
The Single-Stranded DNA-Binding Gene Whirly (Why1) with a Strong Pathogen-Induced Promoter from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Isolation and Characterization of Why1 Genes and Their Promoters
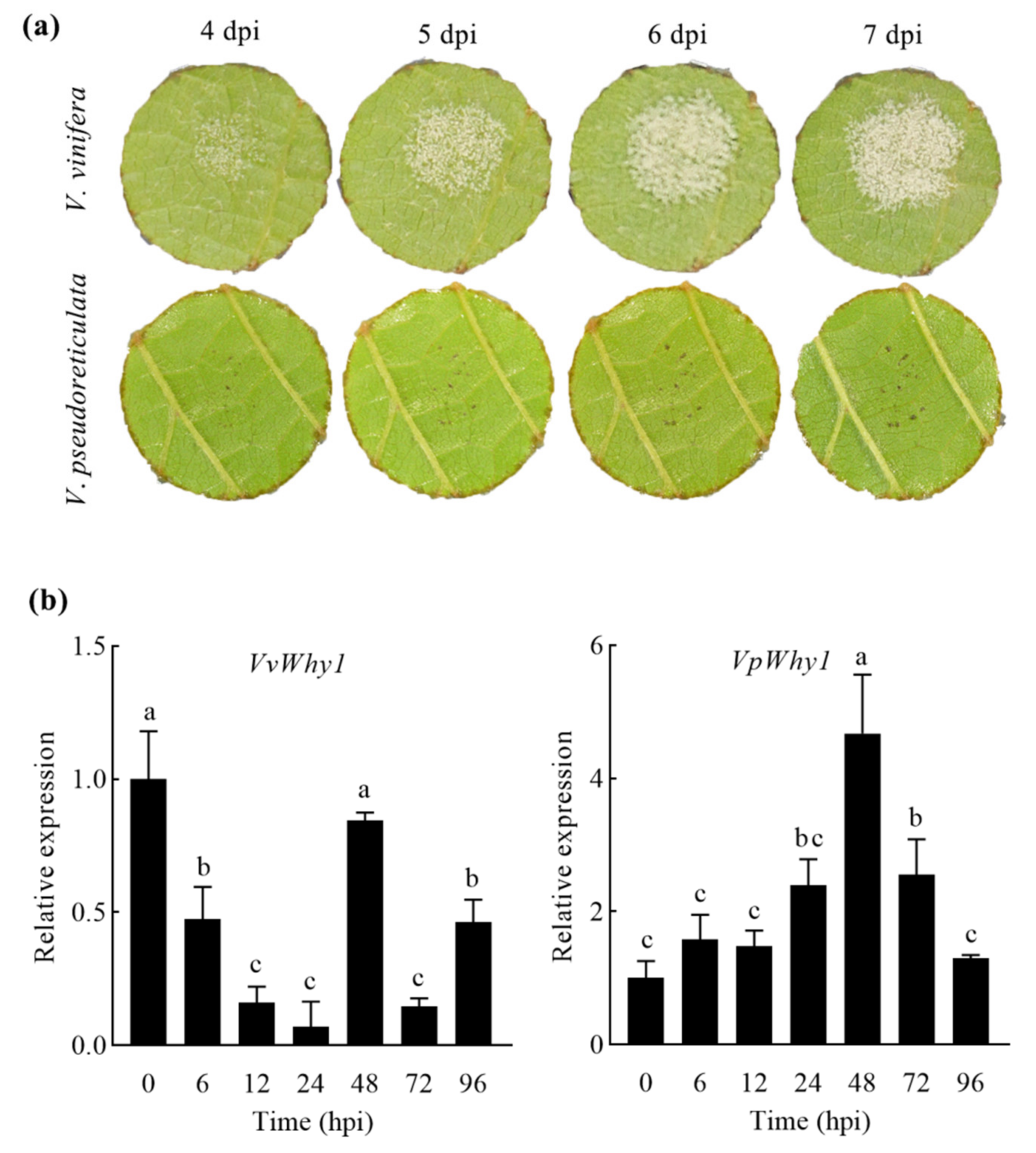
2.2. Why1 Expression under P. viticola Induction in V. vinifera and V. pseudoreticulata
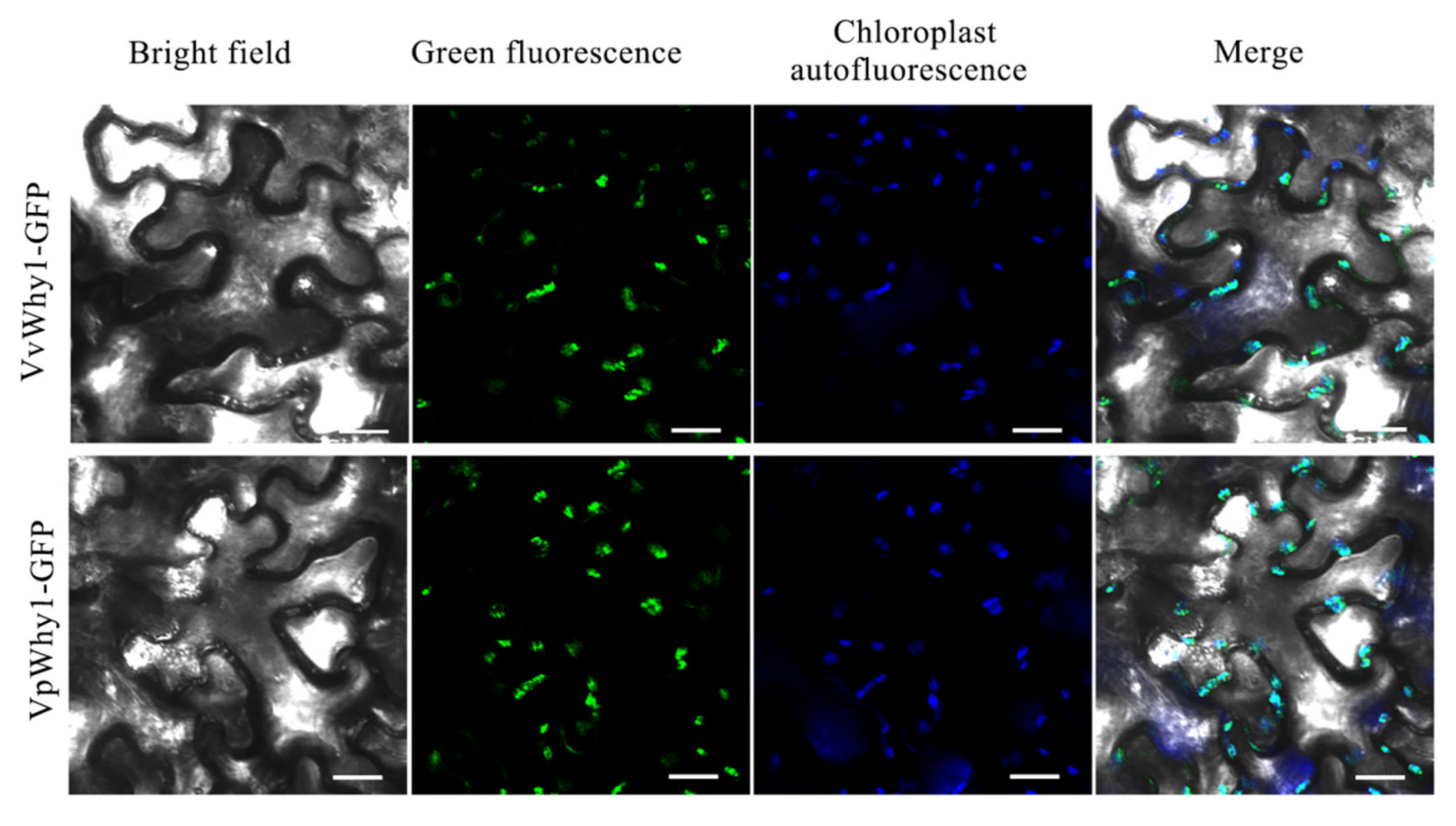
2.3. Subcellular Localization Analysis of Why1
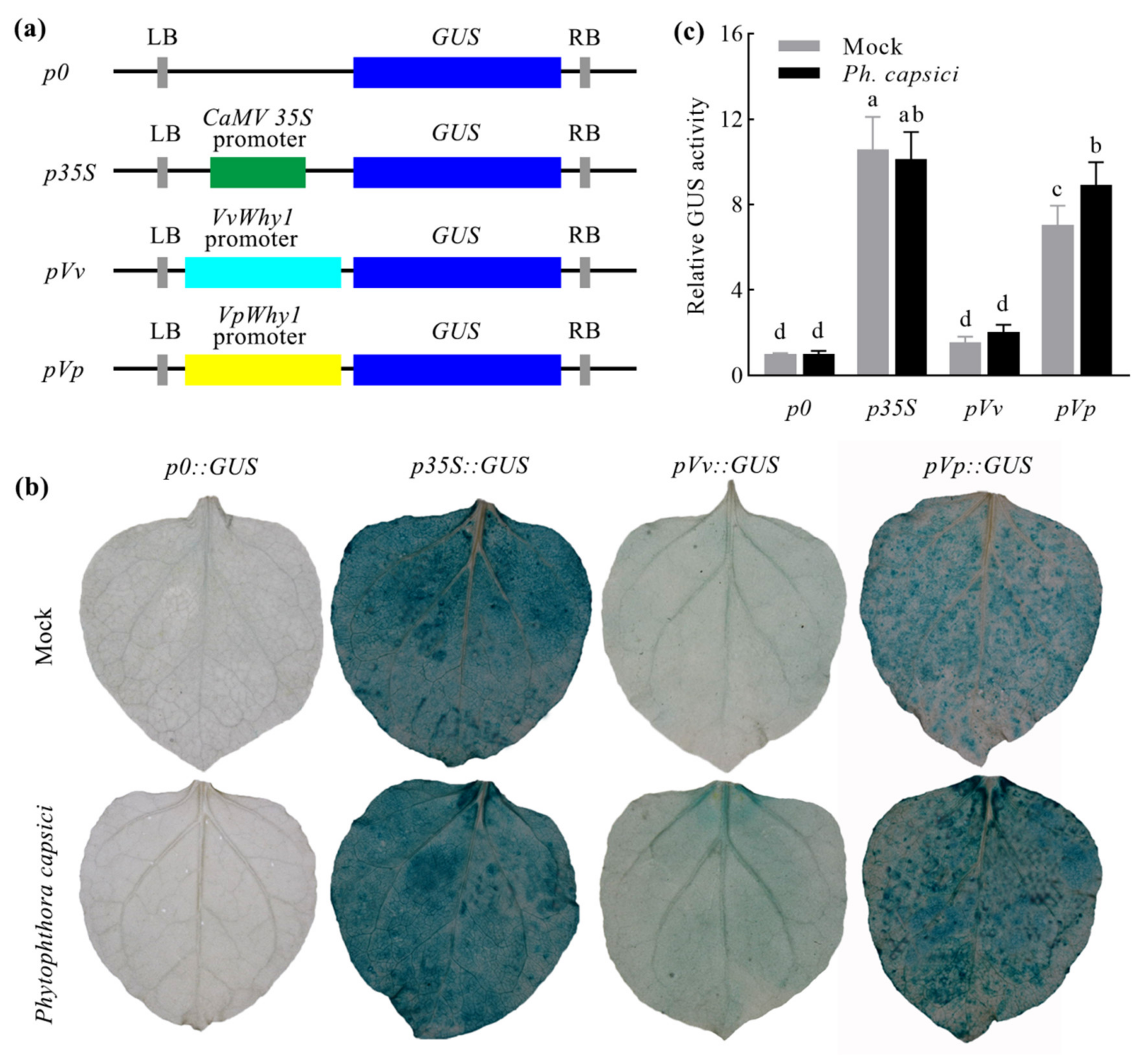
2.4. Why1 Promoter Activity Analysis
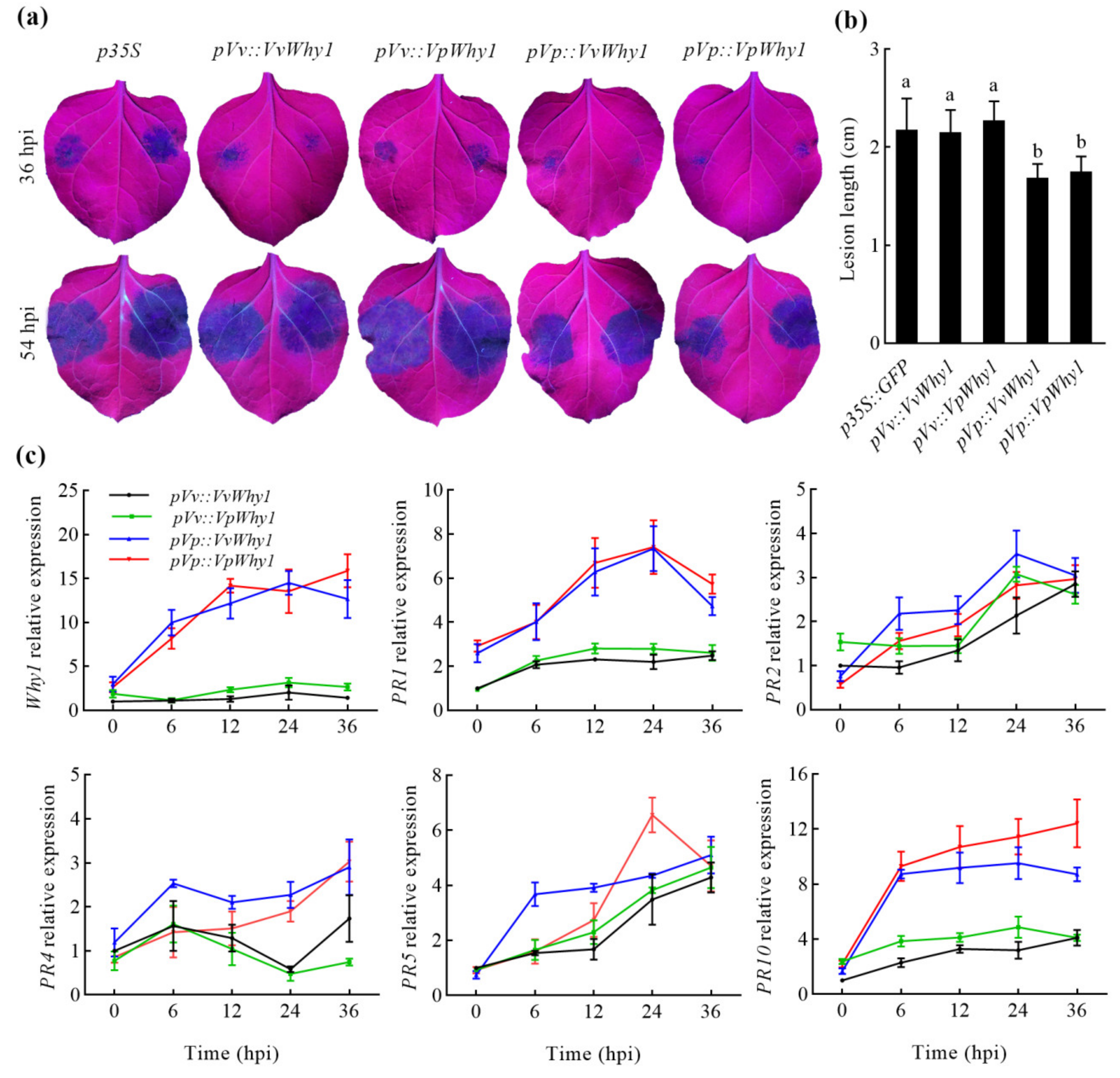
2.5. P. capsici Resistance of Why1 Genes Driven by Native and Exotic Promoters
2.6. Why1 Heterologous Expression and Pathogenesis-Related Genes in Response to P. capsici
3. Discussion
3.1. Dual Localization of Why1
3.2. Promoter Differentiation between VvWhy1 and VpWhy1
3.3. Why1 Driven by VpWhy1 Promoter (pVp) Enhanced Disease Resistance via Regulating the Expression of PR Genes
4. Materials and Methods
4.1. Plant Materials, Pathogen and Treatments
4.2. Gene Cloning and Sequence Analysis
4.3. Quantitative Real-Time PCR Analysis of Why1 under P. viticola Induction
4.4. Subcellular Localization Analysis
4.5. Promoter Activity Analysis
4.6. P. capsici Resistance Assay Transiently Expressing Why1 Driven by Native and Exotic Promoter
4.7. Why1 Heterologous Expression and Pathogenesis-Related Genes in Response to P. capsici
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| P. Viticola | Plasmopara Viticola |
| P. capsica | Phytophthora capsici |
| pVv | VvWhy1 promoter |
| pVp | VpWhy1 promoter |
| PR gene | Pathogenesis-related gene |
| R gene | Resistance gene |
| qRT-PCR | Quantitative real-time PCR |
| GFP | Green fluorescent protein |
| CDS | Coding sequence |
| dpi | Days postinoculation |
| hpi | Hours postinoculation |
References
- Jiang, H.E.; Zhang, Y.B.; Li, X.; Yao, Y.F.; Fergusond, D.K.; Lü, E.G.; Li, C.S. Evidence for early viticulture in China: Proof of a grapevine (Vitis vinifera L., Vitaceae) in the Yanghai Tombs, Xinjiang. J. Archaeol. Sci. 2009, 36, 1458–1465. [Google Scholar] [CrossRef]
- Figueiredo, J.; Cunha, J.; Eiras-Dias, J.; Silva, M.S.; Figueiredo, A. Pathogen-related specificity of subtilase VVISBT4.19 X1 in the Vitis vinifera defence response. Cienc. Tec. Vitivinic. 2020, 35, 42–48. [Google Scholar] [CrossRef]
- Cavaco, A.R.; Laureano, G.; Cunha, J.; Eiras-Dias, J.; Figueiredo, A. Fatty acid modulation and desaturase gene expression are differentially triggered in grapevine incompatible interaction with biotrophs and necrotrophs. Plant Physiol. Biochem. 2021, 163, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Marcianò, D.; Ricciardi, V.; Fassolo, E.M.; Passera, A.; Toffolatti, S.L. RNAi of a putative grapevine susceptibility gene as a possible downy mildew control strategy. Front. Plant Sci. 2021, 12, 667319. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Wang, X.; Liu, H.; Wang, Y.; Nocker, S.V.; Tu, M.; Fang, J.; Guo, J.; Li, Z.; Wang, X. Overexpression of VqWRKY31 enhances powdery mildew resistance in grapevine by promoting salicylic acid signaling and specific metabolite synthesis. Hortic. Res. 2022, 9, uhab064. [Google Scholar] [CrossRef]
- Yan, X.; Qiao, H.; Zhang, X.; Guo, C.; Wang, M.; Wang, Y.; Wang, X. Analysis of the grape (Vitis vinifera L.) thaumatin-like protein (TLP) gene family and demonstration that TLP29 contributes to disease resistance. Sci. Rep. 2017, 7, 4269. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, M.C.; Perazzolli, M.; Matafora, V.; Moretto, M.; Bachi, A.; Pertot, I. Proteomic analysis of grapevine resistance induced by Trichoderma harzianum T39 reveals specific defence pathways activated against downy mildew. J. Exp. Bot. 2012, 63, 6237–6251. [Google Scholar] [CrossRef]
- Li, M.Y.; Jiao, Y.T.; Wang, Y.T.; Zhang, N.; Wang, B.B.; Liu, R.Q.; Yin, X.; Xu, Y.; Liu, G.T. CRISPR/Cas9-mediated VvPR4b editing decreases downy mildew resistance in grapevine (Vitis vinifera L.). Hortic. Res. 2020, 7, 149. [Google Scholar] [CrossRef]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar] [CrossRef]
- Ma, H.; Xiang, G.Q.; Li, Z.Q.; Wang, Y.T.; Dou, M.R.; Su, L.; Yin, X.; Liu, R.Q.; Wang, Y.J.; Xu, Y. Grapevine VpPR10.1 functions in resistance to Plasmopara viticola through triggering a cell death-like defence response by interacting with VpVDAC3. Plant Biotechnol. J. 2018, 16, 1488–1501. [Google Scholar] [CrossRef] [Green Version]
- Sargolzaei, M.; Maddalena, G.; Bitsadze, N.; Maghradze, D.; Bianco, P.A.; Failla, O.; Toffolatti, S.L.; Lorenzis, G.D. Rpv29, Rpv30 and Rpv31: Three novel genomic loci associated with resistance to Plasmopara viticola in Vitis vinifera. Front. Plant Sci. 2020, 11, 562432. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Wu, W.; Lai, G.; Li, R.; Peng, Y.; Yang, B.; Wang, B.; Yin, L.; Qu, J.; Song, S.; et al. Identifying Plasmopara viticola resistance Loci in grapevine (Vitis amurensis) via genotyping-by-sequencing-based QTL mapping. Plant Physiol. Biochem. 2020, 154, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Ciubotaru, R.M.; Franceschi, P.; Zulini, L.; Stefanini, M.; Škrab, D.; Rossarolla, M.D.; Robatscher, P.; Oberhuber, M.; Vrhovsek, U.; Chitarrini, G. Mono-locus and pyramided resistant grapevine cultivars reveal early putative biomarkers upon artificial inoculation with Plasmopara viticola. Front. Plant Sci. 2021, 12, 693887. [Google Scholar] [CrossRef] [PubMed]
- Rossarolla, M.D.; Tomazetti, T.C.; Welter, L.J.; Santos, H.P.; Stefanini, M.; Trapp, O.; Guerra, M.P.; Nodari, R.O. The interplay between hormone signaling and defense gene expression in grapevine genotypes carrying genetic resistance against Plasmopara viticola. Vitis 2021, 60, 195–206. [Google Scholar] [CrossRef]
- Liu, R.; Weng, K.; Dou, M.; Chen, T.; Xu, Y. Transcriptomic analysis of Chinese wild Vitis pseudoreticulata in response to Plasmopara viticola. Protoplasma 2019, 256, 1409–1424. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Ma, F.; Li, R.; Zhou, Q.; Yao, W.; Jiao, Y.; Zhang, C.; Zhang, J.; Wang, X.; Xu, Y. VpSTS29/STS2 enhances fungal tolerance in grapevine through a positive feedback loop. Plant Cell Environ. 2019, 42, 2979–2998. [Google Scholar] [CrossRef]
- Lai, G.; Fu, P.; Liu, Y.; Jiang, X.; Lu, J. Molecular characterization and overexpression of VpRPW8s from Vitis pseudoreticulata enhances resistance to Phytophthora capsici in Nicotiana benthamiana. Int. J. Mol. Sci. 2018, 19, 839. [Google Scholar] [CrossRef] [Green Version]
- Lai, G.; Song, S.; Liu, Y.; Fu, P.; Xiang, J.; Lu, J. RPW8 promoter is involved in pathogen- And stress-inducible expression from Vitis pseudoreticulata. J. Phytopathol. 2019, 167, 65–74. [Google Scholar] [CrossRef]
- Desveaux, D.; Després, C.; Joyeux, A.; Brisson, S.N. PBF-2 is a novel single-stranded DNA binding factor implicated in PR-10a gene activation in potato. Plant Cell 2000, 12, 1477–1489. [Google Scholar] [CrossRef] [Green Version]
- Prikryl, J.; Watkins, W.K.; Friso, G.; van Wijk, K.J.; Barkan, A. A member of the Whirly family is a multifunctional RNA- and DNA-binding protein that is essential for chloroplast biogenesis. Nucleic Acids Res. 2008, 36, 5152–5165. [Google Scholar] [CrossRef]
- Krause, K.; Kilbienski, I.; Mulisch, M.; Rdiger, A.; Schfer, A.; Krupinska, K. DNA-binding proteins of the Whirly family in Arabidopsis thaliana are targeted to the organelles. FEBS Lett. 2005, 579, 3707–3712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowski, E.; Miao, Y.; Mulisch, M.; Krupinska, K. Single-stranded DNA-binding protein Whirly1 in barley leaves is located in plastids and the nucleus of the same cell. Plant Physiol. 2008, 147, 1800–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarty, D.R.; Mark Settles, A.; Suzuki, M.; Tan, B.C.; Latshaw, S.; Porch, T.; Robin, K.; Baier, J.; Avigne, W.; Lai, J. Steady-state transposon mutagenesis in inbred maize. Plant J. 2005, 44, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, K.; Kong, F.; Zhang, S.; Meng, C.; Yang, M. Whirly1 enhances tolerance to chilling stress in tomato via protection of photosystem II and regulation of starch degradation. New Phytol. 2018, 221, 1998–2012. [Google Scholar] [CrossRef]
- Zhuang, K.; Gao, Y.; Liu, Z.; Diao, P.; Sui, N.; Meng, Q.; Meng, C.; Kong, F. WHIRLY1 regulates HSP21.5A expression to promote thermotolerance in tomato. Plant Cell Physiol. 2020, 61, 169–177. [Google Scholar] [CrossRef]
- Zhuang, K.; Wang, J.; Jiao, B.; Chen, C.; Zhang, J.; Ma, N.; Meng, Q. WHIRLY1 maintains leaf photosynthetic capacity in tomato by regulating the expression of RbcS1 under chilling stress. J. Exp.Bot. 2020, 71, 3653–3663. [Google Scholar] [CrossRef]
- Cappadocia, L.; Marechal, A.; Parent, J.S.; Lepage, E.; Sygusch, J.; Brisson, N. Crystal structures of DNA-Whirly complexes and their role in Arabidopsis organelle genome repair. Plant Cell 2010, 22, 1849–1867. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.H.; Kwon, C.; Lee, M.M.; Chung, I.K. Single-stranded DNA binding factor AtWHY1 modulates telomere length homeostasis in Arabidopsis. Plant J. 2007, 49, 442–451. [Google Scholar] [CrossRef]
- Miao, Y.; Jiang, J.; Ren, Y.; Zhao, Z. The single-stranded DNA-binding protein WHIRLY1 represses WRKY53 expression and delays leaf senescence in a developmental stage-dependent manner in Arabidopsis. Plant Physiol. 2013, 163, 746–756. [Google Scholar] [CrossRef] [Green Version]
- Desveaux, D.; Subramaniam, R.; Després, C.; Mess, J.N.; Lévesque, C.; Fobert, P.R.; Dangl, J.L.; Brisson, N. A “Whirly” transcription factor is required for salicylic acid-dependent disease resistance in Arabidopsis. Dev. Cell 2004, 6, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.; Van Kammen, A. Polyacrylamide disc electrophoresis of the soluble leaf proteins from Nicotiana tabacum var. ‘Samsun’ and ‘Samsun NN’: II. Changes in protein constitution after infection with tobacco mosaic virus. Virology 1970, 40, 199–211. [Google Scholar] [CrossRef]
- Wang, X.; Bi, W.; Jing, G.; Yu, X.; Wang, H.; Liu, D. Systemic acquired resistance, NPR1, and pathogenesis-related genes in wheat and barley. J. Integr. Agric. 2018, 17, 2468–2477. [Google Scholar] [CrossRef] [Green Version]
- Desveaux, D.; Maréchal, A.; Brisson, N. Whirly transcription factors: Defense gene regulation and beyond. Trends Plant Sci. 2005, 10, 95–102. [Google Scholar] [CrossRef]
- Isemer, R.; Krause, K.; Grabe, N.; Kitahata, N.; Asami, T.; Krupinska, K. Plastid located WHIRLY1 enhances the responsiveness of Arabidopsis seedlings toward abscisic acid. Front. Plant Sci. 2012, 3, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Feng, P.; Xu, X.; Guo, H.; Ma, J.; Chi, W.; Lin, R.; Lu, C.; Zhang, L. A chloroplast envelope-bound PHD transcription factor mediates chloroplast signals to the nucleus. Nat. Commun. 2011, 2, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesaresi, P.; Kim, C. Current understanding of GUN1: A key mediator involved in biogenic retrograde signaling. Plant Cell Rep. 2019, 38, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Z.; Meyer, E.H.; Richter, A.S.; Schuster, M.; Ling, Q.; Schttler, M.A.; Walther, D.; Zoschke, R.; Grimm, B.; Jarvis, R.P.; et al. Control of retrograde signalling by protein import and cytosolic folding stress. Nat. Plants 2019, 5, 525–538. [Google Scholar] [CrossRef]
- Lin, W.; Huang, D.; Shi, X.; Deng, B.; Miao, Y. H2O2 as a feedback signal on dual-located WHIRLY1 associates with leaf senescence in Arabidopsis. Cells 2019, 8, 1585. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Zhang, H.; Huang, D.; Schenke, D.; Cai, D.; Wu, B.; Miao, Y. Dual-localized WHIRLY1 affects salicylic acid biosynthesis via coordination of ISOCHORISMATE SYNTHASE1, PHENYLALANINE AMMONIA LYASE1, and S-ADENOSYL-L-METHIONINE-DEPENDENT METHYLTRANSFERASE1. Plant Physiol. 2020, 184, 1884–1899. [Google Scholar] [CrossRef]
- Foyer, C.H.; Karpinska, B.; Krupinska, K. The functions of WHIRLY1 and REDOX-RESPONSIVE TRANSCRIPTION FACTOR 1 in cross tolerance responses in plants: A hypothesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130226. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Li, Y.; Jiang, Y.; Wu, B.; Ying, M. Phosphorylation of WHIRLY1 by CIPK14 shifts its localization and dual functions in Arabidopsis. Mol. Plant 2017, 10, 749–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nia, M.S.; Repnik, U.; Krupinska, K.; Bilger, W. The plastid-nucleus localized DNA-binding protein WHIRLY1 is required for acclimation of barley leaves to high light. Planta 2022, 255, 84. [Google Scholar] [CrossRef]
- Krause, K.; Krupinska, K. Nuclear regulators with a second home in organelles. Trends Plant Sci. 2009, 14, 194–199. [Google Scholar] [CrossRef]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–470. [Google Scholar] [CrossRef]
- Cui, K.C.; Liu, M.; Ke, G.H.; Zhang, X.Y.; Wen, Y.Q. Transient Silencing of VvCSN5 enhances powdery mildew resistance in grapevine (Vitis vinifera). Plant Cell Tissue Organ Cult. (PCTOC) 2021, 146, 621–633. [Google Scholar] [CrossRef]
- Jiao, Y.; Xu, W.; Duan, D.; Wang, Y.; Nick, P. A stilbene synthase allele from a Chinese wild grapevine confers resistance to powdery mildew by recruiting salicylic acidsignalling for efficient defence. J. Exp. Bot. 2016, 67, 5841–5856. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cuenca, J.; Wang, N.; Liang, Z.; Sun, H.; Gutierrez, B.; Xi, X.; Arro, J.; Wang, Y.; Fan, P.; et al. A key ‘foxy’ aroma gene is regulated by homology-induced promoter indels in the iconic juice grape ‘Concord’. Hortic. Res. 2020, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Desveaux, D.; Allard, J.; Brisson, N.; Sygusch, J. A new family of plant transcription factors displays a novel ssDNA-binding surface. Nat. Struct. Mol. Biol. 2002, 9, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Sinha, R.K.; Verma, S.S.; Rastogi, A. Role of pathogen-related protein 10 (PR 10) under abiotic and biotic stresses in plants. Phyton-Int. J. Exp. Bot. 2020, 89, 167–182. [Google Scholar] [CrossRef]
- Su, H.; Jiao, Y.; Wang, F.; Liu, Y.; Niu, W.; Liu, G.; Xu, Y. Overexpression of VpPR10.1 by an efficient transformation method enhances downy mildew resistance in V. vinifera. Plant Cell Rep. 2018, 37, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qiu, L.; Liu, X.; Zhang, Q.; Zhuansun, X.; Fahima, T.; Krugman, T.; Sun, Q.; Xie, C. Glycerol-induced powdery mildew resistance in wheat by regulating plant fatty acid metabolism, plant hormones cross-talk, and pathogenesis-related genes. Int. J. Mol. Sci. 2020, 21, 673. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouz, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Xiang, J.; Li, X.; Wu, J.; Yin, L.; Zhang, Y.; Lu, J. Studying the mechanism of Plasmopara viticola RxLR effectors on suppressing plant immunity. Front. Microbiol. 2016, 7, 709. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Yin, L.; Lu, J. Overexpression of pathogen-induced grapevine TIR-NB-LRR gene VaRGA1 enhances disease resistance and drought and salt tolerance in Nicotiana benthamiana. Protoplasma 2017, 254, 957–969. [Google Scholar] [CrossRef]
- Jefferson, A.R. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep. 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.; Que, Q.; Pan, R.; Wang, Q.; Gao, H.; Guan, X.; Che, J.; Lai, G. The Single-Stranded DNA-Binding Gene Whirly (Why1) with a Strong Pathogen-Induced Promoter from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici. Int. J. Mol. Sci. 2022, 23, 8052. https://doi.org/10.3390/ijms23148052
Lai C, Que Q, Pan R, Wang Q, Gao H, Guan X, Che J, Lai G. The Single-Stranded DNA-Binding Gene Whirly (Why1) with a Strong Pathogen-Induced Promoter from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici. International Journal of Molecular Sciences. 2022; 23(14):8052. https://doi.org/10.3390/ijms23148052
Chicago/Turabian StyleLai, Chengchun, Qiuxia Que, Ruo Pan, Qi Wang, Huiying Gao, Xuefang Guan, Jianmei Che, and Gongti Lai. 2022. "The Single-Stranded DNA-Binding Gene Whirly (Why1) with a Strong Pathogen-Induced Promoter from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici" International Journal of Molecular Sciences 23, no. 14: 8052. https://doi.org/10.3390/ijms23148052
APA StyleLai, C., Que, Q., Pan, R., Wang, Q., Gao, H., Guan, X., Che, J., & Lai, G. (2022). The Single-Stranded DNA-Binding Gene Whirly (Why1) with a Strong Pathogen-Induced Promoter from Vitis pseudoreticulata Enhances Resistance to Phytophthora capsici. International Journal of Molecular Sciences, 23(14), 8052. https://doi.org/10.3390/ijms23148052