
The Fate of Epidermal Tight Junctions in the stratum corneum: Their Involvement in the Regulation of Desquamation and Phenotypic Expression of Certain Skin Conditions

 ,
,
Abstract
1. Introduction
2. Results
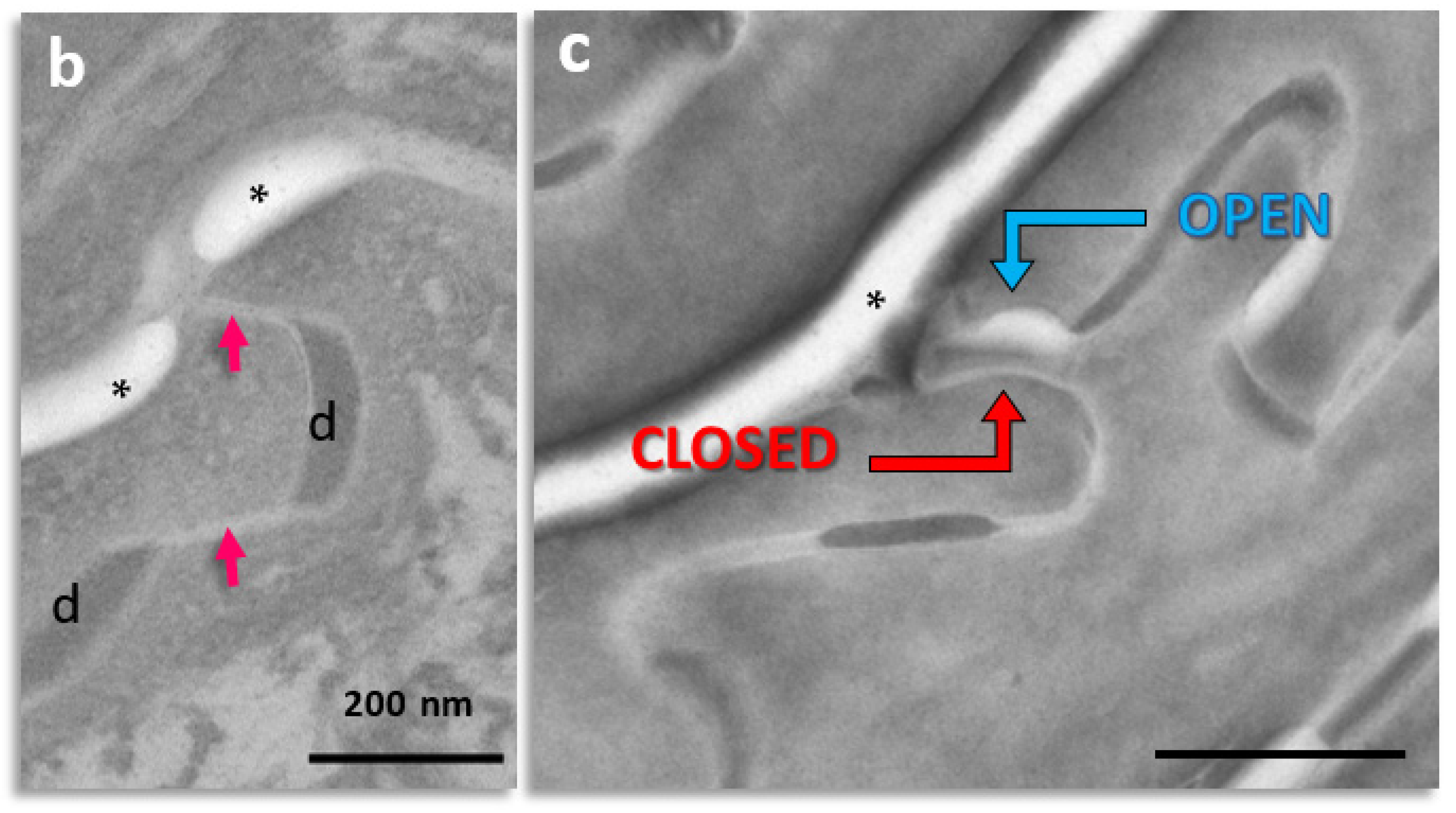
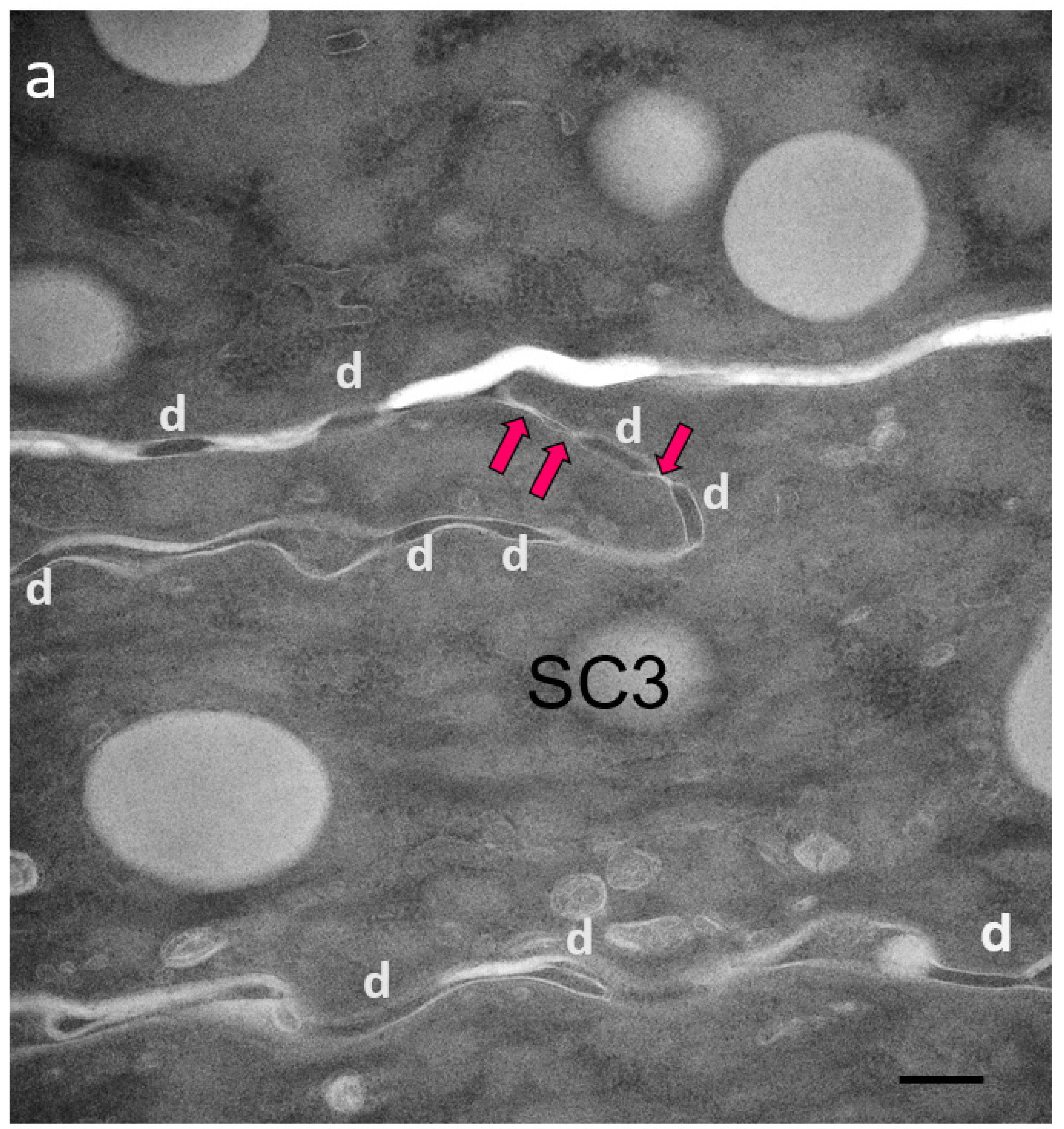
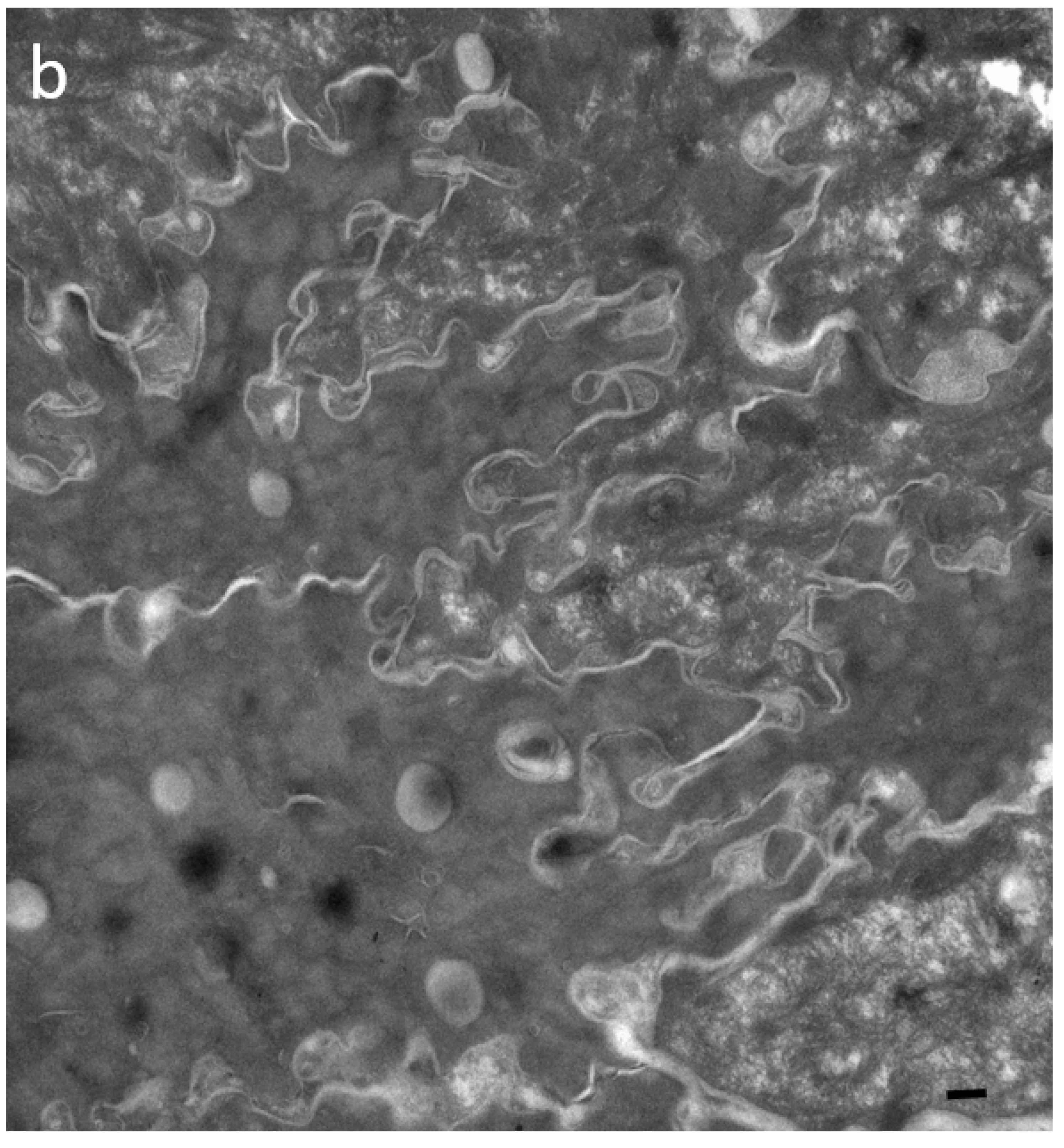
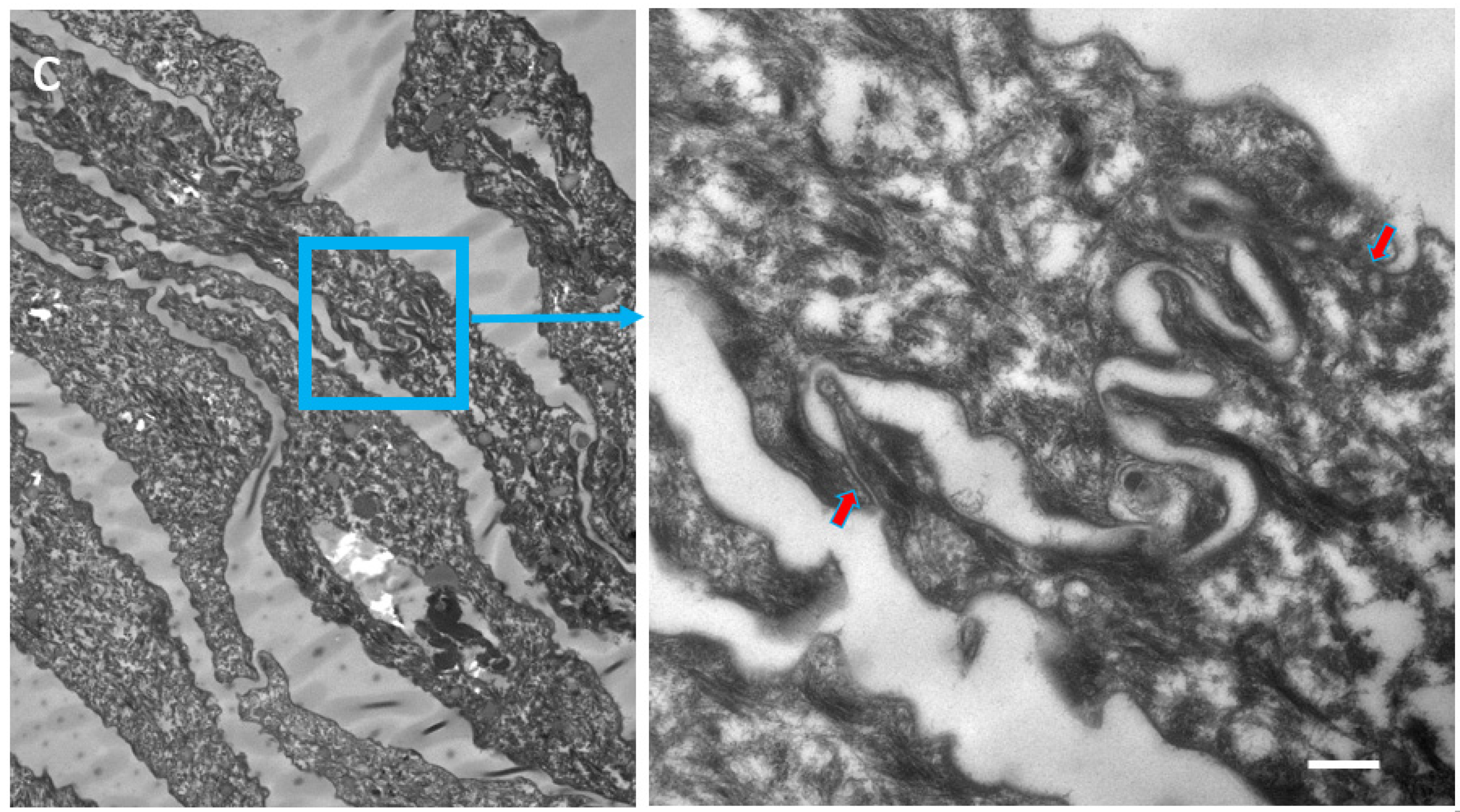
2.1. Visualization and Quantitation of TJ-Derived Points of Fusion between Corneocytes in the SC Compactum of Normal Human Skin (NHS)
2.2. Quantitation of Tight Junction Remnants in SC Reveals a Highly Interactive Line of Protection against Environmental Aggression
2.3. Regulation of SC Cohesion/Desquamation by Persistent TJ Remnants in Skin Pathology
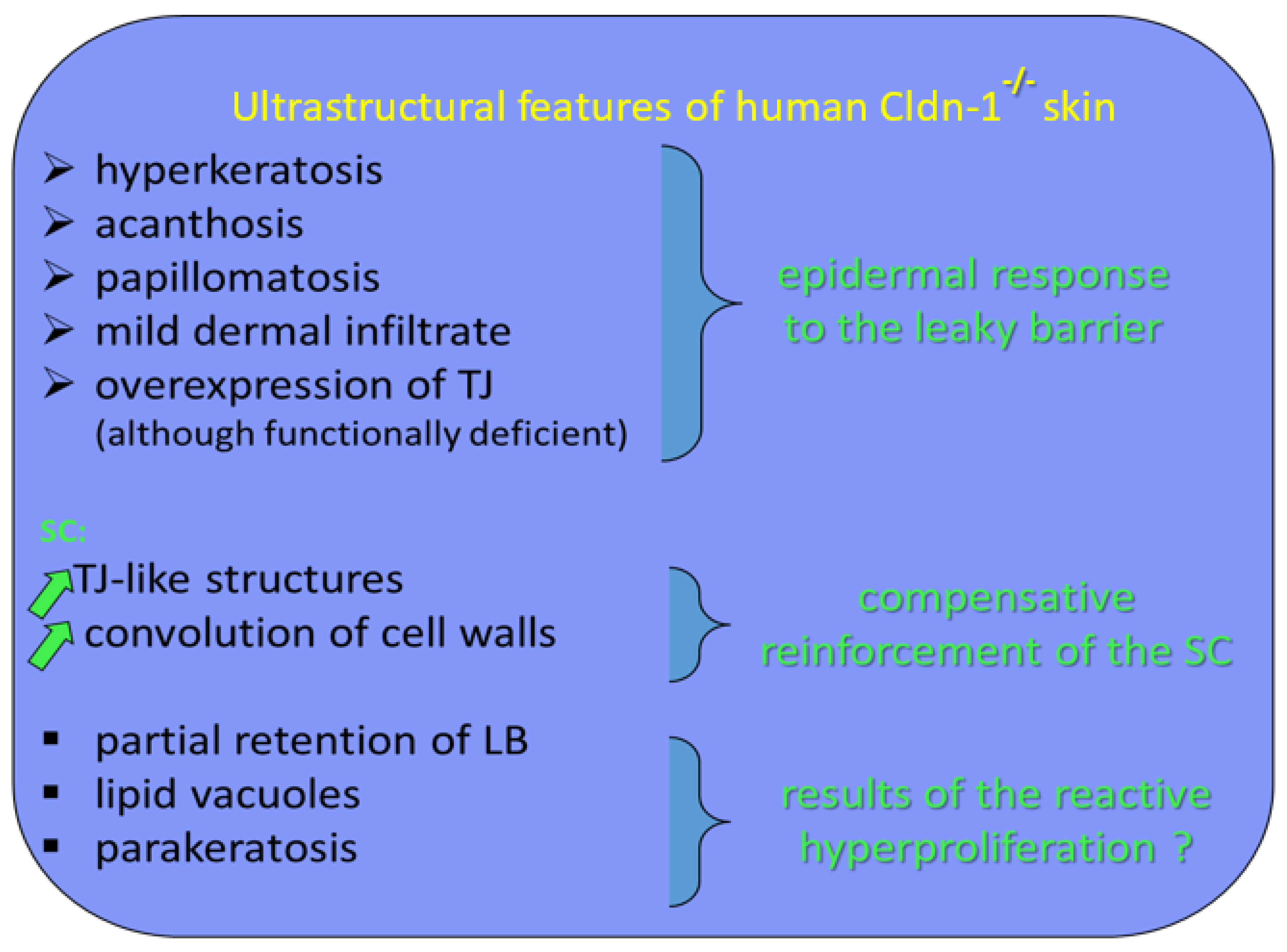
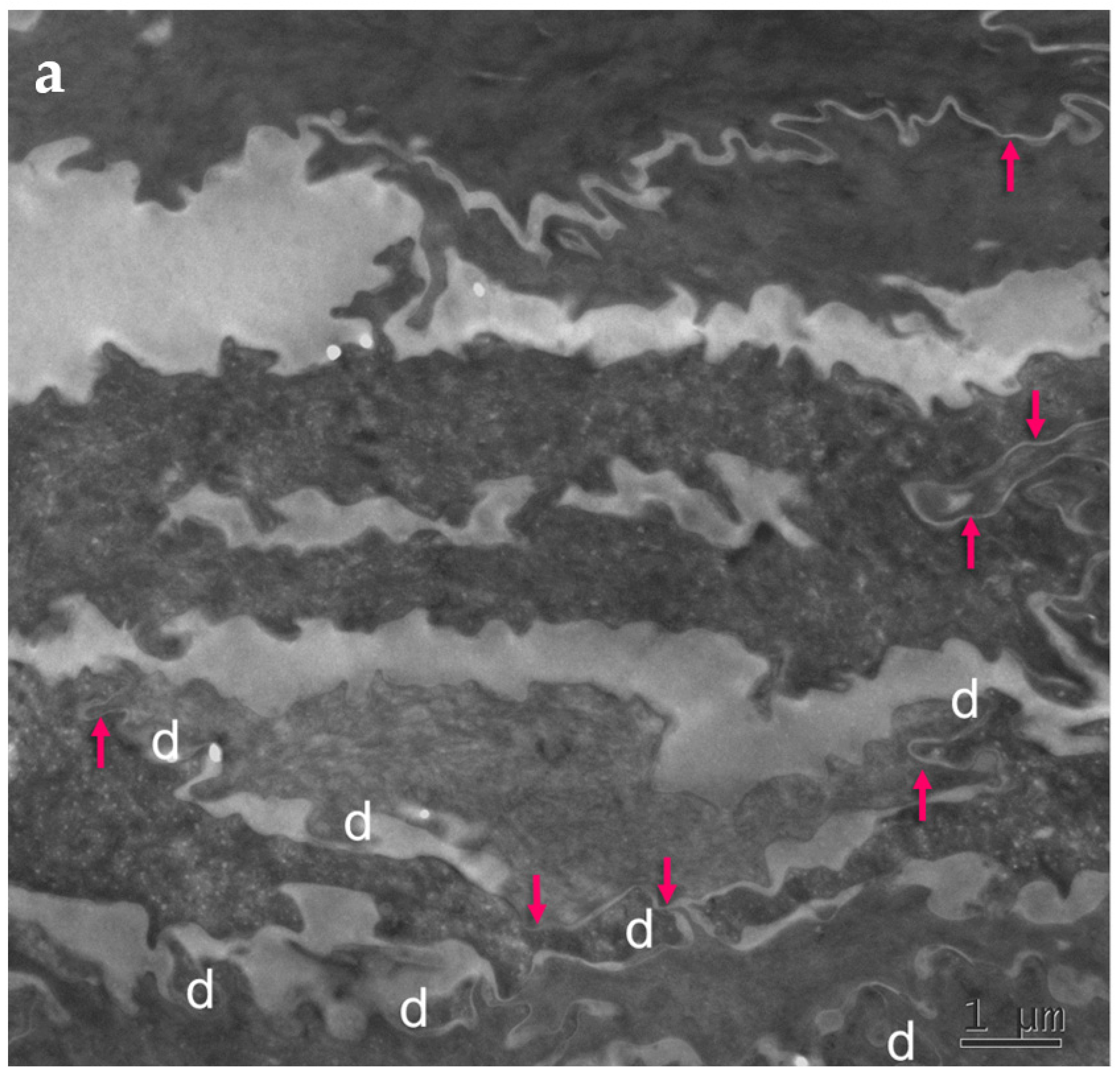
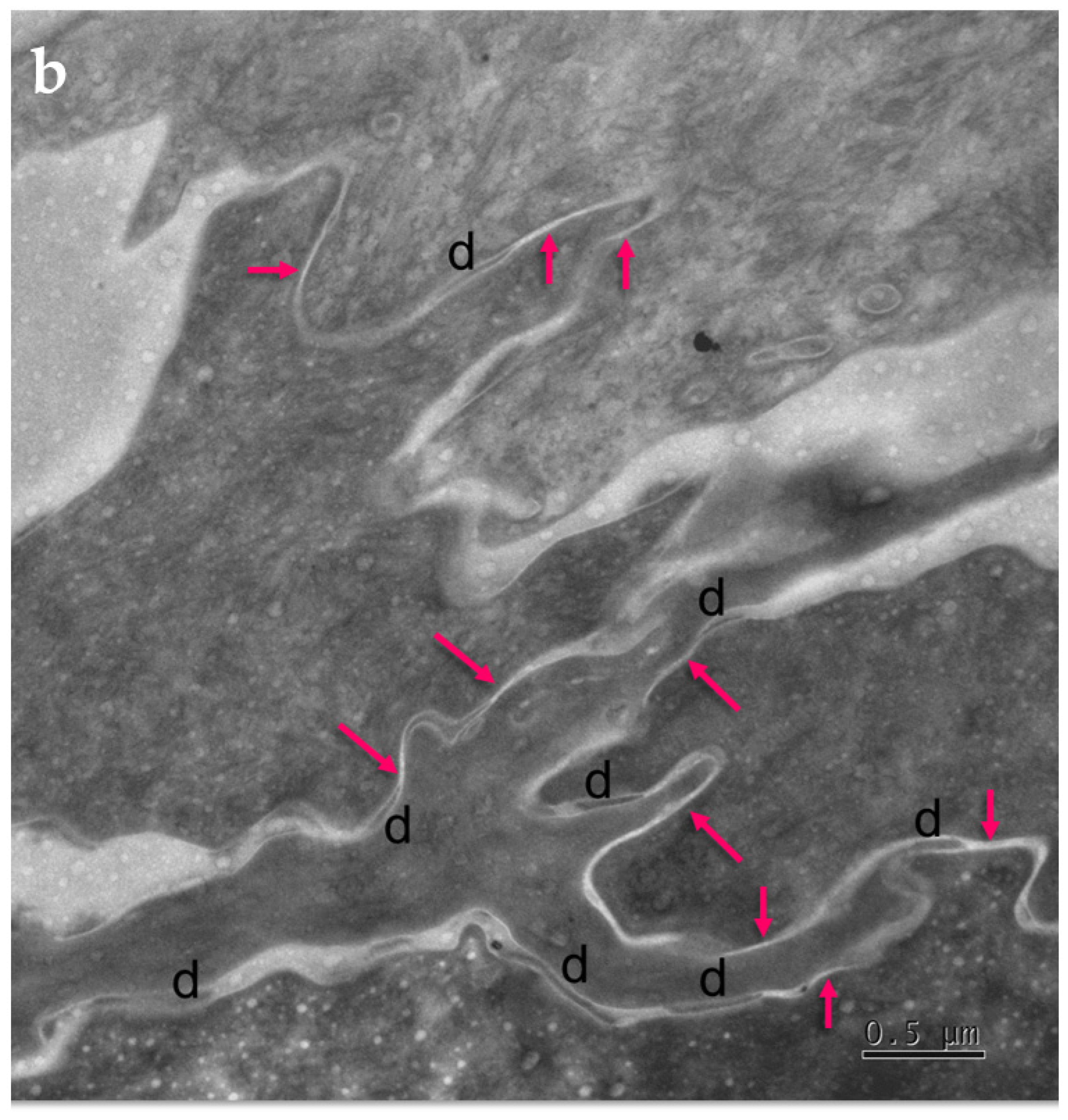
2.3.1. Tight Junction Remnants Contribute to the Compensatory Hyperkeratosis in Cldn1-Deficient Patients with Ichthyosis Hypotrichosis Sclerosing Cholangitis (IHSC) Syndrome
2.3.2. Ultrastructural Evaluation of the stratum corneum in Peeling Skin Disease (PSD) Suggests a Compensatory TJ Upregulation
3. Discussion
4. Material and Methods
4.1. Skin Samples from Healthy Volunteers and Patients
4.2. Electron Microscopy
4.3. Skin Explants
4.4. RHE
4.5. Quantitation of TJ Remnants
4.6. Statistical Analysis
5. Conclusions
- Constitute an additional permeability barrier, beneath the SC;
- Are involved in the process of formation of the SC barrier;
- Are mostly fragmented but may be up- or downregulated in SG;
- Their cross-linked TJ remnants participate in the SC cohesion and impact desquamation.
- The process of terminal differentiation and cornification;
- The persistence of TJ remnants in the SC and, thus, on desquamation;
- The permeability of the epidermal barrier.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haftek, M. ‘Memory’ of the stratum corneum: Exploration of the epidermis’ past. Br. J. Dermatol. 2014, 171 (Suppl. S3), 6–9. [Google Scholar] [CrossRef] [PubMed]
- Haftek, M.; Dragomir, A. Cell Junctions. In Ghadially’s Ultrastructural Pathology of the Cell and Matrix, 5th ed.; Tylor and Francis Group: London, UK, in press.
- Tsukita, S.; Furuse, M. Claudin-based barrier in simple and stratified cellular sheets. Curr. Opin. Cell Biol. 2002, 14, 531–536. [Google Scholar] [CrossRef]
- Yuki, T.; Haratake, A.; Koishikawa, H.; Morita, K.; Miyachi, Y.; Inoue, S. Tight junction proteins in keratinocytes: Localization and contribution to barrier function. Exp. Dermatol. 2007, 16, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Brandner, J.; Zorn-Kruppa, M.; Yoshida, T.; Moll, I.; Beck, L.; De Benedetto, A. Epidermal tight junctions in health and disease. Tissue Barriers 2015, 3, e974451. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, N.; Houdek, P.; Fromm, M.; Moll, I.; Brandner, J.M. Tight junctions form a barrier in human epidermis. Eur. J. Cell Biol. 2010, 89, 839–842. [Google Scholar] [CrossRef]
- Hashimoto, K. Intercellular Spaces of the Human Epidermis as Demonstrated with Lanthanum. J. Investig. Dermatol. 1971, 57, 17–31. [Google Scholar] [CrossRef][Green Version]
- Elias, P.M.; Menon, G.K. Structural and Lipid Biochemical Correlates of the Epidermal Permeability Barrier. Adv. Lipid Res. 1991, 24, 1–26. [Google Scholar] [CrossRef]
- Madison, K.C. Barrier Function of the Skin: “La Raison d’Être” of the Epidermis. J. Investig. Dermatol. 2003, 121, 231–241. [Google Scholar] [CrossRef]
- Bouwstra, J.A.; Ponec, M. The skin barrier in healthy and diseased state. Biochim. Biophys. Acta (BBA) Biomembr. 2006, 1758, 2080–2095. [Google Scholar] [CrossRef]
- Wertz, P. Roles of Lipids in the Permeability Barriers of Skin and Oral Mucosa. Int. J. Mol. Sci. 2021, 22, 5229. [Google Scholar] [CrossRef]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: A lesson from claudin-1-deficient mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Seker, M.; Fernández-Rodríguez, C.; Martínez-Cruz, L.A.; Müller, D. Mouse Models of Human Claudin-Associated Disorders: Benefits and Limitations. Int. J. Mol. Sci. 2019, 20, 5504. [Google Scholar] [CrossRef] [PubMed]
- Brandner, J.M.; Kief, S.; Grund, C.; Rendl, M.; Houdek, P.; Kuhn, C.; Tschachler, E.; Franke, W.W.; Moll, I. Organization and formation of the tight junction system in human epidermis and cultured keratinocytes. Eur. J. Cell Biol. 2002, 81, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Ishida-Yamamoto, A.; Kishibe, M.; Murakami, M.; Honma, M.; Takahashi, H.; Iizuka, H. Lamellar Granule Secretion Starts before the Establishment of Tight Junction Barrier for Paracellular Tracers in Mammalian Epidermis. PLoS ONE 2012, 7, e31641. [Google Scholar] [CrossRef] [PubMed]
- Yokouchi, M.; Kubo, A. Maintenance of tight junction barrier integrity in cell turnover and skin diseases. Exp. Dermatol. 2018, 27, 876–883. [Google Scholar] [CrossRef]
- Zorn-Kruppa, M.; Vidal-Y-Sy, S.; Houdek, P.; Wladykowski, E.; Grzybowski, S.; Gruber, R.; Gorzelanny, C.; Harcup, J.; Schneider, S.W.; Majumdar, A.; et al. Tight Junction barriers in human hair follicles—Role of claudin-1. Sci. Rep. 2018, 8, 12800. [Google Scholar] [CrossRef]
- Taïeb, A. Skin barrier in the neonate. Pediatr. Dermatol. 2018, 35 (Suppl. S1), s5–s9. [Google Scholar] [CrossRef]
- Choi, E.H. Aging of the skin barrier. Clin. Dermatol. 2019, 37, 336–345. [Google Scholar] [CrossRef]
- Krug, S.M.; Fromm, M. Special Issue on “The Tight Junction and Its Proteins: More than Just a Barrier”. Int. J. Mol. Sci. 2020, 21, 4612. [Google Scholar] [CrossRef]
- De Benedetto, A. Tight junctions in the skin: Still a lot to learn. Br. J. Dermatol. 2021, 184, 388–389. [Google Scholar] [CrossRef]
- Haftek, M. Epidermal Tight Junctions: How Do They Fit into the Barrier Function? Exp. Dermatol. 2010, 19, 1029–1035. [Google Scholar]
- Haftek, M.; Callejon, S.; Sandjeu, Y.; Padois, K.; Falson, F.; Pirot, F.; Portes, P.; Demarne, F.; Jannin, V. Compartmentalization of the human stratum corneum by persistent tight junction-like structures. Exp. Dermatol. 2011, 20, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Igawa, S.; Kishibe, M.; Murakami, M.; Honma, M.; Takahashi, H.; Iizuka, H.; Ishida-Yamamoto, A. Tight junctions in the stratum corneum explain spatial differences in corneodesmosome degradation. Exp. Dermatol. 2011, 20, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Kalinin, A.E.; Kajava, A.V.; Steinert, P.M. Epithelial barrier function: Assembly and structural features of the cornified cell envelope. BioEssays 2002, 24, 789–800. [Google Scholar] [CrossRef]
- Arif, T. Salicylic acid as a peeling agent: A comprehensive review. Clin. Cosmet. Investig. Dermatol. 2015, 8, 455–461. [Google Scholar] [CrossRef]
- Wohlrab, J. Influence of keratolytics on cutaneous pharmacokinetics of glucocorticoids. J. Dtsch. Dermatol. Ges. 2021, 19, 554–561. [Google Scholar] [CrossRef]
- Corcuff, P.; Fiat, F.; Minondo, A.-M.; Lévèque, J.-L.; Rougier, A. A comparative ultrastructural study of hydroxyacids induced desquamation. Eur. J. Dermatol. 2002, 12, XXXIX–XLIII. [Google Scholar]
- He, H.; Bissonnette, R.; Wu, J.; Diaz, A.; Proulx, E.S.-C.; Maari, C.; Jack, C.; Louis, M.; Estrada, Y.; Krueger, J.G.; et al. Tape strips detect distinct immune and barrier profiles in atopic dermatitis and psoriasis. J. Allergy Clin. Immunol. 2021, 147, 199–212. [Google Scholar] [CrossRef]
- De Benedetto, A.; Rafaels, N.M.; McGirt, L.Y.; Ivanov, A.I.; Georas, S.N.; Cheadle, C.; Berger, A.E.; Zhang, K.; Vidyasagar, S.; Yoshida, T.; et al. Tight junction defects in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2011, 127, 773–786.e7. [Google Scholar] [CrossRef]
- Egawa, G.; Kabashima, K. Barrier dysfunction in the skin allergy. Allergol. Int. 2018, 67, 3–11. [Google Scholar] [CrossRef]
- Nakahara, T.; Kido-Nakahara, M.; Tsuji, G.; Furue, M. Basics and recent advances in the pathophysiology of atopic dermatitis. J. Dermatol. 2021, 48, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Medgyesi, B.; Dajnoki, Z.; Béke, G.; Gáspár, K.; Szabó, I.L.; Janka, E.A.; Póliska, S.; Hendrik, Z.; Méhes, G.; Törőcsik, D.; et al. Rosacea Is Characterized by a Profoundly Diminished Skin Barrier. J. Investig. Dermatol. 2020, 140, 1938–1950.e5. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Ma, Y.; Wang, Y.; Wang, X.; Qian, C.; Hocquet, D.; Zheng, S.; Mac-Mary, S.; Humbert, P. Rosacea is associated with conjoined interactions between physical barrier of the skin and microorganisms: A pilot study. J. Clin. Lab. Anal. 2020, 34, e23363. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Chen, M.; Xie, H.; Jian, D.; Xu, S.; Peng, Q.; Sha, K.; Liu, Y.; Zhang, Y.; Shi, W.; et al. Claudin reduction may relate to an impaired skin barrier in rosacea. J. Dermatol. 2019, 46, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Tohgasaki, T.; Ozawa, N.; Yoshino, T.; Ishiwatari, S.; Matsukuma, S.; Yanagi, S.; Fukuda, H. Enolase-1 expression in the stratum corneum is elevated with parakeratosis of atopic dermatitis and disrupts the cellular tight junction barrier in keratinocytes. Int. J. Cosmet. Sci. 2018, 40, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Hadj-Rabia, S.; Baala, L.; Vabres, P.; Hamel-Teillac, D.; Jacquemin, E.; Fabre, M.; Lyonnet, S.; de Prost, Y.; Munnich, A.; Hadchouel, M.; et al. Claudin-1 gene mutations in neonatal sclerosing cholangitis associated with ichthyosis: A tight junction disease. Gastroenterology 2004, 127, 1386–1390. [Google Scholar] [CrossRef] [PubMed]
- Feldmeyer, L.; Huber, M.; Fellmann, F.; Beckmann, J.S.; Frenk, E.; Hohl, D. Confirmation of the origin of NISCH syndrome. Hum. Mutat. 2006, 27, 408–410. [Google Scholar] [CrossRef]
- Oji, V.; Eckl, K.-M.; Aufenvenne, K.; Nätebus, M.; Tarinski, T.; Ackermann, K.; Seller, N.; Metze, D.; Nürnberg, G.; Fölster-Holst, R.; et al. Loss of Corneodesmosin Leads to Severe Skin Barrier Defect, Pruritus, and Atopy: Unraveling the Peeling Skin Disease. Am. J. Hum. Genet. 2010, 87, 274–281. [Google Scholar] [CrossRef]
- Milani, P.; Chlasta, J.; Abdayem, R.; Kezic, S.; Haftek, M. Changes in nano-mechanical properties of human epidermal cornified cells depending on their proximity to the skin surface. J. Mol. Recognit. 2018, 31, e2722. [Google Scholar] [CrossRef]
- Yuki, T.; Komiya, A.; Kusaka, A.; Kuze, T.; Sugiyama, Y.; Inoue, S. Impaired tight junctions obstruct stratum corneum formation by altering polar lipid and profilaggrin processing. J. Dermatol. Sci. 2013, 69, 148–158. [Google Scholar] [CrossRef]
- Bergmann, S.; Von Buenau, B.; Vidal-Y-Sy, S.; Haftek, M.; Wladykowski, E.; Houdek, P.; Lezius, S.; Duplan, H.; Bäsler, K.; Dähnhardt-Pfeiffer, S.; et al. Claudin-1 decrease impacts epidermal barrier function in atopic dermatitis lesions dose-dependently. Sci. Rep. 2020, 10, 2024. [Google Scholar] [CrossRef] [PubMed]
- Niehues, H.; Bouwstra, J.A.; El Ghalbzouri, A.; Brandner, J.M.; Zeeuwen, P.L.J.M.; Van Den Bogaard, E.H. 3D skin models for 3R research: The potential of 3D reconstructed skin models to study skin barrier function. Exp. Dermatol. 2018, 27, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Akagi, T.; Sasano, Y.; Chiba, T.; Narita, H.; Shimoda, H.; Akashi, M. Observation of a tight junction structure generated in LbL-3D skin reconstructed by layer-by-layer cell coating technique. J. Tissue Eng. Regen. Med. 2021, 15, 798–803. [Google Scholar] [CrossRef]
- Brewer, M.G.; Yoshida, T.; Kuo, F.I.; Fridy, S.; Beck, L.A.; De Benedetto, A. Antagonistic Effects of IL-4 on IL-17A-Mediated Enhancement of Epidermal Tight Junction Function. Int. J. Mol. Sci. 2019, 20, 4070. [Google Scholar] [CrossRef] [PubMed]
- Netzlaff, F.; Lehr, C.-M.; Wertz, P.W.; Schaefer, U.F. The human epidermis models EpiSkin®, SkinEthic® and EpiDerm®: An evaluation of morphology and their suitability for testing phototoxicity, irritancy, corrosivity, and substance transport. Eur. J. Pharm. Biopharm. 2005, 60, 167–178. [Google Scholar] [CrossRef]
- Mieremet, A.; Helder, R.; Nadaban, A.; Boiten, W.; Gooris, G.; El Ghalbzouri, A.; Bouwstra, J. Multitargeted Approach for the Optimization of Morphogenesis and Barrier Formation in Human Skin Equivalents. Int. J. Mol. Sci. 2021, 22, 5790. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.E.; Choi, K.J.; Choi, E.H.; Lee, S.H. Acute Modulations in stratum corneum Permeability Barrier Function Affect Claudin Expression and Epidermal Tight Junction Function via Changes of Epidermal Calcium Gradient. Yonsei Med. J. 2013, 54, 523–528. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Menon, G.K.; Lee, S.E.; Lee, S.-H. An overview of epidermal lamellar bodies: Novel roles in biological adaptations and secondary barriers. J. Dermatol. Sci. 2018, 92, 10–17. [Google Scholar] [CrossRef]
- Kurasawa, M.; Maeda, T.; Oba, A.; Yamamoto, T.; Sasaki, H. Tight junction regulates epidermal calcium ion gradient and differentiation. Biochem. Biophys. Res. Commun. 2011, 406, 506–511. [Google Scholar] [CrossRef]
- Kapitány, A.; Medgyesi, B.; Jenei, A.; Somogyi, O.; Szabó, L.; Gáspár, K.; Méhes, G.; Hendrik, Z.; Dócs, K.; Szücs, P.; et al. Regional Differences in the Permeability Barrier of the Skin—Implications in Acantholytic Skin Diseases. Int. J. Mol. Sci. 2021, 22, 10428. [Google Scholar] [CrossRef]
- Zhao, X.; Zeng, H.; Lei, L.; Tong, X.; Yang, L.; Yang, Y.; Li, S.; Zhou, Y.; Luo, L.; Huang, J.; et al. Tight junctions and their regulation by non-coding RNAs. Int. J. Biol. Sci. 2021, 17, 712–727. [Google Scholar] [CrossRef] [PubMed]
- Minowa, E.; Kurashige, Y.; Islam, S.T.; Yoshida, K.; Sakakibara, S.; Okada, Y.; Fujita, Y.; Bolortsetseg, D.; Murai, Y.; Abiko, Y.; et al. Increased integrity of cell–cell junctions accompanied by increased expression of claudin 4 in keratinocytes stimulated with vitamin D3. Med. Mol. Morphol. 2021, 54, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Marras, L.; Caputo, M.; Bisicchia, S.; Soato, M.; Bertolino, G.; Vaccaro, S.; Inturri, R. The Role of Bifidobacteria in Predictive and Preventive Medicine: A Focus on Eczema and Hypercholesterolemia. Microorganisms 2021, 9, 836. [Google Scholar] [CrossRef] [PubMed]
- El-Chami, C.; Foster, A.; Johnson, C.; Clausen, R.; Cornwell, P.; Haslam, I.; Steward, M.; Watson, R.; Young, H.; O’Neill, C. Organic osmolytes increase expression of specific tight junction proteins in skin and alter barrier function in keratinocytes*. Br. J. Dermatol. 2021, 184, 482–494. [Google Scholar] [CrossRef]
- El-Chami, C.; Haslam, I.S.; Steward, M.C.; O’Neill, C.A. Organic osmolytes preserve the function of the developing tight junction in ultraviolet B-irradiated rat epidermal keratinocytes. Sci. Rep. 2018, 8, 5167, Erratum in Sci. Rep. 2020, 10, 8639. [Google Scholar] [CrossRef]
- Fukuta, T.; Tanaka, D.; Inoue, S.; Michiue, K.; Kogure, K. Overcoming thickened pathological skin in psoriasis via iontophoresis combined with tight junction-opening peptide AT1002 for intradermal delivery of NF-κB decoy oligodeoxynucleotide. Int. J. Pharm. 2021, 602, 120601. [Google Scholar] [CrossRef]
- Brewer, M.G.; Anderson, E.A.; Pandya, R.P.; De Benedetto, A.; Yoshida, T.; Hilimire, T.A.; Martinez-Sobrido, L.; Beck, L.A.; Miller, B.L. Peptides Derived from the Tight Junction Protein CLDN1 Disrupt the Skin Barrier and Promote Responsiveness to an Epicutaneous Vaccine. J. Investig. Dermatol. 2020, 140, 361–369.e3. [Google Scholar] [CrossRef]
- Gorzelanny, C.; Mess, C.; Schneider, S.W.; Huck, V.; Brandner, J.M. Skin Barriers in Dermal Drug Delivery: Which Barriers Have to Be Overcome and How Can We Measure Them? Pharmaceutics 2020, 12, 684. [Google Scholar] [CrossRef]
- Finbloom, J.A.; Sousa, F.; Stevens, M.M.; Desai, T.A. Engineering the drug carrier biointerface to overcome biological barriers to drug delivery. Adv. Drug Deliv. Rev. 2020, 167, 89–108. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Skin Source | Additional Pretreatment | % of Apical-Lateral Intercorneocyte Contacts Showing Close Apposition (Fusion) of the Lipid Envelopes (χ² Test) | |
|---|---|---|---|
| Normal skin explants (abdominoplasty; 24 h, 37 °C) | non-occluded (n = 5) | 64.26 |  |
| occluded (n = 5) | 59.14 | ||
| Fresh skin biopsies (arm) | non-occluded (n = 20) | 40.15 ± 6.11 | |
| Vehicle | 1% LSA | (χ² test) | |
|---|---|---|---|
| Skin explants ex vivo (n = 10) | 44% ± 8 | 48% ± 12 | NS |
| Reconstructed epidermis RHE (n = 10) | 57% ± 6 | 70% ± 6 | p < 0.05 |
| SC 1–4 | % “Fused” Apical-Lateral Contacts |
|---|---|
| IHSC patient 1 | 70.4 |
| IHSC patient 2 | 59.8 |
| NHS (n = 35) | 51.5 ± 7.6 |
| SC 1–4 | % “Fused” Lateral-Apical Contacts |
|---|---|
| PSD patient P2 | 79.2 |
| PSD patient P3 | 74.0 |
| NHS (n = 35) | 51.5 ± 7.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haftek, M.; Oji, V.; Feldmeyer, L.; Hohl, D.; Hadj-Rabia, S.; Abdayem, R. The Fate of Epidermal Tight Junctions in the stratum corneum: Their Involvement in the Regulation of Desquamation and Phenotypic Expression of Certain Skin Conditions. Int. J. Mol. Sci. 2022, 23, 7486. https://doi.org/10.3390/ijms23137486
Haftek M, Oji V, Feldmeyer L, Hohl D, Hadj-Rabia S, Abdayem R. The Fate of Epidermal Tight Junctions in the stratum corneum: Their Involvement in the Regulation of Desquamation and Phenotypic Expression of Certain Skin Conditions. International Journal of Molecular Sciences. 2022; 23(13):7486. https://doi.org/10.3390/ijms23137486
Chicago/Turabian StyleHaftek, Marek, Vinzenz Oji, Laurence Feldmeyer, Daniel Hohl, Smaïl Hadj-Rabia, and Rawad Abdayem. 2022. "The Fate of Epidermal Tight Junctions in the stratum corneum: Their Involvement in the Regulation of Desquamation and Phenotypic Expression of Certain Skin Conditions" International Journal of Molecular Sciences 23, no. 13: 7486. https://doi.org/10.3390/ijms23137486
APA StyleHaftek, M., Oji, V., Feldmeyer, L., Hohl, D., Hadj-Rabia, S., & Abdayem, R. (2022). The Fate of Epidermal Tight Junctions in the stratum corneum: Their Involvement in the Regulation of Desquamation and Phenotypic Expression of Certain Skin Conditions. International Journal of Molecular Sciences, 23(13), 7486. https://doi.org/10.3390/ijms23137486