef] [PubMed]
- Flusberg, D.A.; Sorger, P.K. Surviving apoptosis: Life-death signaling in single cells. Trends Cell Biol. 2015, 25, 446–458. [Google Scholar] [CrossRef]
- Herbomel, G.; Kloster-Landsberg, M.; Folco, E.G.; Col, E.; Usson, Y.; Vourc’h, C.; Delon, A.; Souchier, C. Dynamics of the full length and mutated heat shock factor 1 in human cells. PLoS ONE 2013, 8, e67566. [Google Scholar] [CrossRef]
- Anquez, F.; El Yazidi-Belkoura, I.; Randoux, S.; Suret, P.; Courtade, E. Cancerous cell death from sensitizer free photoactivation of singlet oxygen. Photochem. Photobiol. 2012, 88, 167–174. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Description | Value |
|---|---|---|---|
| (μM) | denaturation rate | 1.76 | |
| renaturation rate | 17.7 | ||
| (μM) | HSP basal transcription rate | 1.47 × | |
| (μM) | HSP active transcription rate | 0.78 | |
| (μM) | HSP transcription regulation threshold | 0.18 | |
| HSP translation rate | 10 | ||
| (μM) | translation regulation threshold | 0.32 | |
| (μM) | HSF1 concentration | 4.0 × | |
| (h) | incubator rise time | 1/15 | |
| (h) | MFP lifetime | 0.5 | |
| (h) | mHSP lifetime | 1 | |
| (h) | HSP lifetime time | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guilbert, M.; Courtade, E.; Thommen, Q. Cellular Environment and Phenotypic Heterogeneity: How Data-Driven Modeling Finds the Smoking Gun. Int. J. Mol. Sci. 2022, 23, 6536. https://doi.org/10.3390/ijms23126536
Guilbert M, Courtade E, Thommen Q. Cellular Environment and Phenotypic Heterogeneity: How Data-Driven Modeling Finds the Smoking Gun. International Journal of Molecular Sciences. 2022; 23(12):6536. https://doi.org/10.3390/ijms23126536
Chicago/Turabian StyleGuilbert, Marie, Emmanuel Courtade, and Quentin Thommen. 2022. "Cellular Environment and Phenotypic Heterogeneity: How Data-Driven Modeling Finds the Smoking Gun" International Journal of Molecular Sciences 23, no. 12: 6536. https://doi.org/10.3390/ijms23126536
APA StyleGuilbert, M., Courtade, E., & Thommen, Q. (2022). Cellular Environment and Phenotypic Heterogeneity: How Data-Driven Modeling Finds the Smoking Gun. International Journal of Molecular Sciences, 23(12), 6536. https://doi.org/10.3390/ijms23126536