
The Intracellular Distribution of the Small GTPase Rho5 and Its Dimeric Guanidine Nucleotide Exchange Factor Dck1/Lmo1 Determine Their Function in Oxidative Stress Response


Abstract
:1. Introduction
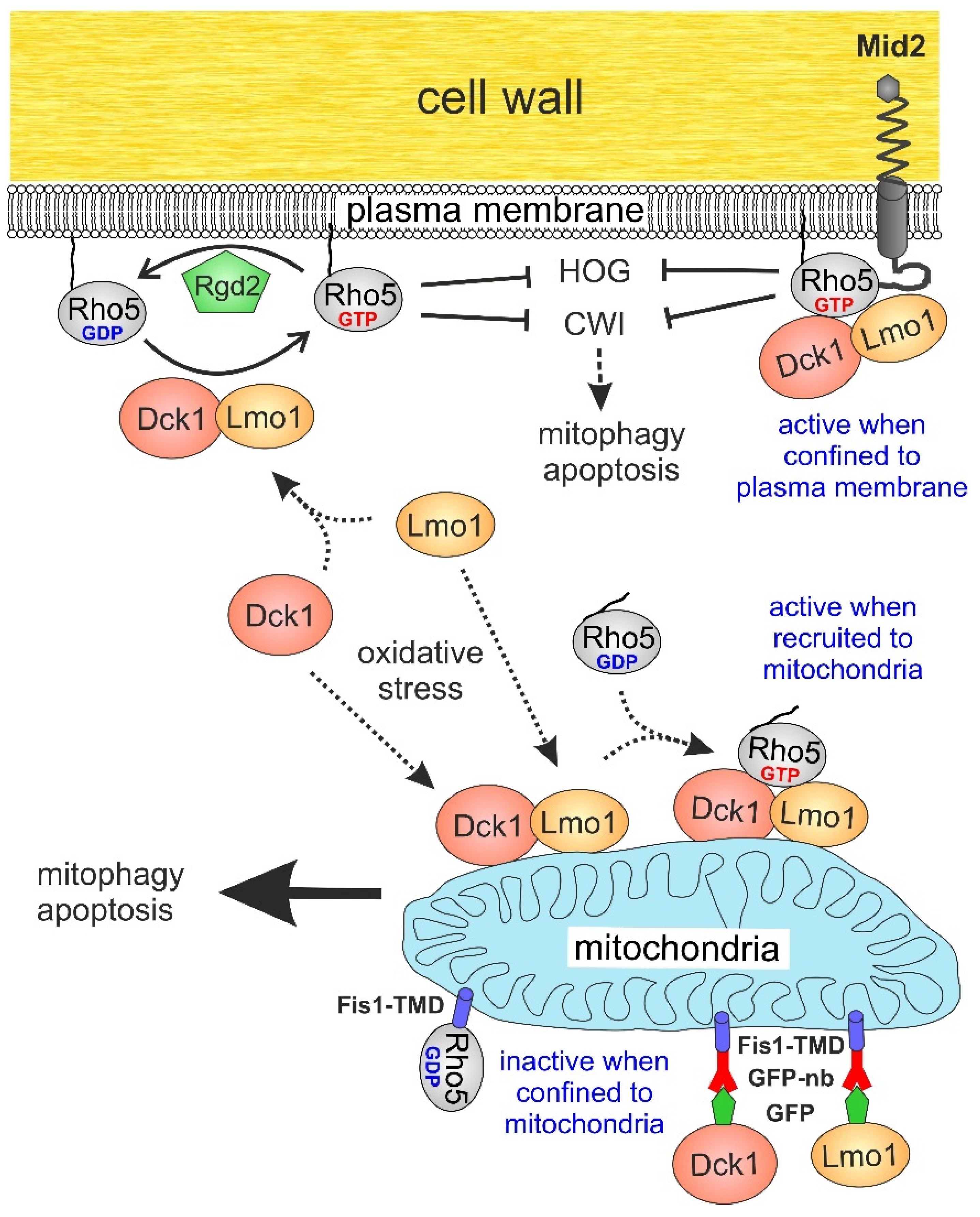
2. Results
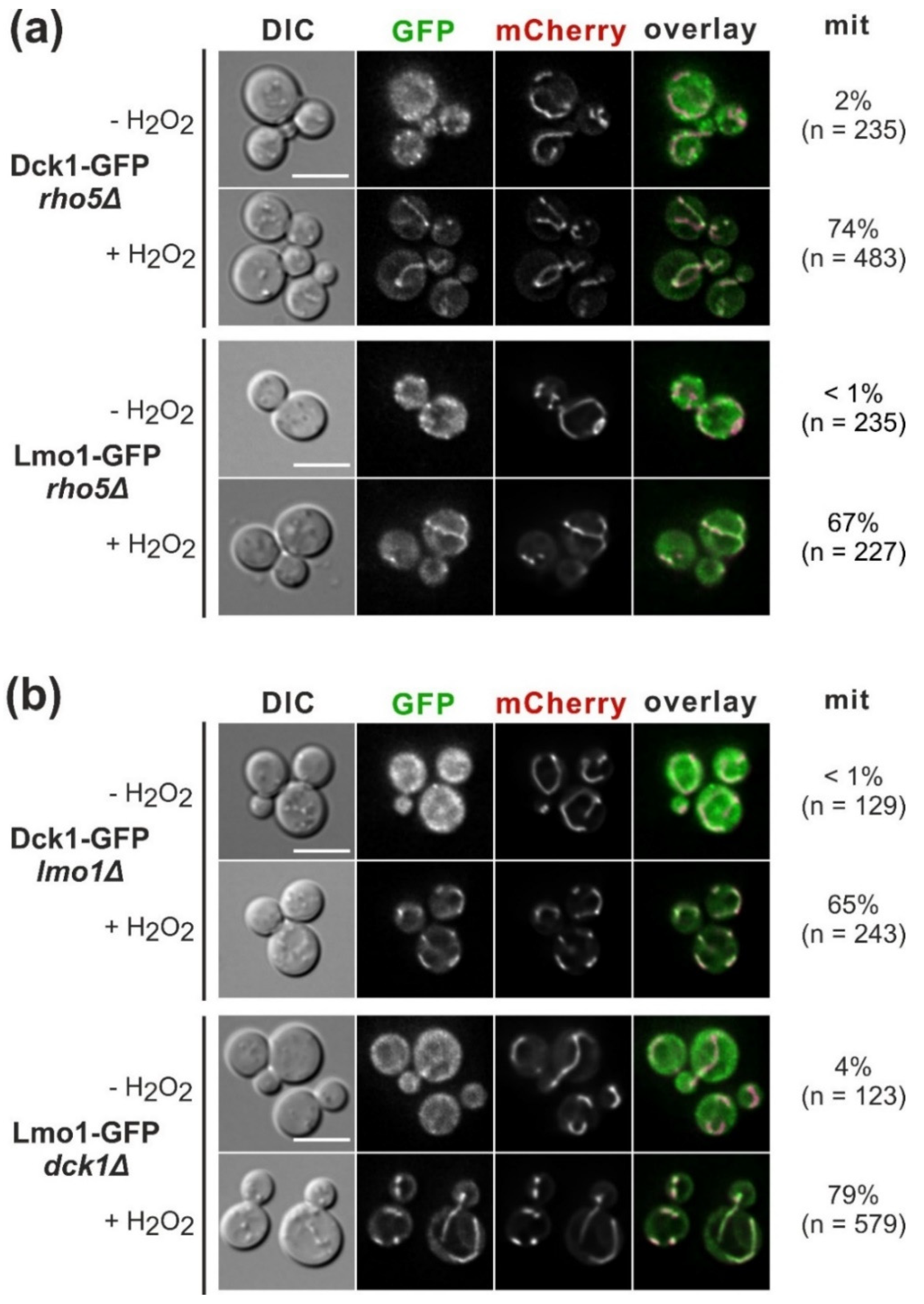
2.1. The GEF Subunits Dck1 and Lmo1 Translocate to Mitochondria Independent from Each Other or Rho5
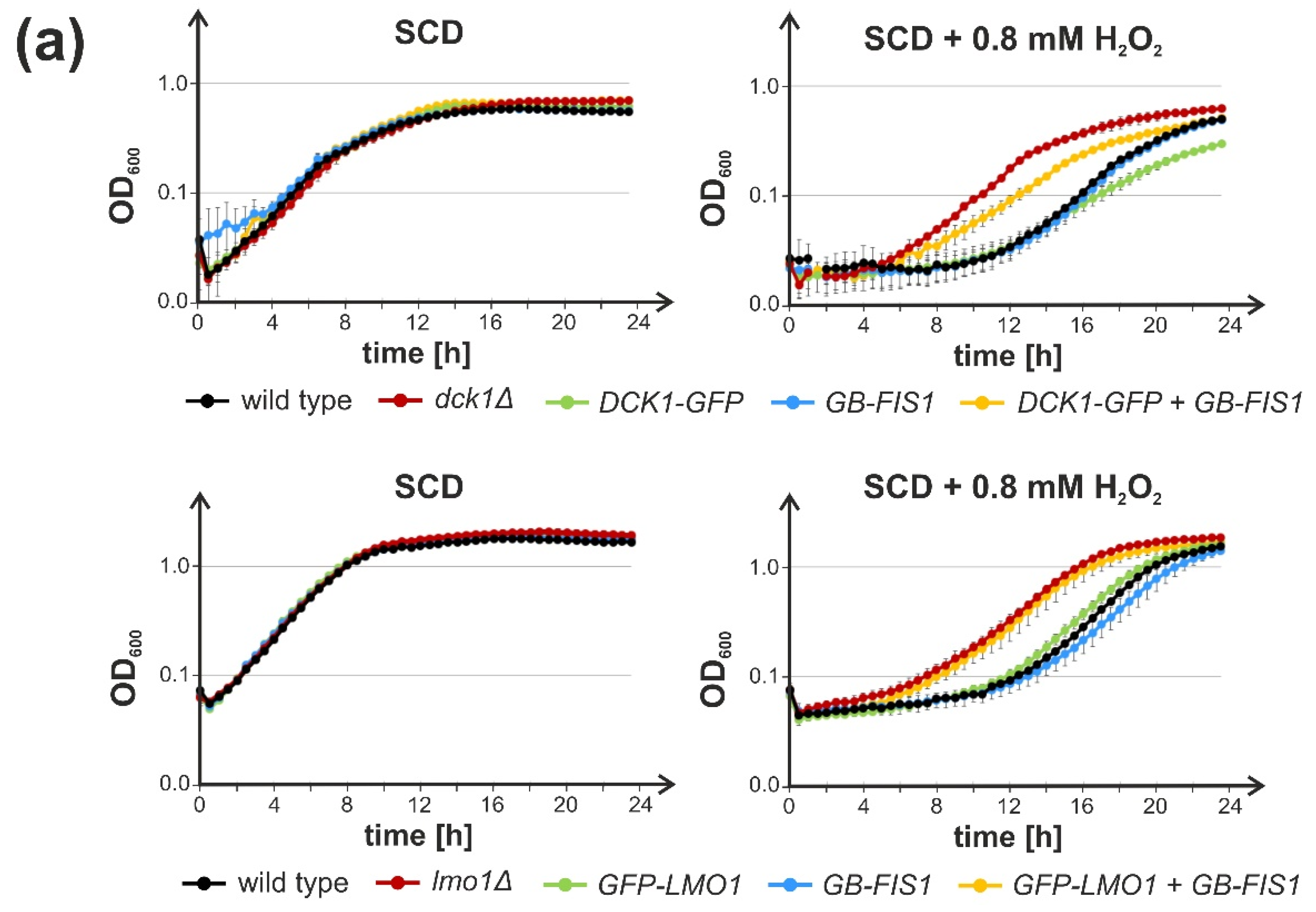
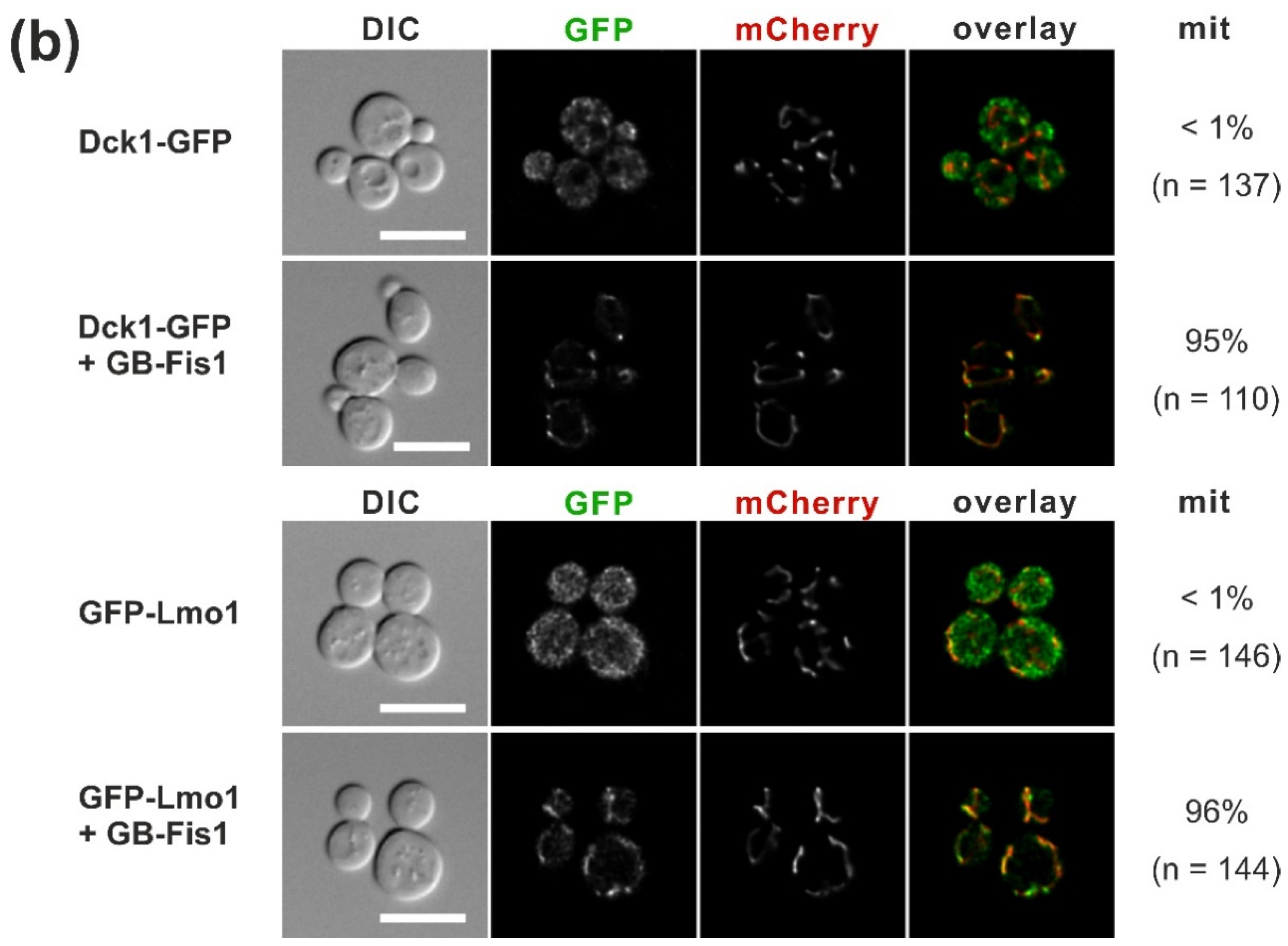
2.2. Trapping of Dck1 or Lmo1 to Mitochondria Impedes Rho5 Function in the Oxidative Stress Response
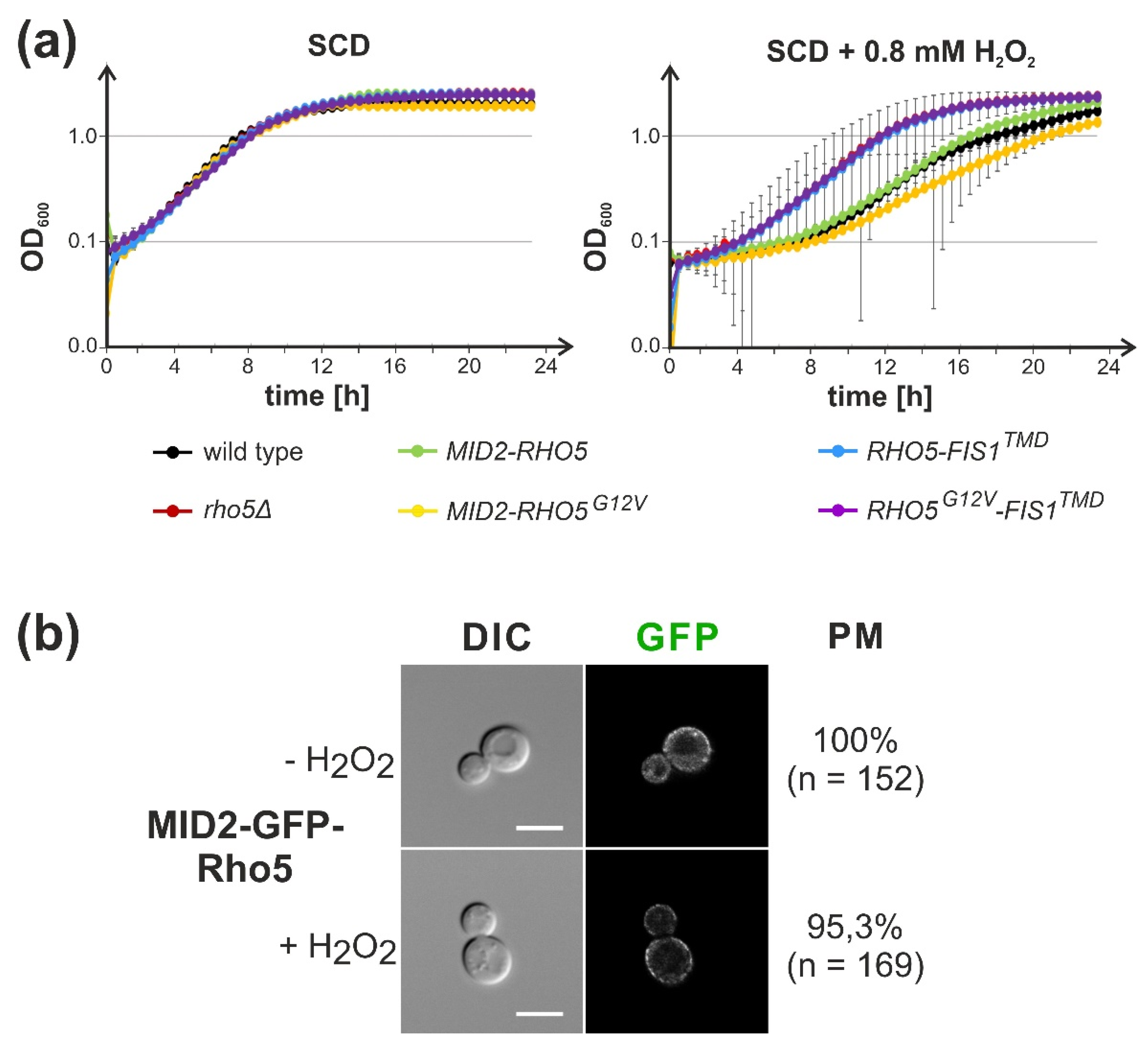
2.3. Attachment of Rho5 or its GEF Subunits to the Plasma Membrane Does Not Impair Its Repressor Function in Oxidative Stress Response
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Construction of Plasmids, Deletion Mutants and Gene Tagging
4.3. Fluorescence Microscopy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hall, A. Rho family GTPases. Biochem. Soc. Trans. 2012, 40, 1378–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, P.; Rincon, S.A. Rho GTPases: Regulation of cell polarity and growth in yeasts. Biochem. J. 2010, 426, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiya-Kawasaki, M.; Abe, M.; Saka, A.; Watanabe, D.; Kono, K.; Minemura-Asakawa, M.; Ishihara, S.; Watanabe, T.; Ohya, Y. Dissection of upstream regulatory components of the Rho1p effector, 1,3-beta-glucan synthase, in Saccharomyces cerevisiae. Genetics 2002, 162, 663–676. [Google Scholar] [CrossRef]
- Smith, J.A.; Hall, A.E.; Rose, M.D. Membrane curvature directs the localization of Cdc42p to novel foci required for cell-cell fusion. J. Cell Biol. 2017, 216, 3971–3980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, A.L.; Hall, A. Rho GTPases and their effector proteins. Biochem. J. 2000, 348 Pt 2, 241–255. [Google Scholar] [CrossRef]
- Bos, J.L.; Rehmann, H.; Wittinghofer, A. GEFs and GAPs: Critical elements in the control of small G proteins. Cell 2007, 129, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-binding proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar] [CrossRef]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell. Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef]
- Wennerberg, K.; Der, C.J. Rho-family GTPases: It’s not only Rac and Rho (and I like it). J. Cell Sci. 2004, 117, 1301–1312. [Google Scholar] [CrossRef] [Green Version]
- Roberts, P.J.; Mitin, N.; Keller, P.J.; Chenette, E.J.; Madigan, J.P.; Currin, R.O.; Cox, A.D.; Wilson, O.; Kirschmeier, P.; Der, C.J. Rho Family GTPase modification and dependence on CAAX motif-signaled posttranslational modification. J. Biol. Chem. 2008, 283, 25150–25163. [Google Scholar] [CrossRef] [Green Version]
- Tiedje, C.; Sakwa, I.; Just, U.; Hofken, T. The Rho GDI Rdi1 regulates Rho GTPases by distinct mechanisms. Mol. Biol. Cell. 2008, 19, 2885–2896. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, H.P.; Huppert, S.; Lorberg, A.; Heinisch, J.J. Rho5p downregulates the yeast cell integrity pathway. J. Cell Sci. 2002, 115, 3139–3148. [Google Scholar] [CrossRef] [PubMed]
- Hühn, J.; Musielak, M.; Schmitz, H.P.; Heinisch, J.J. Fungal homologues of human Rac1 as emerging players in signal transduction and morphogenesis. Int. Microbiol. 2020, 23, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Annan, R.B.; Wu, C.; Waller, D.D.; Whiteway, M.; Thomas, D.Y. Rho5p is involved in mediating the osmotic stress response in Saccharomyces cerevisiae, and its activity is regulated via Msi1p and Npr1p by phosphorylation and ubiquitination. Eukaryot. Cell. 2008, 7, 1441–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, H.P.; Jendretzki, A.; Sterk, C.; Heinisch, J.J. The small yeast GTPase Rho5 and its dimeric GEF Dck1/Lmo1 respond to glucose starvation. Int. J. Mol. Sci. 2018, 19, 2186. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Kang, P.J.; Park, H.O. The Rho5 GTPase is necessary for oxidant-induced cell death in budding yeast. Proc. Natl. Acad. Sci. USA 2008, 105, 1522–1527. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, H.P.; Jendretzki, A.; Wittland, J.; Wiechert, J.; Heinisch, J.J. Identification of Dck1 and Lmo1 as upstream regulators of the small GTPase Rho5 in Saccharomyces cerevisiae. Mol. Microbiol. 2015, 96, 306–324. [Google Scholar] [CrossRef] [Green Version]
- Mao, K.; Klionsky, D.J. MAPKs regulate mitophagy in Saccharomyces cerevisiae. Autophagy 2011, 7, 1564–1565. [Google Scholar] [CrossRef] [Green Version]
- Mao, K.; Wang, K.; Zhao, M.; Xu, T.; Klionsky, D.J. Two MAPK-signaling pathways are required for mitophagy in Saccharomyces cerevisiae. J. Cell Biol. 2011, 193, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Lee, M.E.; Entezari, M.; Jung, C.H.; Kim, Y.; Park, Y.; Fioretti, J.D.; Huh, W.K.; Park, H.O.; Kang, P.J. Genome-wide studies of Rho5-interacting proteins that are involved in oxidant-induced cell death in budding yeast. G3 Genes Genomes Genet. 2019, 9, 921–931. [Google Scholar] [CrossRef] [Green Version]
- Lam, B.D.; Hordijk, P.L. The Rac1 hypervariable region in targeting and signaling: A tail of many stories. Small GTPases 2013, 4, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterk, C.; Graber, L.; Schmitz, H.P.; Heinisch, J.J. Analysis of functional domains in Rho5, the yeast homolog of human Rac1 GTPase, in oxidative stress response. Int. J. Mol. Sci. 2019, 20, 5550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musielak, M.; Sterk, C.C.; Schubert, F.; Meyer, C.; Paululat, A.; Heinisch, J.J. The small GTPase KlRho5 responds to oxidative stress and affects cytokinesis. J. Cell Sci. 2021, 134, jcs258301. [Google Scholar] [CrossRef] [PubMed]
- Grimsley, C.M.; Kinchen, J.M.; Tosello-Trampont, A.C.; Brugnera, E.; Haney, L.B.; Lu, M.; Chen, Q.; Klingele, D.; Hengartner, M.O.; Ravichandran, K.S. Dock180 and ELMO1 proteins cooperate to promote evolutionarily conserved Rac-dependent cell migration. J. Biol. Chem. 2004, 279, 6087–6097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugnera, E.; Haney, L.; Grimsley, C.; Lu, M.; Walk, S.F.; Tosello-Trampont, A.C.; Macara, I.G.; Madhani, H.; Fink, G.R.; Ravichandran, K.S. Unconventional Rac-GEF activity is mediated through the Dock180-ELMO complex. Nat. Cell Biol. 2002, 4, 574–582. [Google Scholar] [CrossRef]
- Cote, J.F.; Vuori, K. Identification of an evolutionarily conserved superfamily of DOCK180-related proteins with guanine nucleotide exchange activity. J. Cell Sci. 2002, 115, 4901–4913. [Google Scholar] [CrossRef] [Green Version]
- Gadea, G.; Blangy, A. Dock-family exchange factors in cell migration and disease. Eur. J. Cell Biol. 2014. [Google Scholar] [CrossRef]
- Marei, H.; Malliri, A. Rac1 in human diseases: The therapeutic potential of targeting Rac1 signaling regulatory mechanisms. Small GTPases 2017, 8, 139–163. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.F. Rho GTPases, their post-translational modifications, disease-associated mutations and pharmacological inhibitors. Small GTPases. 2018, 9, 203–215. [Google Scholar] [CrossRef]
- Takenaka, N.; Nihata, Y.; Satoh, T. Rac1 activation caused by membrane translocation of a guanine nucleotide exchange factor in Akt2-mediated insulin signaling in mouse skeletal muscle. PLoS ONE 2016, 11, e0155292. [Google Scholar] [CrossRef] [Green Version]
- Kock, C.; Arlt, H.; Ungermann, C.; Heinisch, J.J. Yeast cell wall integrity sensors form specific plasma membrane microdomains important for signalling. Cell. Microbiol. 2016, 18, 1251–1267. [Google Scholar] [CrossRef] [Green Version]
- Straede, A.; Heinisch, J.J. Functional analyses of the extra- and intracellular domains of the yeast cell wall integrity sensors Mid2 and Wsc1. FEBS Lett. 2007, 581, 4495–4500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchers, A.C.; Langemeyer, L.; Ungermann, C. Who’s in control? Principles of Rab GTPase activation in endolysosomal membrane trafficking and beyond. J. Cell Biol. 2021, 220, e202105120. [Google Scholar] [CrossRef] [PubMed]
- Payapilly, A.; Malliri, A. Compartmentalisation of RAC1 signalling. Curr. Opin. Cell Biol. 2018, 54, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Remorino, A.; De Beco, S.; Cayrac, F.; Di Federico, F.; Cornilleau, G.; Gautreau, A.; Parrini, M.C.; Masson, J.B.; Dahan, M.; Coppey, M. Gradients of Rac1 nanoclusters support spatial patterns of Rac1 signaling. Cell Rep. 2017, 21, 1922–1935. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Strich, R.; Cooper, K.F. Slt2p phosphorylation induces cyclin C nuclear-to-cytoplasmic translocation in response to oxidative stress. Mol. Biol. Cell. 2014, 25, 1396–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, S.D.; Hanley, S.E.; Beishke, T.; Tati, P.D.; Cooper, K.F. Ubiquitin-proteasome-mediated cyclin C degradation promotes cell survival following nitrogen starvation. Mol. Biol. Cell. 2020, 31, 1015–1031. [Google Scholar] [CrossRef] [PubMed]
- Montella-Manuel, S.; Pujol-Carrion, N.; de la Torre-Ruiz, M.A. The cell wall integrity receptor Mtl1 contributes to articulate autophagic responses when glucose availability is compromised. J. Fungi 2021, 7, 903. [Google Scholar] [CrossRef]
- Kirchrath, L.; Lorberg, A.; Schmitz, H.P.; Gengenbacher, U.; Heinisch, J.J. Comparative genetic and physiological studies of the MAP kinase Mpk1p from Kluyveromyces lactis and Saccharomyces cerevisiae. J. Mol. Biol. 2000, 300, 743–758. [Google Scholar] [CrossRef]
- Schacherer, J.; Ruderfer, D.M.; Gresham, D.; Dolinski, K.; Botstein, D.; Kruglyak, L. Genome-wide analysis of nucleotide-level variation in commonly used Saccharomyces cerevisiae strains. PLoS ONE 2007, 2, e322. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.D.; Winston, F.; Hieter, P. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1990. [Google Scholar]
- Rothstein, R. Targeting, disruption, replacement, and allele rescue: Integrative DNA transformation in yeast. Methods Enzymol. 1991, 194, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Gueldener, U.; Heinisch, J.; Koehler, G.J.; Voss, D.; Hegemann, J.H. A second set of loxP marker cassettes for Cre-mediated multiple gene knockouts in budding yeast. Nucleic Acids Res. 2002, 30, e23. [Google Scholar] [CrossRef] [Green Version]
- Gietz, R.D.; Sugino, A. New yeast-Escherichia coli shuttle vectors constructed with in vitro mutagenized yeast genes lacking six-base pair restriction sites. Gene 1988, 74, 527–534. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Genotype 1 |
|---|---|
| FSO4-3A | MATalpha ura3-52 his3-11,15 leu2-3,112 |
| FSO4-3D | MATalpha ura3-52 his3-11,15 leu2-3,112 rho5::kanMX |
| FSO4-8A | MATa ura3-52 his3-11,15 leu2-3,112 |
| FSO4-8B | MATa ura3-52 his3-11,15 leu2-3,112 rho5::kanMX |
| HCLO01-3B | MATa ura3-52 his3-11,15 leu2-3,112 GFP-LMO1-KlLEU2 |
| HCLO01-8B | MATalpha ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] |
| HCLO01-15C | MATa ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] GFP-LMO1-KlLEU2 |
| HCLO01-23D | MATa ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] GFP-LMO1-KlLEU2 |
| HCLO01-27D | MATalpha ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] |
| HCSO20 | MATalpha ura3-52 his3-11,5 leu2-3,112 rho5::kanMX DCK1-3GFP-SkHIS3 |
| HCSO25 | MATa ura3-52 his3-11,5 leu2-3,112 rho5::kanMX LMO1-3GFP-SkHIS3 |
| HCSO26 | MATalpha ura3-52 his3-11,5 leu2-3,112 lmo1::kanMX DCK1-3GFP-SkHIS3 |
| HCSO33 | MATalpha ura3-52 his3-11,5 leu2-3,112 dck1::kanMX LMO1-3GFP-SkHIS3 |
| HD56-5A/ dck1ΔH1 | MATalpha ura3-52 his3-11,15 leu2-3,112 dck1::SpHIS5 |
| HD56-5A/ dck1ΔKL6 | MATalpha ura3-52 his3-11,15 leu2-3,112 dck1::KlLEU2 |
| HFSO83 | MATalpha ura3-52 his3-11,15 leu2-3,112 rho5:kanMX MID2-RHO5G12V::SkHIS3 |
| HLBO19-1B | MATalpha ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] GFP-LMO1-KlLEU2 IDP1-mCherry-kanMX |
| HLBO19-3A | MATalpha ura3-52 his3-11,5 leu2-3,112 GFP-LMO1-KlLEU2 |
| HLBO19-3B | MATa ura3-52 his3-11,5 leu2-3,112 GFP-LMO1-KlLEU2 IDP1-mCherry-kanMX |
| HLBO19-5A | MATalpha ura3-52 his3-11,5 leu2-3,112 GFP-LMO1-KlLEU2 IDP1-mCherry-kanMX |
| HLBO19-8C | MATa ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] GFP-LMO1-KlLEU2 IDP1-mCherry-kanMX |
| HLBO20-4B | MATa ura3-52 his3-11,15 leu2-3,112 |
| HLBO21-2A | MATa ura3-52 his3-11,15 leu2-3,112 DCK1-3GFP-SpHIS5 IDP1-mCherry-kanMX |
| HLBO21-3D | MATa ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] DCK1-3GFP-SpHIS5IDP1-mCherry-kanMX |
| HLBO21-6A | MATalpha ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] DCK1-3GFP-SpHIS5IDP1-mCherry-kanMX |
| HLBO21-10A | MATalpha ura3-52 his3-11,15 leu2-3,112 DCK1-3GFP-SpHIS5 IDP1-mCherry-kanMX |
| HLBO22-2A | MATa ura3-52 his3-11,15 leu2-3,112 DCK1-3GFP-SpHIS5 |
| HLBO22-2D | MATalpha ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] |
| HLBO22-3B | MATalpha ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] DCK1-3GFP-SpHIS5 |
| HLBO22-4D | MATalpha ura3-52 his3-11,15 leu2-3,112 DCK1-3GFP-SpHIS5 |
| HLBO22-5B | MATa ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] DCK1-3GFP-SpHIS5 |
| HLBO22-9D | MATa ura3-52 his3-11,15 leu2-3,112::pLA012 [GB-FIS1TMD] |
| HOD464-7B | MATa ura3-52 his3-11,15 leu2-3,112 lmo1::kanMX |
| HOD500-10A | MATa ura3-52 leu2-3,113 his3-11,15 rho5::kanMX MID2-RHO5-SkHIS3 |
| HOD500-11B | MATalpha ura3-52 leu2-3,113 his3-11,15 rho5::kanMX MID2-RHO5-SkHIS3 |
| HOD512-3A | MATa ura3-52 his3-11,15 leu2-3,112 MID2-GFP-RHO5-SkHIS3 |
| HOD529-2B | MATa ura3-52 his3-11,15 leu2-3,112::pJJH3096 [LEU2-RHO5G12V-FIS1TMD] rho5::kanMX |
| HOD529-5C | MATalpha ura3-52 his3-11,15 leu2-3,112::pJJH3096 [LEU2-RHO5G12V-FIS1TMD] rho5::kanMX |
| LBO81 | MATa ura3-52 his3-11,15 leu2-3,112 lmo1::KlURA3 |
| LBO130 | MATalpha ura3-52 his3-11,15 leu2-3,112::pJJH3024 [RHO5-FIS1TMD] rho5::kanMX |
| LBO132 | MATa ura3-52 his3-11,15 leu2-3,112::pJJH3024 [RHO5-FIS1TMD] rho5::kanMX |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bischof, L.; Schweitzer, F.; Sterk, C.C.; Heinisch, J.J. The Intracellular Distribution of the Small GTPase Rho5 and Its Dimeric Guanidine Nucleotide Exchange Factor Dck1/Lmo1 Determine Their Function in Oxidative Stress Response. Int. J. Mol. Sci. 2022, 23, 7896. https://doi.org/10.3390/ijms23147896
Bischof L, Schweitzer F, Sterk CC, Heinisch JJ. The Intracellular Distribution of the Small GTPase Rho5 and Its Dimeric Guanidine Nucleotide Exchange Factor Dck1/Lmo1 Determine Their Function in Oxidative Stress Response. International Journal of Molecular Sciences. 2022; 23(14):7896. https://doi.org/10.3390/ijms23147896
Chicago/Turabian StyleBischof, Linnet, Franziska Schweitzer, Carolin C. Sterk, and Jürgen J. Heinisch. 2022. "The Intracellular Distribution of the Small GTPase Rho5 and Its Dimeric Guanidine Nucleotide Exchange Factor Dck1/Lmo1 Determine Their Function in Oxidative Stress Response" International Journal of Molecular Sciences 23, no. 14: 7896. https://doi.org/10.3390/ijms23147896