
Altered Balance of Reelin Proteolytic Fragments in the Cerebrospinal Fluid of Alzheimer’s Disease Patients


Abstract
:1. Introduction
2. Results
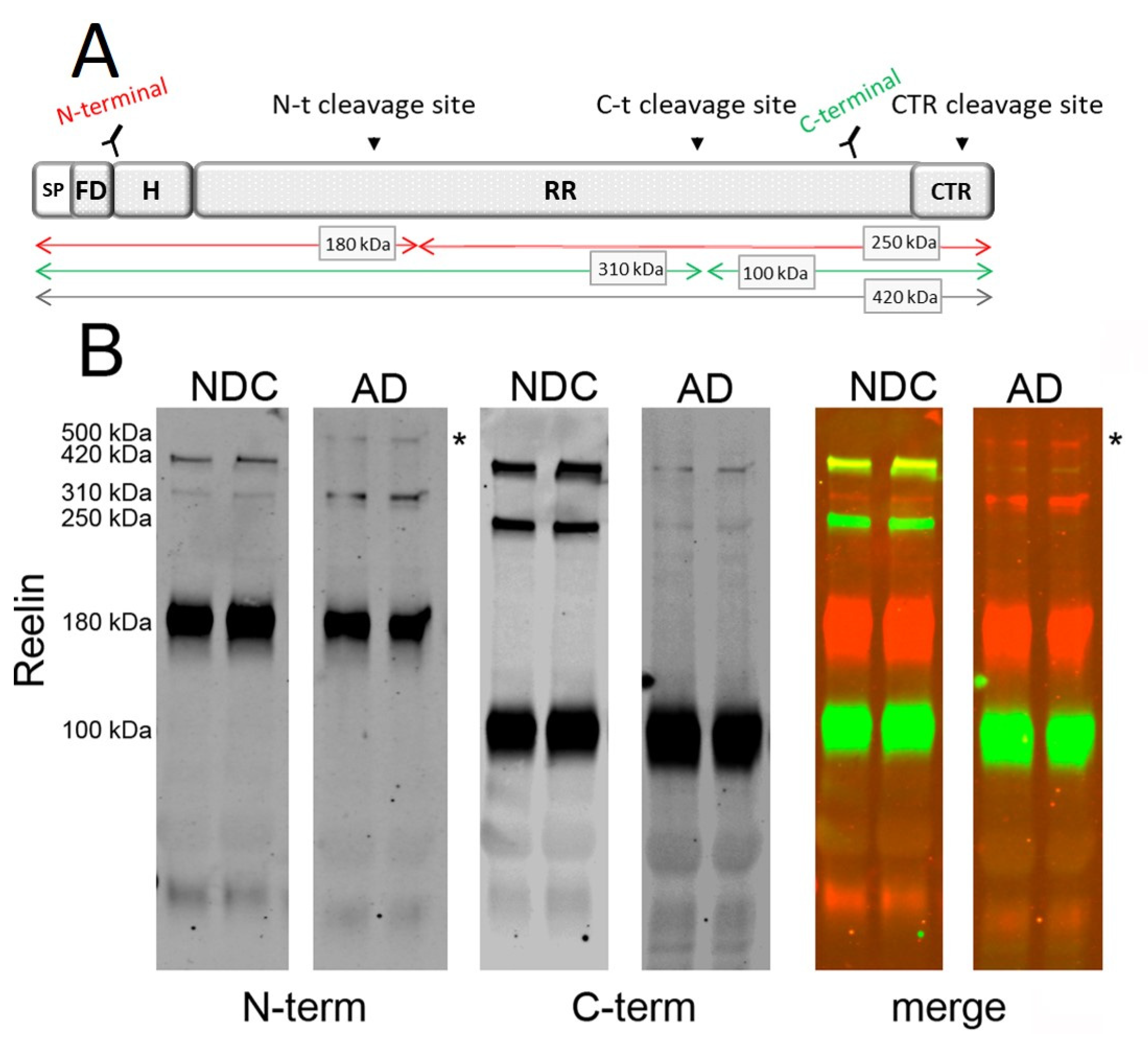
2.1. Characterization of Reelin Species in AD CSF
2.2. Determination of Reelin Species in AD CSF
2.3. Occurrence of the 500 kDa Reelin Species in Culture Media of Aβ42-Treated Cells
2.4. Determination of CSF apoER2 in AD Subjects Subgrouped by APOE Genotype
3. Discussion
4. Materials and Methods
4.1. Human CSF Samples
4.2. Cell Culture and Aβ Treatment
4.3. Western Blot
4.4. Measurement of T-tau and Aβ42 by ELISA
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Scheltens, P.; de Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Ranson, J.M.; Rittman, T.; Hayat, S.; Brayne, C.; Jessen, F.; Blennow, K.; van Duijn, C.; Barkhof, F.; Tang, E.; Mummery, C.J.; et al. Modifiable risk factors for dementia and dementia risk profiling. A user manual for Brain Health Services-part 2 of 6. Alzheimers Res. Ther. 2021, 13, 169. [Google Scholar] [CrossRef]
- Rabaneda-Bueno, R.; Mena-Montes, B.; Torres-Castro, S.; Torres-Carrillo, N.; Torres-Carrillo, N.M. Advances in Genetics and Epigenetic Alterations in Alzheimer's Disease: A Notion for Therapeutic Treatment. Genes 2021, 12, 1959. [Google Scholar]
- Panpalli, A.M.; Karaman, Y.; Guntekin, S.; Ergun, M.A. Analysis of genetics and risk factors of Alzheimer’s Disease. Neuroscience 2016, 325, 124–131. [Google Scholar] [CrossRef]
- Zhao, N.; Liu, C.C.; Qiao, W.; Bu, G. Apolipoprotein, E, receptors, and modulation of Alzheimer’s Disease. Biol. Psychiatry 2018, 83, 347–357. [Google Scholar] [CrossRef]
- Corder, E.H.; Saunders, A.M.; Strittmatter, W.J.; Schmechel, D.E.; Gaskell, P.C.; Small, G.W.; Roses, A.D.; Haines, J.L.; Pericak-Vance, M.A. Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer’s disease in late onset families. Science 1993, 261, 921–923. [Google Scholar] [CrossRef]
- Holtzman, D.M.; Herz, J.; Bu, G. Apolipoprotein E and apolipoprotein E receptors: Normal biology and roles in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006312. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, E.; Paudel, Y.N.; Papageorgiou, S.G.; Piperi, C. APOE Genotype and Alzheimer’s Disease: The Influence of Lifestyle and Environmental Factors. ACS Chem. Neurosci. 2021, 12, 2749–2764. [Google Scholar] [CrossRef]
- Dyer, C.A.; Smith, R.S.; Curtiss, L.K. Only multimers of a synthetic peptide of human apolipoprotein E are biologically active. J. Biol. Chem. 1991, 266, 15009–15015. [Google Scholar] [CrossRef]
- DeSilva, U.; D’Arcangelo, G.; Braden, V.V.; Chen, J.; Miao, G.G.; Curran, T.; Green, E.D. The human reelin gene: Isolation, sequencing and mapping on chromosome 7. Genome Res. 1997, 7, 157–164. [Google Scholar] [CrossRef]
- D’Arcangelo, G.; Homayouni, R.; Keshvara, L.; Rice, D.S.; Sheldon, M.; Curran, T. Reelin is a ligand for lipoprotein receptors. Neuron 1999, 24, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Hiesberger, T.; Trommsdorff, M.; Howell, B.W.; Goffinet, A.; Mumby, M.C.; Cooper, J.A.; Herz, J. Direct binding of Reelin to VLDL receptor and ApoE receptor 2 induces tyrosine phosphorylation of disabled-1 and modulates tau phosphorylation. Neuron 1999, 24, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Tissir, F.; Goffinet, A.M. Reelin and brain development. Nat. Rev. Neurosci. 2003, 4, 496–505. [Google Scholar] [CrossRef]
- Chai, X.; Frotscher, M. How does Reelin signaling regulate the neuronal cytoskeleton during migration? Neurogenesis 2016, 3, e1242455. [Google Scholar] [CrossRef] [Green Version]
- Lane-Donovan, C.; Herz, J. The ApoE receptors Vldlr and Apoer2 in central nervous system function and disease. J. Lipid Res. 2017, 58, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Sáez-Valero, J.; Costell, M.; Sjogren, M.; Andreasen, N.; Blennow, K.; Luque, J.M. Altered levels of cerebrospinal fluid reelin in frontotemporal dementia and Alzheimer’s disease. J. Neurosci. Res. 2003, 72, 132–136. [Google Scholar] [CrossRef]
- Botella-Lopez, A.; Burgaya, F.; Gavin, R.; Garcia-Ayllon, M.S.; Gomez-Tortosa, E.; Peña-Casanova, J.; Urena, J.M.; del Rio, J.A.; Blesa, R.; Soriano, E.; et al. Reelin expression and glycosylation patterns are altered in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2006, 103, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Cuchillo-Ibañez, I.; Mata-Balaguer, T.; Balmaceda, V.; Arranz, J.J.; Nimpf, J.; Sáez-Valero, J. The beta-amyloid peptide compromises Reelin signaling in Alzheimer’s disease. Sci. Rep. 2016, 6, 31646. [Google Scholar] [CrossRef] [Green Version]
- Cuchillo-Ibañez, I.; Balmaceda, V.; Botella-Lopez, A.; Rabano, A.; Avila, J.; Sáez-Valero, J. Beta-amyloid impairs reelin signaling. PLoS ONE 2013, 8, e72297. [Google Scholar] [CrossRef] [Green Version]
- Kubo, K.; Mikoshiba, K.; Nakajima, K. Secreted Reelin molecules form homodimers. Neurosci. Res. 2002, 43, 381–388. [Google Scholar] [CrossRef]
- Yasui, N.; Kitago, Y.; Beppu, A.; Kohno, T.; Morishita, S.; Gomi, H.; Nagae, M.; Hattori, M.; Takagi, J. Functional importance of covalent homodimer of reelin protein linked via its central region. J. Biol. Chem. 2011, 286, 35247–35256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dlugosz, P.; Tresky, R.; Nimpf, J. Differential Action of Reelin on Oligomerization of ApoER2 and VLDL Receptor in HEK293 Cells Assessed by Time-Resolved Anisotropy and Fluorescence Lifetime Imaging Microscopy. Front. Mol. Neurosci. 2019, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Jossin, Y.; Gui, L.; Goffinet, A.M. Processing of Reelin by embryonic neurons is important for function in tissue but not in dissociated cultured neurons. J. Neurosci. 2007, 27, 4243–4252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoe, H.S.; Cooper, M.J.; Burns, M.P.; Lewis, P.A.; van der Brug, M.; Chakraborty, G.; Cartagena, C.M.; Pak, D.T.; Cookson, M.R.; Rebeck, G.W. The metalloprotease inhibitor TIMP-3 regulates amyloid precursor protein and apolipoprotein E receptor proteolysis. J. Neurosci. 2007, 27, 10895–10905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibi, T.; Hattori, M. The N-terminal fragment of Reelin is generated after endocytosis and released through the pathway regulated by Rab11. FEBS Lett. 2009, 583, 1299–1303. [Google Scholar] [CrossRef] [Green Version]
- Balmaceda, V.; Cuchillo-Ibáñez, I.; Pujadas, L.; García-Ayllón, M.S.; Saura, C.A.; Nimpf, J.; Soriano, E.; Sáez-Valero, J. ApoER2 processing by presenilin-1 modulates reelin expression. FASEB J. 2014, 28, 1543–1554. [Google Scholar] [CrossRef] [Green Version]
- Hattori, M.; Kohno, T. Regulation of Reelin functions by specific proteolytic processing in the brain. J. Biochem. 2021, 169, 511–516. [Google Scholar] [CrossRef]
- Turk, L.S.; Kuang, X.; Dal Pozzo, V.; Patel, K.; Chen, M.; Huynh, K.; Currie, M.J.; Mitchell, D.; Dobson, R.C.J.; D’Arcangelo, G.; et al. The structure-function relationship of a signaling-competent; dimeric Reelin fragment. Structure 2021, 29, 1156–1170. [Google Scholar] [CrossRef]
- Koch, S.; Strasser, V.; Hauser, C.; Fasching, D.; Brandes, C.; Bajari, T.M.; Schneider, W.J.; Nimpf, J. A secreted soluble form of ApoE receptor 2 acts as a dominant-negative receptor and inhibits Reelin signaling. EMBO J. 2002, 21, 5996–6004. [Google Scholar] [CrossRef] [Green Version]
- Smalheiser, N.R.; Costa, E.; Guidotti, A.; Impagnatiello, F.; Auta, J.; Lacor, P.; Kriho, V.; Pappas, G.D. Expression of reelin in adult mammalian blood; liver; pituitary pars intermedia; and adrenal chromaffin cells. Proc. Natl. Acad. Sci. USA 2000, 97, 1281–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohno, T.; Honda, T.; Kubo, K.; Nakano, Y.; Tsuchiya, A.; Murakami, T.; Banno, H.; Nakajima, K.; Hattori, M. Importance of Reelin C-terminal region in the development and maintenance of the postnatal cerebral cortex and its regulation by specific proteolysis. J. Neurosci. 2015, 35, 4776–4787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ignatova, N.; Sindic, C.J.; Goffinet, A.M. Characterization of the various forms of the Reelin protein in the cerebrospinal fluid of normal subjects and in neurological diseases. Neurobiol. Dis. 2004, 15, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Lugli, G.; Krueger, J.M.; Davis, J.M.; Persico, A.M.; Keller, F.; Smalheiser, N.R. Methodological factors influencing measurement and processing of plasma reelin in humans. BMC Biochem. 2003, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef] [PubMed]
- Hansson, O.; Zetterberg, H.; Buchhave, P.; Londos, E.; Blennow, K.; Minthon, L. Association between CSF biomarkers and incipient Alzheimer’s disease in patients with mild cognitive impairment: A follow-up study. Lancet Neurol. 2006, 5, 228–234. [Google Scholar] [CrossRef] [Green Version]
- Strasser, V.; Fasching, D.; Hauser, C.; Mayer, H.; Bock, H.H.; Hiesberger, T.; Herz, J.; Weeber, E.J.; Sweatt, J.D.; Pramatarova, A.; et al. Receptor clustering is involved in Reelin signaling. Mol. Cell Biol. 2004, 24, 1378–1386. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Font, I.; Iborra-Lazaro, G.; Sanchez-Valle, R.; Molinuevo, J.L.; Cuchillo-Ibañez, I.; Sáez-Valero, J. CSF-ApoER2 fragments as a read-out of reelin signaling: Distinct patterns in sporadic and autosomal-dominant Alzheimer disease. Clin. Chim. Acta 2019, 490, 6–11. [Google Scholar] [CrossRef]
- Botella-Lopez, A.; Cuchillo-Ibañez, I.; Cotrufo, T.; Mok, S.S.; Li, Q.X.; Barquero, M.S.; Dierssen, M.; Soriano, E.; Sáez-Valero, J. Beta-amyloid controls altered Reelin expression and processing in Alzheimer’s disease. Neurobiol. Dis. 2010, 37, 682–691. [Google Scholar] [CrossRef] [Green Version]
- Mata-Balaguer, T.; Cuchillo-Ibañez, I.; Calero, M.; Ferrer, I.; Sáez-Valero, J. Decreased generation of C-terminal fragments of ApoER2 and increased reelin expression in Alzheimer’s disease. FASEB J. 2018, 32, 3536–3546. [Google Scholar] [CrossRef] [Green Version]
- Wirths, O.; Multhaup, G.; Czech, C.; Blanchard, V.; Tremp, G.; Pradier, L.; Beyreuther, K.; Bayer, T.A. Reelin in plaques of beta-amyloid precursor protein and presenilin-1 double-transgenic mice. Neurosci. Lett. 2001, 316, 145–148. [Google Scholar] [CrossRef]
- Chin, J.; Massaro, C.M.; Palop, J.J.; Thwin, M.T.; Yu, G.Q.; Bien-Ly, N.; Bender, A.; Mucke, L. Reelin depletion in the entorhinal cortex of human amyloid precursor protein transgenic mice and humans with Alzheimer’s disease. J. Neurosci. 2007, 27, 2727–2733. [Google Scholar] [CrossRef]
- Herring, A.; Donath, A.; Steiner, K.M.; Widera, M.P.; Hamzehian, S.; Kanakis, D.; Kolble, K.; ElAli, A.; Hermann, D.M.; Paulus, W.; et al. Reelin depletion is an early phenomenon of Alzheimer’s pathology. J. Alzheimers Dis. 2012, 30, 963–979. [Google Scholar] [CrossRef] [PubMed]
- Cuchillo-Ibañez, I.; Balmaceda, V.; Mata-Balaguer, T.; Lopez-Font, I.; Sáez-Valero, J. Reelin in Alzheimer’s Disease; Increased Levels but Impaired Signaling: When More is Less. J. Alzheimers Dis. 2016, 52, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Mathews, S.T.; Plaisance, E.P.; Kim, T. Imaging systems for westerns: Chemiluminescence vs. infrared detection. Methods Mol. Biol. 2009, 536, 499–513. [Google Scholar] [PubMed]
- Gingrich, J.C.; Davis, D.R.; Nguyen, Q. Multiplex detection and quantitation of proteins on western blots using fluorescent probes. Biotechniques 2000, 29, 636–642. [Google Scholar] [CrossRef] [Green Version]
- Ogino, H.; Hisanaga, A.; Kohno, T.; Kondo, Y.; Okumura, K.; Kamei, T.; Sato, T.; Asahara, H.; Tsuiji, H.; Fukata, M.; et al. Secreted Metalloproteinase ADAMTS-3 Inactivates Reelin. J. Neurosci. 2017, 37, 3181–3191. [Google Scholar] [CrossRef]
- Jossin, Y. Reelin Functions; Mechanisms of Action and Signaling Pathways During Brain Development and Maturation. Biomolecules 2020, 10, 964. [Google Scholar] [CrossRef]
- Krstic, D.; Rodriguez, M.; Knuesel, I. Regulated proteolytic processing of Reelin through interplay of tissue plasminogen activator (tPA), ADAMTS-4, ADAMTS-5 and their modulators. PLoS ONE 2012, 7, e47793. [Google Scholar] [CrossRef]
- Tsuneura, Y.; Sawahata, M.; Itoh, N.; Miyajima, R.; Mori, D.; Kohno, T.; Hattori, M.; Sobue, A.; Nagai, T.; Mizoguchi, H.; et al. Analysis of Reelin signaling and neurodevelopmental trajectory in primary cultured cortical neurons with RELN deletion identified in schizophrenia. Neurochem. Int. 2021, 144, 104954. [Google Scholar] [CrossRef]
- Jefferson, T.; Čausević, M.; auf dem Keller, U.; Schilling, O.; Isbert, S.; Geyer, R.; Maier, W.; Tschickardt, S.; Jumpertz, T.; Weggen, S.; et al. Metalloprotease meprin beta generates nontoxic N-terminal amyloid precursor protein fragments in vivo. J. Biol. Chem. 2011, 286, 27741–27750. [Google Scholar] [CrossRef] [Green Version]
- Bien, J.; Jefferson, T.; Causevic, M.; Jumpertz, T.; Munter, L.; Multhaup, G.; Weggen, S.; Becker-Pauly, C.; Pietrzik, C.U. The metalloprotease meprin beta generates amino terminal-truncated amyloid beta peptide species. J. Biol. Chem. 2012, 287, 33304–33313. [Google Scholar] [CrossRef] [Green Version]
- Medoro, A.; Bartollino, S.; Mignogna, D.; Marziliano, N.; Porcile, C.; Nizzari, M.; Florio, T.; Pagano, A.; Raimo, G.; Intrieri, M.; et al. Proteases Upregulation in Sporadic Alzheimer’s Disease Brain. J. Alzheimers Dis. 2019, 68, 931–938. [Google Scholar] [CrossRef]
- Angelucci, F.; Cechova, K.; Prusa, R.; Hort, J. Amyloid beta soluble forms and plasminogen activation system in Alzheimer’s disease: Consequences on extracellular maturation of brain-derived neurotrophic factor and therapeutic implications. CNS Neurosci. Ther. 2019, 25, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Hanzel, C.E.; Iulita, M.F.; Eyjolfsdottir, H.; Hjorth, E.; Schultzberg, M.; Eriksdotter, M.; Cuello, A.C. Analysis of matrix metallo-proteases and the plasminogen system in mild cognitive impairment and Alzheimer’s disease cerebrospinal fluid. J. Alzheimers Dis. 2014, 40, 667–678. [Google Scholar] [CrossRef]
- Whelan, C.D.; Mattsson, N.; Nagle, M.W.; Vijayaraghavan, S.; Hyde, C.; Janelidze, S.; Stomrud, E.; Lee, J.; Fitz, L.; Samad, T.A.; et al. Multiplex proteomics identifies novel CSF and plasma biomarkers of early Alzheimer’s disease. Acta Neuropathol. Commun. 2019, 7, 169. [Google Scholar] [CrossRef]
- García-Ayllón, M.S.; Campanari, M.L.; Brinkmalm, G.; Rabano, A.; Alom, J.; Saura, C.A.; Andreasen, N.; Blennow, K.; Sáez-Valero, J. CSF Presenilin-1 complexes are increased in Alzheimer’s disease. Acta Neuropathol. Commun. 2013, 1, 46. [Google Scholar] [CrossRef] [Green Version]
- Sogorb-Esteve, A.; Garcia-Ayllon, M.S.; Fortea, J.; Sanchez-Valle, R.; Lleo, A.; Molinuevo, J.L.; Sáez-Valero, J. Cerebrospinal fluid Presenilin-1 increases at asymptomatic stage in genetically determined Alzheimer’s disease. Mol. Neurodegener. 2016, 11, 66. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Nordberg, A.; Darreh-Shori, T. Amyloid-beta peptides act as allosteric modulators of cholinergic signalling through formation of soluble BAbetaACs. Brain 2016, 139, 174–192. [Google Scholar] [CrossRef] [Green Version]
- Kanekiyo, T.; Xu, H.; Bu, G. ApoE and Abeta in Alzheimer’s disease: Accidental encounters or partners? Neuron 2014, 81, 740–754. [Google Scholar] [CrossRef] [Green Version]
- Tai, L.M.; Mehra, S.; Shete, V.; Estus, S.; Rebeck, G.W.; Bu, G.; LaDu, M.J. Soluble apoE/Abeta complex: Mechanism and therapeutic target for APOE4-induced AD risk. Mol. Neurodegener. 2014, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Pujadas, L.; Rossi, D.; Andres, R.; Teixeira, C.M.; Serra-Vidal, B.; Parcerisas, A.; Maldonado, R.; Giralt, E.; Carulla, N.; Soriano, E. Reelin delays amyloid-beta fibril formation and rescues cognitive deficits in a model of Alzheimer’s disease. Nat. Commun. 2014, 5, 3443. [Google Scholar] [CrossRef] [PubMed]
- Beffert, U.; Weeber, E.J.; Durudas, A.; Qiu, S.; Masiulis, I.; Sweatt, J.D.; Li, W.P.; Adelmann, G.; Frotscher, M.; Hammer, R.E.; et al. Modulation of synaptic plasticity and memory by Reelin involves differential splicing of the lipoprotein receptor Apoer2. Neuron 2005, 47, 567–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Cooley, K.; Chung, C.H.; Dashti, N.; Tang, J. Apolipoprotein receptor 2 and X11 alpha/beta mediate apolipoprotein E-induced endocytosis of amyloid-beta precursor protein and beta-secretase; leading to amyloid-beta production. J. Neurosci. 2007, 27, 4052–4060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Moerman-Herzog, A.M.; Slaton, A.; Barger, S.W. Presenilin 1 mutations influence processing and trafficking of the ApoE receptor apoER2. Neurobiol. Aging 2017, 49, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.J.; Cao, F.; Wu, Y.; Peng, J.H.; Zhong, J.J.; Jiang, Y.; Yin, C.; Guo, Z.D.; Sun, X.C.; Jiang, L.; et al. Apolipoprotein E promotes white matter remodeling via the Dab1-dependent pathway after traumatic brain injury. CNS Neurosci. Ther. 2020, 26, 698–710. [Google Scholar] [CrossRef] [Green Version]
- Xian, X.; Pohlkamp, T.; Durakoglugil, M.S.; Wong, C.H.; Beck, J.K.; Lane-Donovan, C.; Plattner, F.; Herz, J. Reversal of ApoE4-induced recycling block as a novel prevention approach for Alzheimer’s disease. eLife 2018, 30, e40048. [Google Scholar] [CrossRef]
- Divekar, S.D.; Burrell, T.C.; Lee, J.E.; Weeber, E.J.; Rebeck, G.W. Ligand-induced homotypic and heterotypic clustering of apolipoprotein E receptor 2. J. Biol. Chem. 2014, 289, 15894–15903. [Google Scholar] [CrossRef] [Green Version]
- Weisgraber, K.H.; Shinto, L.H. Identification of the disulfide-linked homodimer of apolipoprotein E3 in plasma. Impact on receptor binding activity. J. Biol. Chem. 1991, 266, 12029–12034. [Google Scholar] [CrossRef]
- Chen, Y.; Durakoglugil, M.S.; Xian, X.; Herz, J. ApoE4 reduces glutamate receptor function and synaptic plasticity by selectively impairing ApoE receptor recycling. Proc. Natl. Acad. Sci. USA 2010, 107, 12011–12016. [Google Scholar] [CrossRef] [Green Version]
- Ringland, C.; Schweig, J.E.; Paris, D.; Shackleton, B.; Lynch, C.E.; Eisenbaum, M.; Mullan, M.; Crawford, F.; Abdullah, L.; Bachmeier, C. Apolipoprotein E isoforms differentially regulate matrix metallopeptidase 9 function in Alzheimer’s disease. Neurobiol. Aging 2020, 95, 56–68. [Google Scholar] [CrossRef]
- Hisanaga, A.; Morishita, S.; Suzuki, K.; Sasaki, K.; Koie, M.; Kohno, T.; Hattori, M. A disintegrin and metalloproteinase with thrombospondin motifs 4 (ADAMTS-4) cleaves Reelin in an isoform-dependent manner. FEBS Lett. 2012, 586, 3349–3353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane-Donovan, C.; Philips, G.T.; Wasser, C.R.; Durakoglugil, M.S.; Masiulis, I.; Upadhaya, A.; Pohlkamp, T.; Coskun, C.; Kotti, T.; Steller, L.; et al. Reelin protects against amyloid β toxicity in vivo. Sci. Signal. 2015, 8, ra67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durakoglugil, M.S.; Chen, Y.; White, C.L.; Kavalali, E.T.; Herz, J. Reelin signaling antagonizes beta-amyloid at the synapse. Proc. Natl. Acad. Sci. USA 2009, 106, 15938–15943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuneura, Y.; Nakai, T.; Mizoguchi, H.; Yamada, K. New Strategies for the Treatment of Neuropsychiatric Disorders Based on Reelin Dysfunction. Int. J. Mol. Sci. 2022, 23, 1829. [Google Scholar] [CrossRef]
- Rogers, J.T.; Rusiana, I.; Trotter, J.; Zhao, L.; Donaldson, E.; Pak, D.T.; Babus, L.W.; Peters, M.; Banko, J.L.; Chavis, P.; et al. Reelin supplementation enhances cognitive ability, synaptic plasticity, and dendritic spine density. Learn. Mem. 2011, 18, 558–564. [Google Scholar] [CrossRef] [Green Version]
- Blennow, K.; Ricksten, A.; Prince, J.A.; Brookes, A.J.; Emahazion, T.; Wasslavik, C.; Bogdanovic, N.; Andreasen, N.; Batsman, S.; Marcusson, J.; et al. No association between the alpha2-macroglobulin (A2M) deletion and Alzheimer’s disease; and no change in A2M mRNA, protein or protein expression. J. Neural Transm. 2000, 107, 1065–1079. [Google Scholar] [CrossRef]
- Palmqvist, S.; Zetterberg, H.; Blennow, K.; Vestberg, S.; Andreasson, U.; Brooks, D.J.; Owenius, R.; Hagerstrom, D.; Wollmer, P.; Minthon, L.; et al. Accuracy of brain amyloid detection in clinical practice using cerebrospinal fluid beta-amyloid 42: A cross-validation study against amyloid positron emission tomography. JAMA Neurol. 2014, 71, 1282–1289. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CSF Cohort | |||||||
|---|---|---|---|---|---|---|---|
| Control | Alzheimer’s Disease | ||||||
| APOE | ε3/3 | ε3/4 | All | ε3/3 | ε3/4 | ε4/4 | All |
| N | 9 | 5 | 14 | 15 | 13 | 15 | 43 |
| Age (Years) | 69 ± 2 | 62 ± 5 | 67 ± 3 | 79 ± 2 | 78 ± 1 | 73 ± 1 | 77 ± 1 * |
| Age (Range) | 60–81 | 44–75 | 44–81 | 62–88 | 69–84 | 63–83 | 62–88 |
| Female/Male | 5/4 | 2/3 | 7/7 | 11/4 | 10/3 | 9/6 | 31/14 |
| CSF Aβ42 (pg/mL) | 845 ± 96 | 746 ± 121 | 804 ± 74 | 470 ± 13 * | 484 ± 9 * | 419 ± 21 | 457 ± 10 * |
| CSF Total Tau (pg/mL) | 317 ± 53 | 303 ± 34 | 312 ± 35 | 816 ± 88 * | 1004 ± 127 * | 731 ± 53 | 840 ± 52 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Font, I.; Lennol, M.P.; Iborra-Lazaro, G.; Zetterberg, H.; Blennow, K.; Sáez-Valero, J. Altered Balance of Reelin Proteolytic Fragments in the Cerebrospinal Fluid of Alzheimer’s Disease Patients. Int. J. Mol. Sci. 2022, 23, 7522. https://doi.org/10.3390/ijms23147522
Lopez-Font I, Lennol MP, Iborra-Lazaro G, Zetterberg H, Blennow K, Sáez-Valero J. Altered Balance of Reelin Proteolytic Fragments in the Cerebrospinal Fluid of Alzheimer’s Disease Patients. International Journal of Molecular Sciences. 2022; 23(14):7522. https://doi.org/10.3390/ijms23147522
Chicago/Turabian StyleLopez-Font, Inmaculada, Matthew P. Lennol, Guillermo Iborra-Lazaro, Henrik Zetterberg, Kaj Blennow, and Javier Sáez-Valero. 2022. "Altered Balance of Reelin Proteolytic Fragments in the Cerebrospinal Fluid of Alzheimer’s Disease Patients" International Journal of Molecular Sciences 23, no. 14: 7522. https://doi.org/10.3390/ijms23147522