
Knockdown of VEGFB/VEGFR1 Signaling Promotes White Adipose Tissue Browning and Skeletal Muscle Development

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
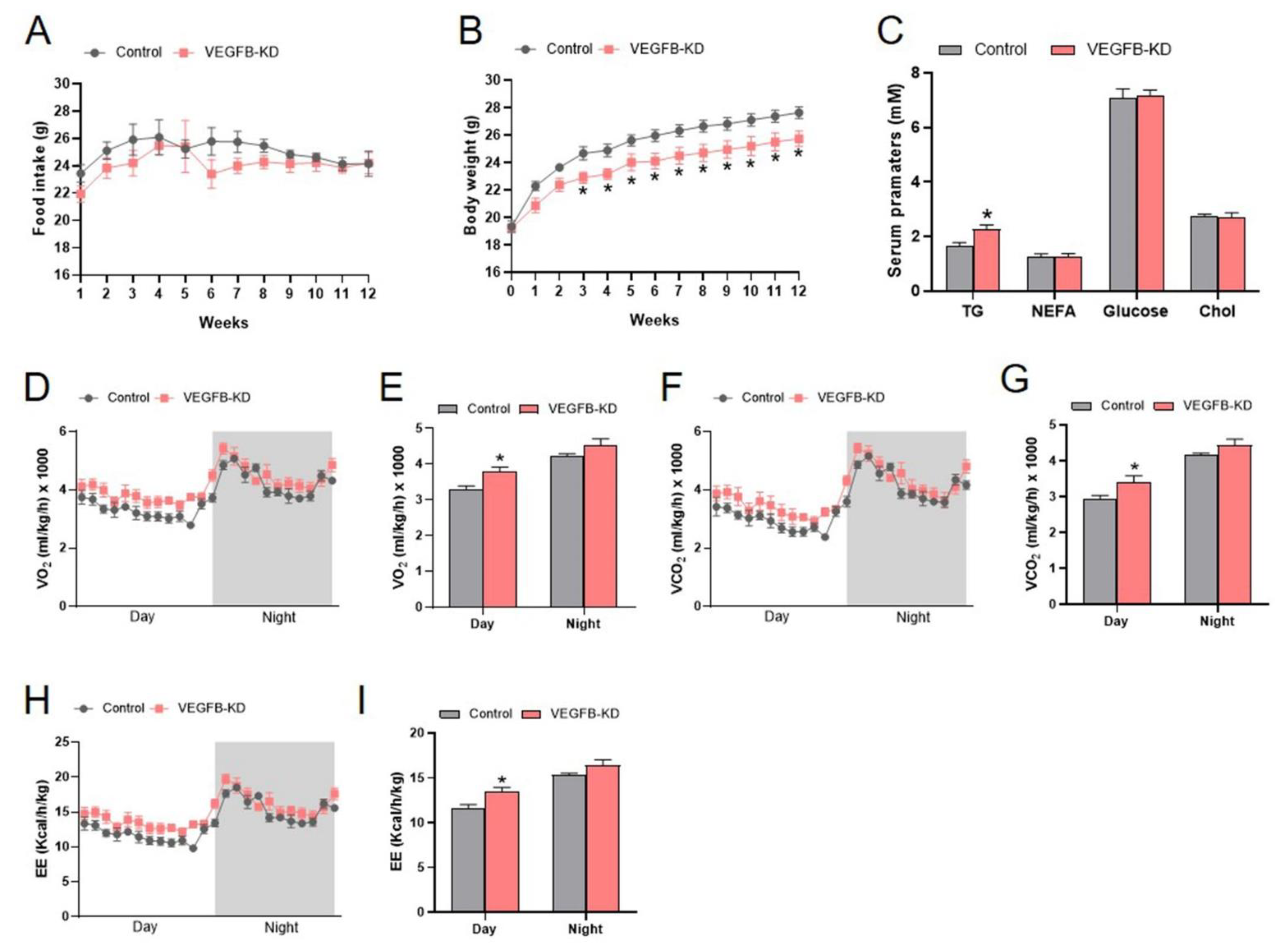
2.1. Knockdown of VEGFB Decreased Body Weight and Increased Energy Expenditure of Mice
2.2. Knockdown of VEGFB Stimulated the Browning of iWAT in Mice
2.3. Knockdown of VEGFB Increased Gastrocnemius Muscle Index with Elevated Proliferation in Mice
2.4. Knockdown Effect of Endothelial VEGFR1 on the Growth and Energy Expenditure of Mice
2.5. Knockdown of Endothelial VEGFR1 Stimulated the Browning of iWAT in Mice
2.6. Knockdown of Endothelial VEGFR1 Did Not Affect GAS Muscle Index but Inhibited Its Differentiation
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Animals
4.3. Adeno-Associated Viral Vector (AAV) Injection
4.4. Metabolic Measures
4.5. Locomotor Activity
4.6. Blood Parameters
4.7. Glucose and Insulin Tolerance Tests
4.8. Hematoxylin and Eosin (H&E) Staining
4.9. Real-Time Quantitative PCR (RT-PCR) Analysis
4.10. Western Blotting Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal muscle atrophy: From mechanisms to treatments. Pharmacol. Res. 2021, 172, 105807. [Google Scholar] [CrossRef]
- Tardif, N.; Salles, J.; Guillet, C.; Tordjman, J.; Reggio, S.; Landrier, J.F.; Giraudet, C.; Patrac, V.; Bertrand-Michel, J.; Migne, C.; et al. Muscle ectopic fat deposition contributes to anabolic resistance in obese sarcopenic old rats through eIF2alpha activation. Aging. Cell 2014, 13, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking adipogenesis during white adipose tissue development, expansion and regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar] [CrossRef]
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008417. [Google Scholar] [CrossRef] [Green Version]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Alipoor, E.; Hosseinzadeh-Attar, M.J.; Rezaei, M.; Jazayeri, S.; Chapman, M. White adipose tissue browning in critical illness: A review of the evidence, mechanisms and future perspectives. Obes Rev. 2020, 21, e13085. [Google Scholar] [CrossRef]
- Kaisanlahti, A.; Glumoff, T. Browning of white fat: Agents and implications for beige adipose tissue to type 2 diabetes. J. Physiol. Biochem. 2019, 75, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mu, W.; Qian, S.; Song, Y.; Yang, L.; Song, S.; Yang, Q.; Liu, H.; Liu, Y.; Pan, D.; Tang, Y.; et al. BMP4-mediated browning of perivascular adipose tissue governs an anti-inflammatory program and prevents atherosclerosis. Redox. Biol. 2021, 43, 101979. [Google Scholar] [CrossRef]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.; Tanaka, T.; Mulati, M.; Ochi, H.; Sato, S.; Kaldis, P.; Yoshii, T.; Okawa, A.; Inose, H. Cyclin-Dependent Kinase 1 Is Essential for Muscle Regeneration and Overload Muscle Fiber Hypertrophy. Front. Cell Dev. Biol. 2020, 8, 564581. [Google Scholar] [CrossRef] [PubMed]
- Ewen, M.E. Where the cell cycle and histones meet. Genes Dev. 2000, 14, 2265–2270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol 2011, 12, 349–361. [Google Scholar] [CrossRef]
- Devlin, R.B.; Emerson, C.P., Jr. Coordinate regulation of contractile protein synthesis during myoblast differentiation. Cell 1978, 13, 599–611. [Google Scholar] [CrossRef]
- Yanagiuchi, A.; Miyake, H.; Nomi, M.; Takenaka, A.; Fujisawa, M. Modulation of the microenvironment by growth factors regulates the in vivo growth of skeletal myoblasts. BJU Int. 2009, 103, 1569–1573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Song, M.; Chen, L.; Yang, X.; Li, F.; Yang, Q.; Duan, C.; Ling, M.; Lai, X.; Zhu, X.; et al. Dietary Supplementation of Lauric Acid Alleviates the Irregular Estrous Cycle and the Impaired Metabolism and Thermogenesis in Female Mice Fed with High-Fat Diet (HFD). J. Agric. Food Chem. 2020, 68, 12631–12640. [Google Scholar] [CrossRef]
- Duan, C.; Yin, C.; Ma, Z.; Li, F.; Zhang, F.; Yang, Q.; Lin, M.; Feng, S.; Zhu, C.; Wang, L.; et al. trans 10, cis 12, but Not cis 9, trans 11 Conjugated Linoleic Acid Isomer Enhances Exercise Endurance by Increasing Oxidative Skeletal Muscle Fiber Type via Toll-like Receptor 4 Signaling in Mice. J. Agric. Food Chem. 2021, 69, 15636–15648. [Google Scholar] [CrossRef]
- Huey, K.A. Potential Roles of Vascular Endothelial Growth Factor During Skeletal Muscle Hypertrophy. Exerc. Sport Sci. Rev. 2018, 46, 195–202. [Google Scholar] [CrossRef]
- Robciuc, M.R.; Kivela, R.; Williams, I.M.; de Boer, J.F.; van Dijk, T.H.; Elamaa, H.; Tigistu-Sahle, F.; Molotkov, D.; Leppanen, V.M.; Kakela, R.; et al. VEGFB/VEGFR1-Induced Expansion of Adipose Vasculature Counteracts Obesity and Related Metabolic Complications. Cell Metabolism. 2016, 23, 712–724. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Hu, S.; Liao, Y.; Zheng, J.; Zeng, T.; Zhong, X.; Liu, G.; Gou, L.; Chen, L. Vascular endothelial growth factor B promotes transendothelial fatty acid transport into skeletal muscle via histone modifications during catch-up growth. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E1031–E1043. [Google Scholar] [CrossRef]
- Ling, M.; Quan, L.; Lai, X.; Lang, L.; Li, F.; Yang, X.; Fu, Y.; Feng, S.; Yi, X.; Zhu, C.; et al. VEGFB Promotes Myoblasts Proliferation and Differentiation through VEGFR1-PI3K/Akt Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 3352. [Google Scholar] [CrossRef]
- Shibuya, M. Vascular endothelial growth factor receptor-1 (VEGFR-1/Flt-1): A dual regulator for angiogenesis. Angiogenesis 2006, 9, 225–230; discussion 231. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Positive and negative modulation of angiogenesis by VEGFR1 ligands. Sci. Signal. 2009, 2, re1. [Google Scholar] [CrossRef] [PubMed]
- Lal, N.; Puri, K.; Rodrigues, B. Vascular Endothelial Growth Factor B and Its Signaling. Front. Cardiovasc. Med. 2018, 5, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, T.; Hosaka, K.; Fischer, C.; Lim, S.; Andersson, P.; Abe, M.; Iwamoto, H.; Gao, Y.; Wang, X.; Fong, G.H.; et al. Ablation of endothelial VEGFR1 improves metabolic dysfunction by inducing adipose tissue browning. J. Exp. Med. 2018, 215, 611–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagberg, C.E.; Falkevall, A.; Wang, X.; Larsson, E.; Huusko, J.; Nilsson, I.; van Meeteren, L.A.; Samen, E.; Lu, L.; Vanwildemeersch, M.; et al. Vascular endothelial growth factor B controls endothelial fatty acid uptake. Nature 2010, 464, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Li, D.; Wang, X.; Jia, J.; Chen, Y.; Yao, Y.; Zhao, C.; Lu, X.; Zhang, S.; Togo, J.; et al. VEGF and VEGFB Play Balancing Roles in Adipose Differentiation, Gene Expression, and Function. Endocrinology 2018, 159, 2036–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhao, M.; Wang, C.; Wen, H.; Zhang, Y.; Lu, M.; Adlat, S.; Zheng, T.; Zhang, M.; Li, D.; et al. Adipose vascular endothelial growth factor B is a major regulator of energy metabolism. J. Endocrinol. 2020, 244, 511–521. [Google Scholar] [CrossRef]
- Cao, Y. Angiogenesis and vascular functions in modulation of obesity, adipose metabolism, and insulin sensitivity. Cell Metab. 2013, 18, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Honek, J.; Xue, Y.; Seki, T.; Cao, Z.; Andersson, P.; Yang, X.; Hosaka, K.; Cao, Y. Cold-induced activation of brown adipose tissue and adipose angiogenesis in mice. Nat. Protoc. 2012, 7, 606–615. [Google Scholar] [CrossRef]
- Hagberg, C.; Mehlem, A.; Falkevall, A.; Muhl, L.; Eriksson, U. Endothelial fatty acid transport: Role of vascular endothelial growth factor B. Physiology 2013, 28, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagberg, C.E.; Mehlem, A.; Falkevall, A.; Muhl, L.; Fam, B.C.; Ortsater, H.; Scotney, P.; Nyqvist, D.; Samen, E.; Lu, L.; et al. Targeting VEGF-B as a novel treatment for insulin resistance and type 2 diabetes. Nature 2012, 490, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Ning, F.C.; Jensen, N.; Mi, J.; Lindstrom, W.; Balan, M.; Muhl, L.; Eriksson, U.; Nilsson, I.; Nyqvist, D. VEGF-B ablation in pancreatic beta-cells upregulates insulin expression without affecting glucose homeostasis or islet lipid uptake. Sci. Rep. 2020, 10, 923. [Google Scholar] [CrossRef]
- Hsueh, T.Y.; Baum, J.I.; Huang, Y. Effect of Eicosapentaenoic Acid and Docosahexaenoic Acid on Myogenesis and Mitochondrial Biosynthesis during Murine Skeletal Muscle Cell Differentiation. Front. Nutr. 2018, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasanen, M.; Sultan, I.; Paech, J.; Hemanthakumar, K.A.; Yu, W.; He, L.; Tang, J.; Sun, Y.; Hlushchuk, R.; Huan, X.; et al. VEGF-B Promotes Endocardium-Derived Coronary Vessel Development and Cardiac Regeneration. Circulation 2021, 143, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Huusko, J.; Lottonen, L.; Merentie, M.; Gurzeler, E.; Anisimov, A.; Miyanohara, A.; Alitalo, K.; Tavi, P.; Yla-Herttuala, S. AAV9-mediated VEGF-B gene transfer improves systolic function in progressive left ventricular hypertrophy. Mol. Ther. 2012, 20, 2212–2221. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kumar, A.; Zhang, F.; Lee, C.; Tang, Z. Complicated life, complicated VEGF-B. Trends. Mol. Med. 2012, 18, 119–127. [Google Scholar] [CrossRef]
- Lal, N.; Chiu, A.P.; Wang, F.; Zhang, D.; Jia, J.; Wan, A.; Vlodavsky, I.; Hussein, B.; Rodrigues, B. Loss of VEGFB and its signaling in the diabetic heart is associated with increased cell death signaling. Am. J. Physiol. Heart Circ. Physiol 2017, 312, H1163–H1175. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Jin, K.; Childs, J.T.; Xie, L.; Mao, X.O.; Greenberg, D.A. Vascular endothelial growth factor-B (VEGFB) stimulates neurogenesis: Evidence from knockout mice and growth factor administration. Dev. Biol. 2006, 289, 329–335. [Google Scholar] [CrossRef] [Green Version]
- d’Audigier, C.; Gautier, B.; Yon, A.; Alili, J.M.; Guerin, C.L.; Evrard, S.M.; Godier, A.; Haviari, S.; Reille-Serroussi, M.; Huguenot, F.; et al. Targeting VEGFR1 on endothelial progenitors modulates their differentiation potential. Angiogenesis 2014, 17, 603–616. [Google Scholar] [CrossRef]
- Kivela, R.; Hemanthakumar, K.A.; Vaparanta, K.; Robciuc, M.; Izumiya, Y.; Kidoya, H.; Takakura, N.; Peng, X.; Sawyer, D.B.; Elenius, K.; et al. Endothelial Cells Regulate Physiological Cardiomyocyte Growth via VEGFR2-Mediated Paracrine Signaling. Circulation 2019, 139, 2570–2584. [Google Scholar] [CrossRef] [PubMed]
- Bosco, J.; Zhou, Z.; Gabriels, S.; Verma, M.; Liu, N.; Miller, B.K.; Gu, S.; Lundberg, D.M.; Huang, Y.; Brown, E.; et al. VEGFR-1/Flt-1 inhibition increases angiogenesis and improves muscle function in a mouse model of Duchenne muscular dystrophy. Mol. Ther. Methods Clin. Dev. 2021, 21, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.; Shimizu-Motohashi, Y.; Asakura, Y.; Ennen, J.P.; Bosco, J.; Zhou, Z.; Fong, G.H.; Josiah, S.; Keefe, D.; Asakura, A. Inhibition of FLT1 ameliorates muscular dystrophy phenotype by increased vasculature in a mouse model of Duchenne muscular dystrophy. PLoS Genet. 2019, 15, e1008468. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, M.; Lai, X.; Quan, L.; Li, F.; Lang, L.; Fu, Y.; Feng, S.; Yi, X.; Zhu, C.; Gao, P.; et al. Knockdown of VEGFB/VEGFR1 Signaling Promotes White Adipose Tissue Browning and Skeletal Muscle Development. Int. J. Mol. Sci. 2022, 23, 7524. https://doi.org/10.3390/ijms23147524
Ling M, Lai X, Quan L, Li F, Lang L, Fu Y, Feng S, Yi X, Zhu C, Gao P, et al. Knockdown of VEGFB/VEGFR1 Signaling Promotes White Adipose Tissue Browning and Skeletal Muscle Development. International Journal of Molecular Sciences. 2022; 23(14):7524. https://doi.org/10.3390/ijms23147524
Chicago/Turabian StyleLing, Mingfa, Xumin Lai, Lulu Quan, Fan Li, Limin Lang, Yiming Fu, Shengchun Feng, Xin Yi, Canjun Zhu, Ping Gao, and et al. 2022. "Knockdown of VEGFB/VEGFR1 Signaling Promotes White Adipose Tissue Browning and Skeletal Muscle Development" International Journal of Molecular Sciences 23, no. 14: 7524. https://doi.org/10.3390/ijms23147524
APA StyleLing, M., Lai, X., Quan, L., Li, F., Lang, L., Fu, Y., Feng, S., Yi, X., Zhu, C., Gao, P., Zhu, X., Wang, L., Shu, G., Jiang, Q., & Wang, S. (2022). Knockdown of VEGFB/VEGFR1 Signaling Promotes White Adipose Tissue Browning and Skeletal Muscle Development. International Journal of Molecular Sciences, 23(14), 7524. https://doi.org/10.3390/ijms23147524