Abstract
Ethylene response factors (ERFs) are involved in biotic and abiotic stress; however, the drought resistance mechanisms of many ERFs in soybeans have not been resolved. Previously, we proved that GmERF113 enhances resistance to the pathogen Phytophthora sojae in soybean. Here, we determined that GmERF113 is induced by 20% PEG-6000. Compared to the wild-type plants, soybean plants overexpressing GmERF113 (GmERF113-OE) displayed increased drought tolerance which was characterized by milder leaf wilting, less water loss from detached leaves, smaller stomatal aperture, lower Malondialdehyde (MDA) content, increased proline accumulation, and higher Superoxide dismutase (SOD) and Peroxidase (POD) activities under drought stress, whereas plants with GmERF113 silenced through RNA interference were the opposite. Chromatin immunoprecipitation and dual effector-reporter assays showed that GmERF113 binds to the GCC-box in the GmPR10-1 promoter, activating GmPR10-1 expression directly. Overexpressing GmPR10-1 improved drought resistance in the composite soybean plants with transgenic hairy roots. RNA-seq analysis revealed that GmERF113 downregulates abscisic acid 8′-hydroxylase 3 (GmABA8’-OH 3) and upregulates various drought-related genes. Overexpressing GmERF113 and GmPR10-1 increased the abscisic acid (ABA) content and reduced the expression of GmABA8’-OH3 in transgenic soybean plants and hairy roots, respectively. These results reveal that the GmERF113-GmPR10-1 pathway improves drought resistance and affects the ABA content in soybean, providing a theoretical basis for the molecular breeding of drought-tolerant soybean.
1. Introduction
Drought is a major abiotic stress that hampers the growth, development, and productivity of plants. In the context of rising global population and climate change, freshwater resources are becoming increasingly insufficient to meet agricultural needs [1]. In plants, drought leads to a series of changes at the morphological, physiological, and molecular levels. For example, the stomatal aperture is reduced to maintain the plant body moisture, and root growth is promoted to enhance the plant’s capacity to absorb water [2,3]. The activity of antioxidant enzymes such as superoxide dismutase (SOD), peroxidase (POD), catalase, and ascorbate peroxidase (APX) and the contents of osmolytes such as proline increase to help maintain the overall plant homeostasis [4,5,6]. The contents of phytohormones are altered, for example through increased abscisic acid (ABA) accumulation, to regulate stomatal movement, which in turn helps the plants tolerate the effects of extreme water deficit [7,8]. Numerous genes that are related to various signaling pathways, metabolic pathways, and functional proteins are induced to prevent or mitigate the damage that is caused by drought stress; these include genes that are related to ABA synthesis (e.g., NCED5) and catabolism (e.g., CYP707A) and to cell wall components (e.g., β-Galactosidase 10), late embryogenesis abundant (LEA) genes, and heat shock protein (HSP) genes [9,10,11,12].
Transcription factors such as bZIP, NAC, MYB, WRKY, and AP2/ERF family members play key regulatory roles in transcriptional reprogramming, thereby coordinately regulating the expression of many defense-related genes [13,14,15,16,17,18,19]. The AP2/ERF superfamily of plant-specific transcription factors (TFs) are characterized by a 60- to 70-amino-acid conserved AP2/ERF domain [20,21]. The AP2/ERF transcription factor superfamily, which has been identified in various species, is divided into different families based on the number of AP2 domains, including the AP2, ERF, DREB, and RAV families [20,21,22,23,24,25,26].
There is growing evidence that ethylene response factor (ERF) subfamily members are involved in plant responses and adaptation to abiotic stress. For example, overexpressing the AP2/ERF family gene BpERF13 in birch (Betula platyphylla) enhanced cold tolerance by upregulating CBF genes and mitigating reactive oxygen species (ROS)-induced damage [27]. Overexpressing the ethylene response factor gene ERF96 enhanced selenium tolerance in Arabidopsis thaliana [28]. ERF genes from maize (Zea mays L.; ZmERF), alfalfa (Medicago sativa L.; MsERF), Arabidopsis (Arabidopsis thaliana L.; AtERF1), peanut (Arachis hypogaea L.; AhERF019), wheat (Triticum aestivum L.; TaERF3), and soybean (Glycine max (L). Merr.; GmERF5) are all induced by drought [29,30,31,32,33,34,35]. Overexpressing OsERF71 and AtERF019 protected rice (Oryza sativa) and Arabidopsis from drought by altering the structure of rice roots, Arabidopsis stomatal aperture and cell wall permeability, respectively [11,36]. TaERF3-overexpressing wheat plants and OsLG3-overexpressing rice plants increased drought resistance by accumulating proline, chlorophyll, and increasing SOD and POD activities, respectively [32,37]. OsLG3 and OsERF71 also participated in the drought tolerance of rice by regulating the expression of ROS-scavenging and ABA-signaling-related genes [34,37], and the ERF protein MdERF38 responded to drought stress by interacting with a positive modulator of anthocyanin biosynthesis (MdMYB1) and facilitating its binding to its target genes in apple (Malus domestica) [38].
The AP2/ERF domain is required for the activity of AP2/ERF TFs, which it triggers by binding to cis-acting elements in their target genes, such as GCC-box motifs, DRE/CRT, or the TTG motifs in their promoters [32,39,40,41]. AP2/ERF TFs bind to the DRE/CRT or GCC-box motifs in stress-responsive genes to enhance resistance to abiotic stress [42,43]. For example, the AP2 transcription factor NtERF172 confers drought resistance in tobacco (Nicotiana tabacum) by binding to the DRE motif in the NtCAT promoter [44]. Overexpressing AtERF53 improved the drought tolerance in Arabidopsis by binding to the GCC-boxes and/or DRE elements in the promoters of its downstream genes, such as COR15B and P5CS1 [45].
Pathogenesis-related (PR) genes are known to contribute to plant resistance to biotic stresses [46,47,48,49,50,51,52,53]. However, there are also numerous studies showing that PR genes also confer tolerance to abiotic stresses. For example, ectopic overexpression of a salt-stress-induced PR Class 10 protein (PR10) gene from peanut afforded broad-spectrum abiotic stress tolerance in transgenic tobacco (Nicotiana tabacum) [54]. AhSIPR10 from peanut (Arachis hypogaea), RSOsPR10 from rice (Oryza sativa), and ThPR10 from Tamarix hispida are induced by drought and salt stress [54,55,56,57]. In rice, overexpressing the root-specific PR gene RSOsPR10 strongly improved drought tolerance [55] and overexpressing JIOsPR10 increased drought and salt stress tolerance [48]. SlNPR1 enhances the drought tolerance in tomato (Solanum lycopersicum) by regulating key drought-related genes, including SlGST, SlDHN, and SlDREB [58]. PR1-, PR2-, and PR5-overexpressing Arabidopsis plants showed enhanced drought tolerance. Finally, the transcription factor Di19 (drought-induced 19) directly upregulates PR1, PR2, and PR5 in response to drought stress in Arabidopsis [59].
To date, a total of 160 ERF genes have been identified in soybean [60], but the functions of most of these genes have not been resolved. We previously demonstrated that the ERF gene GmERF113 (GenBank accession no. XM_003548806, NCBI protein no. XP_003548854) could bind to the GCC-box and overexpression of GmERF113 transgenic soybean plants enhanced the expression of GmPR10-1 and resistance to P. sojae [61]. The objectives of this study were (i) to investigate whether GmERF113 is involved in drought stress through expression analysis; (ii) to verify the drought resistance function of GmERF113 through the phenotype and the determination of drought-related physiological indicators; and (iii) to resolve a molecular mechanism of GmERF113 in regulating drought resistance in soybean: the GmERF113-GmPR10-1 pathway enhances drought resistance and affects the ABA content in soybean, which will help us better understand the drought resistance function of GmERF113 and provide a theoretical basis for the molecular breeding of drought-tolerant soybean.
2. Results
2.1. GmERF113 Is Induced by Drought
To investigate whether GmERF113 is associated with drought stress, we performed quantitative reverse-transcription PCR to detect the expression of GmERF113 of soybean seedlings at the V2 stage that were treated with 20% PEG-6000 to induce drought (osmotic) stress. Under these PEG stress conditions, the transcript level of GmERF113 increased rapidly from 1 to 9 h (peak level 21.86-fold of the control) and then decreased to a low level by 24 h (Figure 1). These results suggest that GmERF113 is likely involved in the drought response in soybean.
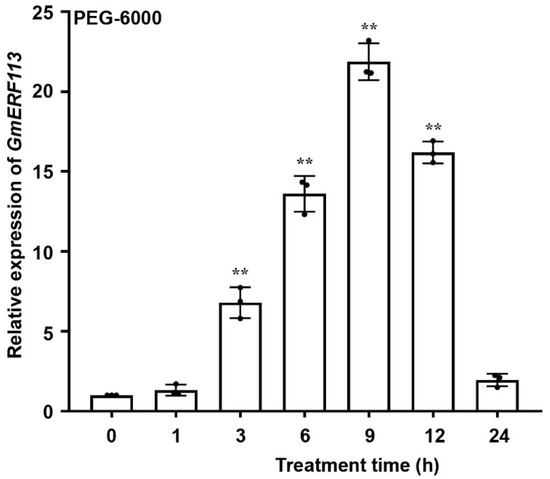
Figure 1.
GmERF113 is induced by drought. Expression patterns of GmERF113 of soybean seedlings at the V2 stage were examined with 20% PEG-6000 treatment. The first trifoliate leaf samples were collected at 0, 1, 3, 6, 9, 12, and 24 h. The reference soybean gene GmActin4 (GenBank accession no. AF049106) and GmTubulin4 (GenBank accession no. XM_003554060) were used as internal controls to normalize the data. The experiment was performed on three biological replicates, each with three technical replicates, and was statistically analyzed using Student’s t-test (** P < 0.01). The bars indicate the standard deviation of the mean.
2.2. Overexpression of GmERF113 Enhances Drought Tolerance in Transgenic Soybean Plants
To explore the role of GmERF113 in drought tolerance, we generated the overexpression vector 35S:GmERF113 and the RNA interference vector GmERF113-RNAi and used them to transform soybean plants via Agrobacterium-mediated transformation. A total of three independent GmERF113-OE T3 lines (GmERF113-OE15, GmERF113-25, and GmERF113-47) and three independent GmERF113-RNAi T3 lines (GmERF113-RNAi15, GmERF113-RNAi17, and GmERF113-RNAi21) were identified using bar strips and qRT-PCR (Figure S1A,C,D). Immunoblot analysis further confirmed that GmERF113 had been successfully introduced into and expressed in the GmERF113-OE soybean plants (Figure S1B).
Since plant water retention capacity contributes to drought resistance, we measured the fresh weights of the detached second trifoliate leaves of the wild-type (WT), GmERF113-OE, and GmERF113-RNAi T3 plants over 4 h following their detachment from the plants. Compared to that in the WT plants, water loss was slower in the leaves from GmERF113-OE plants and faster in the leaves from GmERF113-RNAi plants (Figure 2A). These results indicate that overexpressing GmERF113 enhances the water retention capacity of plants, thus improving their drought resistance.
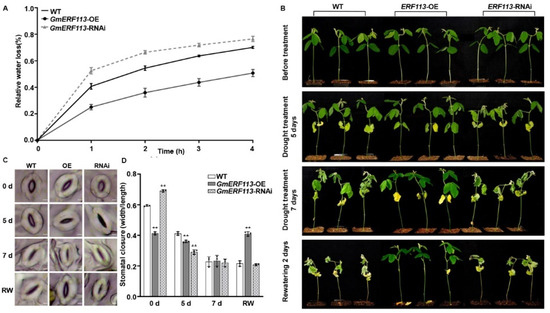
Figure 2.
GmERF113 is a positive regulator of plant response to drought. (A) The relative water loss of the detached leaves in wild-type (WT) plants, GmERF113-OE, and GmERF113-RNAi transgenic soybean plants. The second trifoliate leaves of soybean seedlings at the V3 stage were cut which were weighed every hour. The experiment was carried out in three biological replicates, each with three technical replicates. The bars indicate the standard deviation of the mean. (B) Phenotypes of WT, GmERF113-OE, and GmERF113-RNAi soybean plants that were exposed to drought stress for 0, 5, and 7 days and re-watered for 2 days. (C,D) Stomatal apertures of the second trifoliate leaves of WT, GmERF113-OE, and GmERF113-RNAi soybean plants that were treated with drought for 0, 5, or 7 d and re-watered for 2 days (C), and statistical analysis of the stomatal apertures of each line (bars = 20 μm) (D). The experiment was performed on three biological replicates, each with three technical replicates, and was statistically analyzed using Student’s t-test (** P < 0.01). The bars indicate the standard deviation of the mean.
To further validate the role of GmERF113 in drought response, we assessed the drought tolerance of the WT, GmERF131-OE, and GmERF113-RNAi T3 plants that were grown in soil. When the second trifoliate leaves of the WT, GmERF113-OE, and GmERF131-RNAi T3 plants were fully expanded (until V3 stage), we withheld watering for 7 days and resumed watering for 2 days. There was no difference in the phenotypes of the WT, GmERF113-OE, and GmERF113-RNAi plants under well-watered conditions. However, after 5 days without watering, most of the leaves of GmERF113-RNAi plants started to wilt and crumple, and the lowermost leaves of the WT and GmERF113-OE plants started to turn yellow and wilt, but the extent of leaf wilting was greater in the WT plants than in GmERF113-OE plants. After withholding watering for 7 days, all the leaves of GmERF113-RNAi plants were wilted and brittle, and all the leaves of the WT plants were wilted, while most of the leaves of GmERF113-OE plants were still expanded. After 2 days of re-watering, the WT and GmERF113-RNAi plants did not recover and were dead, whereas GmERF113-OE plants returned to normal growth (Figure 2B).
We also observed the stomata in the second trifoliate leaves of WT, GmERF113-OE, and GmERF113-RNAi plants during drought treatment. Before drought treatment, the stomatal aperture of GmERF113-OE plants was significantly smaller than that of the WT and GmERF113-RNAi plants (** P < 0.01) (Figure 2C,D). After 5 days of drought treatment, the stomatal apertures of GmERF113-OE and GmERF113-RNAi plants were significantly smaller than those of the WT plants (** P < 0.01). At 7 days of drought treatment, the stomata of WT, GmERF113-OE, and GmERF113-RNAi plants were all closed. After 2 days of rehydration, only the stomata of GmERF113-OE plants were open, while the stomata of WT and GmERF113-RNAi plants failed to open (Figure 2C). Together, these results suggest that GmERF113 plays a positive role in regulating the drought response.
2.3. Overexpressing GmERF113 Alters the Physiological and Biochemical Parameters of Transgenic Soybean Plants under Drought Stress
SOD, POD, and proline help to protect plants from oxidative damage under stress, and the MDA content reflects the extent of membrane lipid peroxidation [62,63,64,65,66,67]. Therefore, we evaluated these parameters to examine the adaptation of soybean plants to drought stress. Under drought treatment, the MDA content was significantly lower in the GmERF113-OE transgenic plants than in the WT plants, whereas the MDA content in the GmERF113-RNAi plants remained high and was significantly higher than that of the WT plants (Figure 3A). These results indicate that GmERF113-OE transgenic plants suffered less oxidative damage than the WT and GmERF113-RNAi plants.
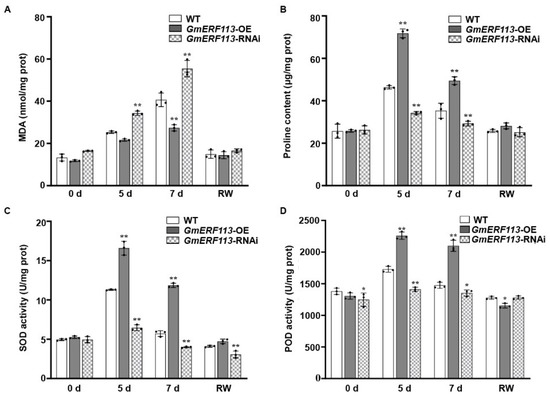
Figure 3.
Overexpression or silencing of GmERF113 alters drought-related parameters that are reflective in soybean plants. (A–D) Malondialdehyde content (MDA; A), proline content (B), and superoxide dismutase (SOD; C) and peroxidase (POD; D) activities of WT, GmERF113-OE, and GmERF113-RNAi soybean plants that were exposed to drought stress for 0, 5, and 7 days and re-watered for 2 days. The experiment was performed on three biological replicates, each with three technical replicates, and was statistically analyzed using Student’s t-test (* P < 0.05, ** P < 0.01). The bars indicate the standard deviation of the mean.
Proline is an important osmolyte that reduces the osmotic potential and alleviates dehydration stress in plants. The proline content of GmERF113-OE plants was more than 1.4-fold higher than that of the WT plants after 5 and 7 days of drought treatment, whereas the proline content of GmERF113-RNAi plants was approximately 26% and 17% lower than that of the WT plants after 5 and 7 days of drought treatment, respectively (Figure 3B).
SOD and POD scavenge superoxide radicals. Therefore, the SOD and POD activity can be used to analyze the drought stress response of plants. Under normal conditions, the SOD and POD activities of WT and transgenic lines were almost the same. Under drought stress conditions, however, the SOD and POD activities were significantly higher in GmERF113-OE than in the WT or GmERF113-RNAi plants (Figure 3C,D). These results suggest that overexpressing GmERF113 affects the accumulation of stress-related compounds such as proline and MDA, as well as the activities of stress-related enzymes such as SOD and POD, thereby improving drought tolerance.
2.4. GmERF113 Directly Activates the Expression of GmPR10-1
The accumulation of PR proteins during microbial infection or under abiotic stress conditions constitute a component of the plant innate immune response [68]. A GCC-box cis-element is present in the promoters of many PR genes. GmERF113 was previously shown to bind to the GCC-box and overexpression of GmERF113 transgenic soybean plants enhanced the expression of GmPR10-1 and resistance to P. sojae [61]. Here, we found that the GmPR10 promoter contains a GCC-box. To investigate whether GmERF113 directly binds to the GCC-box in the GmPR10 promoter, we performed ChIP assays with ERF1-myc seedlings at the V3 stage. The −295/−158 fragment (containing the GCCGCC site at positions −170 to −164) of the GmPR10-1 promoter in immunoprecipitated DNA from GmERF113-myc plants was significantly enriched (8.39-fold) compared with its abundance in the WT plants (Figure 4A,B).To further reveal the role of GmERF113 in regulating GmPR10-1 expression, we cloned the full-length GmPR10-1 promoter sequence (2068 bp) and fused it into the pGreenII 0800-LUC vector to generate the reporter construct p35S: REN-pGmPR10-1: LUC (Figure 4C). The recombinant vector p35S:GmERF113-myc, in which GmERF113 expression is driven by the 35S promoter, was used as the effector construct (Figure 4C). The reporter construct p35S: REN-pGmPR10-1: LUC and the p35S null effector vector, or the reporter construct p35S: REN-pGmPR10-1:LUC and the effector construct p35S:GmERF113-myc, were co-transfected into N. benthamiana leaves. After 3 days of co-transfection, the appropriate amounts of 1 mM D-luciferin were sprayed onto the transfected N. benthamiana leaves. The chemiluminescence signal in N. benthamiana leaves that were co-transfected with the reporter construct p35S: REN-pGmPR10-1:LUC and the effector construct p35S:GmERF113-myc was significantly stronger than that in the leaves harboring the reporter construct p35S:REN-pGmPR10-1:LUC and the effector construct p35S (Figure 4D). The results of the LUC/REN relative activity assay further indicated that GmERF113 directly promotes the expression of GmPR10-1 (Figure 4E). These results imply that GmERF113 binds to the GCC-box in the GmPR10-1 promoter and directly activates its expression.
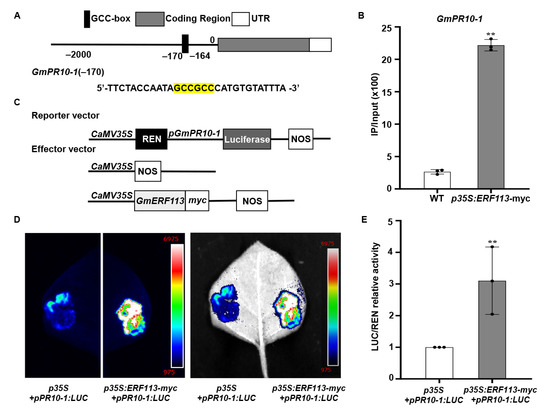
Figure 4.
GmPR10-1 was identified as a target of GmERF113. (A,B) ChIP analysis of GmERF113 binding to the promoter region of GmPR10-1. Chromatin from transgenic soybean plants expressing GmERF113-myc or the WT were immunoprecipitated with or without the anti-myc antibody. The precipitated chromatin fragments were analyzed with qPCR using a pair of specific ChIP-qPCR primers, which amplifies the GCC-box (highlighted in yellow) upstream of GmPR10-1, as indicated. One-tenth of the input (without antibody precipitation) of chromatin was analyzed and used as a control. A total of three biological replicates, each with three technical replicates, were averaged and statistically analyzed using Student’s t-test (** P < 0.01). The bars indicate the standard deviation of the mean. (C) Schematic representation of the reporter and effector constructs that were used in the dual luciferase assays. (D) The dual luciferase assays in tobacco leaves showing that GmERF113 activates the expression of GmPR10-1 by combining the GmPR10-1 promoter. Representative pictures were taken. (E) LUC/REN activity detection to verify GmERF113 activates the expression of GmPR10-1. The combination of the reporter construct (pGmPR10-1: LUC) and the blank effector construct [p35S] were used as the control. These experiments were performed on three biological replicates, each with three technical replicates, and were statistically analyzed using Student’s t-test (** P < 0.01). The bars indicate the standard deviation of the mean. “•” were scattered dots which can reflect the distribution of the data.
2.5. Overexpression of GmPR10-1 Improves Drought Tolerance in Soybean Hairy Roots
To further investigate the role of GmPR10-1 in drought response, we generated GmPR10-1 transgenic soybean hairy roots via Agrobacterium rhizogenes-mediated transformation. GUS staining and qRT-PCR analysis confirmed the generation of transgenic hairy roots that were overexpressing GmPR10-1 (Figure S2A,C). PCR analysis confirmed that the recombinant GmPR10-1 RNAi vector had been successfully transferred into the hairy roots, and qRT-PCR analysis showed that GmPR10-1 expression in GmPR10-1 RNAi transgenic hairy roots was successfully disrupted (Figure S2B,D).
We selected empty vector (EV) control, GmPR10-1-OE, and GmPR10-1-RNAi transgenic soybean composite plants with hairy roots of the same length, transferred them to fresh vermiculite, incubated them for 3 days under normal watering conditions, and then withheld watering. After 5 days of drought treatment, all the leaves of the GmPR10-1 RNAi transgenic composite plants were severely wilted, whereas the second and third trifoliate leaves of the EV composite plants and all the leaves of the GmPR10-1- OE transgenic composite plants showed no obvious wilting. After 7 days of drought treatment, the EV composite plants showed severe leaf dehydration and the GmPR10-1 RNAi composite plants showed more severe leaf abscission compared to the GmPR10-1- OE composite plants. We then resumed watering, and 2 days after the start of rehydration, only the GmPR10-1-OE composite plants regained vitality, whereas both the EV and GmPR10-1-RNAi composite plants were dead (Figure 5B).
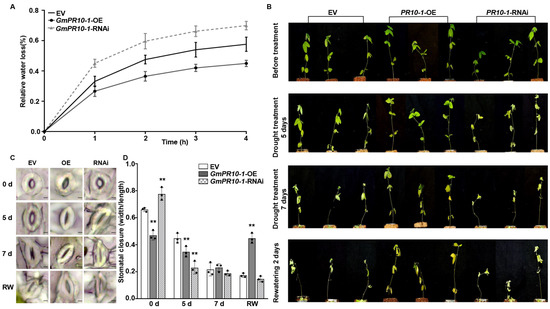
Figure 5.
GmPR10-1 improves the drought tolerance in composite soybean plants with transgenic hairy roots. (A) The relative water loss of detached leaves (the second trifoliate leaves from the top) from EV, GmPR10-1-OE, and GmPR10-1-RNAi composite soybean plants with transgenic hairy roots. Transgenic soybean hairy roots of the same length (~10 cm) were selected, transferred to new pots (soil moisture contents in the pots were kept constant), and incubated for 3 days before the start of the drought treatment. (B) Phenotypes of EV, GmPR10-1-OE, and GmPR10-1-RNAi composite soybean plants with transgenic hairy roots that were exposed to drought stress for 0, 5, and 7 days and re-watered for 2 days. (C,D) Stomatal aperture of the second trifoliate leaves from the top of EV, GmPR10-1-OE, and GmPR10-1-RNAi composite soybean plants with transgenic hairy roots that were treated with drought for 0, 5, and 7 days and re-watered for 2 days (bars = 20 μm). (C), and statistical analysis of the stomatal apertures of each line. (D). The experiments were performed on three biological replicates, each with three technical replicates, and were statistically analyzed using Student’s t-test (* P < 0.05, ** P < 0.01). The bars indicate the standard deviation of the mean.
We then measured the water loss rate and the stomatal aperture of the second trifoliate leaves from the top in EV, GmPR10-1-OE, and GmPR10-1-RNAi composite soybean plants with transgenic hairy roots. Compared to that in the EV control plants, the rate of water loss rate from the leaves was reduced in the leaves of the GmPR10-1-OE plants and elevated in the leaves of the GmPR10-1-RNAi plants (Figure 5A). Before the drought treatment, the stomatal apertures of the GmPR10-1-OE composite plants were significantly smaller than those of the EV and GmPR10-1-RNAi plants (** P < 0.01). After 5 days of drought treatment, the stomatal apertures of GmPR10-1-OE and GmPR10-1-RNAi plants (** P < 0.01) were significantly smaller than those of the EV control plants. After 7 days of drought treatment, the stomata of the EV, GmPR10-1-OE, and GmPR10-1-RNAi plants were all closed. After 2 days of rehydration, only the stomata of GmPR10-1-OE composite plants were open, whereas the EV and GmPR10-1-RNAi stomata failed to open (Figure 5C,D). These results suggest that overexpressing GmPR10-1 enhances drought tolerance in transgenic soybean composite plants with transgenic hairy root.
To further verify the role of GmPR10-1 in drought resistance, we measured the changes in the physiological and biochemical parameters of the leaves of composite plants. Before drought treatment, there were no significant differences in the MDA and proline contents or SOD and POD activities in the leaves of EV, GmPR10-1-OE, and GmPR10-1-RNAi composite plants. After 5 days and 7 days of drought treatment, compared to the EV and GmPR10-1-RNAi composite plants, the MDA content (Figure 6A) in the leaves of GmPR10-1-OE composite plants was significantly reduced, while the proline content and the SOD and POD activities were significantly increased (Figure 6B–D). Together, these results demonstrate that overexpressing GmPR10-1 improves the drought tolerance of soybean composite plants.

Figure 6.
Overexpression or silencing of GmPR10-1 alters the drought-related parameters in composite soybean plants. (A–D) Malondialdehyde content (MDA; A), proline content (B), and superoxide dismutase (SOD; C) and peroxidase (POD; D) activities of EV, GmPR10-1-OE, and GmPR10-1-RNAi composite soybean plants with transgenic hairy roots that were exposed to drought stress for 0, 5, and 7 days and re-watered for 2 days. The second trifoliate from the top was chosen to measure physiological indicators. These experiments were performed on three biological replicates, each with three technical replicates, and were statistically analyzed using Student’s t-test (* P < 0.05, ** P < 0.01). The bars indicate the standard deviation of the mean.
2.6. GmERF113 Differentially Regulates Genes Involved in Drought Response
To further investigate the drought resistance mechanism of GmERF113, we performed comparative transcriptome analysis using leaves from plants of three GmERF113-OE lines and WT plants. Based on the RNA-seq data, we identified 360 differential expression genes (DEGs) in the GmERF113-OE plants compared to the control, including 57 upregulated and 303 downregulated genes (Figure 7A,B). To examine the functions of these DEGs, we performed GO analysis. All of the DEGs were assigned to three major categories: biological processes, cellular components, and molecular functions. Most DEGs in the biological process category were enriched in cellular processes, metabolic processes, responses to stimuli, and biological regulation. Most of the DEGs in the cellular component category were enriched in cells, organelles, and membranes. Most of the DEGs in the molecular function category were associated with binding, catalytic activity, and transporter activity (Figure 7C). Some of the DEGs that are associated with processes that are involved in drought stress response were upregulated more than 1.4-fold, such as genes encoding galactinol-sucrose galactosyltransferase 2-like, ABC transporter C family member 9-like, beta-galactosidase 1-like, tyrosine aminotransferase, UDP-glycosyltransferase 76E11-like, calcium-binding protein CML38-like and peroxidase 15-like, and zinc finger CCCH domain-containing protein 20-like. Some were genes encoding proteins that participate in the drought stress response by affecting the ABA pathway, such as ABA 8’-hydroxylase 3, protein phosphatase 2C 37-like, and E3 ubiquitin-protein ligase RGLG1-like (Table S2). Based on these results, we suggest that GmERF113 plays an important role in the drought stress response by regulating various drought-related genes and may be involved in the ABA-related pathway.
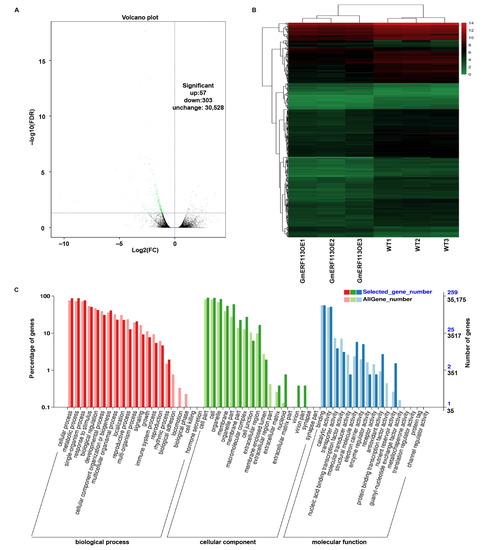
Figure 7.
Transcriptomic analysis of gene expression profiles in response to GmERF113 overexpression. (A) Volcano plots of significantly differentially expressed genes in GmERF113-OE vs. wild-type soybean plants after the RNA-seq analysis. (B) Heat map of significantly differentially expressed genes between the WT and GmERF113-OE transgenic soybean plants, as determined using an RNA-seq analysis. Using a false discovery rate < 0.05 and a fold change ≥1 as the screening criteria, a total of 360 differentially expressed genes (DEGs) were identified. The scale bar indicates the fold changes (log2 values). (C) Gene Ontology functional classification of the differentially expressed genes. The differentially expressed genes were placed into the three main GO categories: biological process, cellular component, and molecular function.
2.7. Overexpression of GmERF113 and GmPR10-1 Influences ABA Content and ABA-Related Gene Expression
To further investigate whether the role of GmERF113 in drought tolerance is associated with the ABA pathway, we measured the ABA contents of the WT, GmERF113-OE, and GmERF113-RNAi plants under normal conditions and drought treatment. We also analyzed the expression of genes that function in drought tolerance and genes that are related to the ABA signaling pathway in WT, GmERF113-OE, and GmERF113-RNAi plants. The ABA levels were significantly higher in GmERF113-OE plants than in the WT and GmERF113-RNAi plants under both the drought treatment and normal conditions (Figure 8A). Moreover, the expression level of GmABA8’-OH 3 (encoding ABA 8’-hydroxylase) was markedly higher in GmERF113-RNAi plants than in the WT and GmERF113-OE plants, and the expression levels of ABA signaling genes GmRGLG1 and GmPP2C37 were significantly higher in the GmERF113-OE plants than in the WT and GmERF113-RNAi plants (Figure 8B).
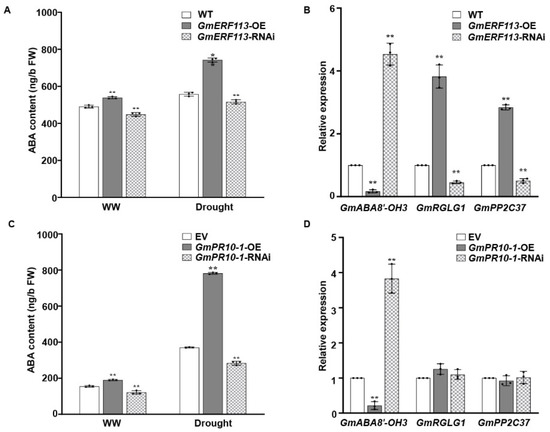
Figure 8.
GmERF113 and GmPR10-1 function in ABA responses. (A) The ABA contents of GmERF113 transgenic soybean plants under normal conditions and drought treatment. (B) The relative expression of three ABA-related genes that were identified by RNA-seq analysis in GmERF113 transgenic soybean plants. (C) The ABA contents of GmPR10-1 transgenic soybean hairy roots under normal conditions and drought treatment. (D) The relative expression of three ABA-related genes that were identified by RNA-seq analysis in GmPR10-1 transgenic soybean hairy roots. The reference soybean gene GmActin4 and GmTubulin4 were used as an internal control to normalize the data. These experiments were performed on three biological replicates, each with three technical replicates, and were statistically analyzed using Student’s t-test (* P < 0.05, ** P < 0.01). The bars indicate the standard deviation of the mean.
Finally, to investigate whether GmPR10-1, the direct downstream target gene of GmERF113, is also involved in the ABA pathway, we performed the same assay as above using transgenic hairy roots from EV, GmPR10-1-OE, and GmPR10-1-RNAi plants. The ABA levels were significantly higher in GmPR10-1-OE than in either EV or GmPR10-1-RNAi transgenic hairy roots under both drought treatment and normal conditions (Figure 8C). GmABA8’-OH 3 was expressed at markedly lower levels in GmPR10-1-OE vs. EV hairy roots, whereas GmRGLG1 and GmPP2C37 expression were not different in GmPR10-1-OE vs. EV and GmPR10-1-RNAi hairy roots (Figure 8D). These results suggest that overexpressing GmERF113 and GmPR10-1 increases the ABA contents in soybean plants and hairy roots by affecting the expression of the ABA 8’-hydroxylase gene GmABA8’-OH 3, thus improving drought resistance.
3. Discussion
3.1. GmERF113 Contributes to Soybean Response to Drought Stress
Soybean is the most important legume crop globally, representing an important source of edible oils and proteins for human consumption [69]. Drought can cause up to 40–60% losses in soybean production worldwide [70]. Among the DEGs in soybean leaf tissue under drought stress, ERF genes show the most significant differential expression, followed by bHLH, MYB, NAC, and WRKY genes [71]. Moreover, there was some evidence that showed that ERF transcription factors were also involved in regulating drought responses in addition to biotic stresses [72,73,74]. For example, the transgenic Arabidopsis overexpressing SlERF84 from tomato (Solanum lycopersicum) not only showed decreased resistance to the bacterial speck pathogen, Pseudomonas syringae pv. tomato DC3000, but also displayed increased tolerance to drought stress [74]. Several studies have demonstrated that the ectopic expression of GmERF3, GmERF4, and GmERF6 improved drought resistance in tobacco or Arabidopsis and overexpressing GmERF75 improved the resistance of soybean hairy roots to osmotic stress [60,75,76,77]. Similarly, we have previously demonstrated that GmERF113 enhanced resistance to P. sojae in soybean [61]. Then, we found that GmERF113 expression was induced by PEG treatment (Figure 1), suggesting that GmERF113 plays a critical role in the drought stress response. Under drought stress, transgenic soybean plants overexpressing GmERF113 (GmERF113-OE) showed significantly slower water loss in the leaves than WT plants and plants with RNAi silencing of the gene (GmERF113-RNAi plants) (Figure 2A). The leaves of GmERF113-OE plants also exhibited less wilting under drought treatment and stronger recovery after rehydration than those from WT and GmERF113-RNAi plants (Figure 2B–D). In addition, GmERF113-OE transgenic plants possessed higher SOD activity, POD activity, and proline content (all protective against drought stress), as well as lower MDA content (indicative of drought damage), compared to the WT and GmERF113-RNAi plants (Figure 3A–D), providing further evidence that overexpressing GmERF113 improves plant resistance to drought stress. Collectively, these results demonstrate that GmERF113 contributes to the response of soybean to drought stress.
3.2. GmERF113 Increases Soybean Drought Resistance by Directly Activating GmPR10-1 and Affecting the Expression of Drought-Related Genes
Pathogenesis-related (PR) genes, a class of defense-related genes, play important roles in plant resistance to biotic and abiotic stresses [51,56,78,79,80,81]. PR10 gene expression is induced in many plants by drought and salt stress [48,82,83,84]. Here, we isolated GmPR10-1 from ‘Dongnong 50’ and produced composite soybean plants with GmPR10-1 transgenic hairy roots by Agrobacterium rhizogenes-mediated transformation. The degree of leaf wilting was lower in the composite plants with GmPR10-1-OE transgenic hairy roots compared to the EV control and GmPR10-1-RNAi lines under drought treatment. After rehydration, the GmPR10-1-OE lines remained viable, whereas the EV and GmPR10-1-RNAi lines were dead, with severe leaf shedding and stem shrinkage (Figure 5B–D). In addition, the SOD and POD activity and proline content in the leaves of GmPR10-1-OE plants were significantly higher than those of the EV and GmPR10-1-RNAi plants, whereas the MDA content showed the opposite trend (Figure 6A–D). Together, these results confirm the role of GmPR10-1 in drought tolerance in soybean, which is consistent with the finding that some PR10 proteins are involved in drought tolerance [48,54,55,57].
We previously found that GmERF113 binds to the GCC-boxes [61]. (Zhao et al., 2017). Here, we demonstrated that GmERF113 binds directly to the GCC-box in the GmPR10-1 promoter by ChIP-qPCR and showed that GmERF113 promotes the expression of GmPR10-1 in a dual luciferase assay. Our RNA-seq results indicated that overexpressing GmERF113 led to the altered expression of several drought-stress-related genes, including genes encoding galactinol-sucrose galactosyltransferase 2-like, ABC transporter C family member 9-like, beta-galactosidase 1-like, tyrosine aminotransferase, UDP-glycosyltransferase 76E11-like, calcium-binding protein CML38-like and peroxidase 15-like, and zinc finger CCCH domain-containing protein 20-like (Table S2). Galactitol-sucrose galactosyltransferase 2 interacts with the MYB gene 1R-MYB in chickpea (Cicer arietinum L.), which is significantly induced by drought; these MYB transcription factors are jointly involved in the drought tolerance pathway [85]. The guard cell plasma membrane ABCC-type ABC transporter protein AtMRP4 is involved in regulating stomatal opening and confers drought tolerance in Arabidopsis [86]. Beta-galactosidase and peroxidase function in abiotic stress responses, and beta-galactosidase is a glycosyl hydrolase that is involved in cell wall modification that plays an important role in plant development and adaptation to environmental stress [87,88]. Ectopic expression in Arabidopsis of the tyrosine aminotransferase gene MdTAT2 from apple (Malus domestica) or the overexpression of this gene in apple callus enhance tolerance to drought and osmotic stress [89]. Arabidopsis plants overexpressing AtUGT76C2 (UDP-glycosyltransferase 76C2) showed enhanced drought tolerance, and the stress-inducible genes DREB2A, RD22, RD29B, LEA, COR47, and KIN1 were significantly upregulated in response to AtUGT76C2 overexpression [90]. An Arabidopsis calmodulin-like (CML) protein with calcium-binding activity was shown to be involved in regulating ABA signaling and drought stress tolerance in guard cells [91]. The CCCH zinc finger protein OsC3H10 is involved in regulating the drought tolerance pathway by modulating the expression of stress-related genes in rice [92]. These findings suggest that GmERF113 increases drought resistance in soybean by directly activating GmPR10-1 and affecting the expression of drought-related genes.
3.3. GmERF113 Might Participate in ABA-Mediated Regulation of Drought Response
The ERF transcription factor genes GmERF3, GmERF4, GmERF5, GmERF6, GmERF7, and GmERF75 are induced by ABA in soybean [35,75,76,77,93]. ABA plays a crucial role in plant development and adaptation to abiotic stress [42,94,95]. Several ABA-related genes regulate stomatal aperture and water potential by affecting the ABA contents to improve the drought resistance of plants [96,97,98]. ABA 8′-hydroxylase is the key enzyme in the oxidative catabolism of ABA. Arabidopsis plants that were treated with Abscinazole-E3M, an inhibitor of ABA 8′-hydroxylase, improved drought resistance [99]. We previously demonstrated that GmERF113 is induced by ABA [61]. Here, we showed that GmERF113-OE plants had significantly smaller stomatal apertures than WT plants under normal conditions (** P < 0.01; Figure 2C,D). We propose that GmERF113 negatively regulates the expression of the ABA 8′-hydroxylase gene, GmABA8′-OH 3 (Table S2, Figure 8B), which in turn leads to significantly higher ABA contents in GmERF113-OE plants than in the WT plants, thereby reducing stomatal opening size in GmERF113-OE plants (Figure 8A).
In addition, RNA-seq analysis revealed the upregulation of several genes encoding proteins that may be involved in the ABA signaling pathway, such as protein phosphatase 2C 37-like and E3 ubiquitin-protein ligase RGLG1-like (Table S1). Type 2C protein phosphatases (PP2Cs) negatively regulate ABA signaling and play diverse roles in plant development responses to various stresses [9,100,101]. Arabidopsis ABA promotes the degradation of PP2CA via the E3 ligase RGLG1 E3, and the RING E3 ligase RGLG2 interacts with AtERF53 to negatively regulate the drought stress response in Arabidopsis [45,102]. These findings suggest that GmERF113 functions in ABA signaling by regulating the genes that are related to the ABA signaling pathway.
Finally, in many important plant pathological systems, phytohormones are essential for the activation of signaling during disease defense responses [103,104,105,106,107]. PR4 and PR10 were significantly upregulated in both the leaf and stem tissues of highly resistant lentil varieties after treatment with ABA, suggesting that ABA is a key hormonal activator of necrotrophic fungal defense response signaling in lentil [108]. Several PR10 genes are also induced by ABA [50,109]. Similarly, we found that the stomatal apertures of GmPR10-1-OE soybean hairy root composite plants were significantly smaller than those of WT plants under normal conditions (** P < 0.01; Figure 5C,D). The ABA content was significantly higher in PR10-1-OE transgenic hairy roots than in EV and PR10-1-RNAi transgenic hairy roots under both normal and drought treatment, and the expression of GmABA8′-OH 3 was significantly lower in PR10-1-OE vs. EV and PR10-1-RNAi transgenic hairy roots (Figure 8C,D).
Based on these results, we propose a model that explains the mechanisms of GmERF113 drought stress (Figure 9). When soybean plants are subjected to drought stress, GmERF113 is rapidly activated and transcribed, and then GmERF113 activates the expression of PR10-1 by binding to the GCC-box in the PR10-1 promoter, thereby enhancing the drought resistance of soybean plants. In addition, GmERF113 promotes the expression of two genes which are in the ABA signaling pathway, GmPP2C37 and GmRGLG1, also both GmERF113 and GmPR10 decrease the expression of an ABA catabolic gene, GmABA8′-OH 3, thereby the ABA content in the plant can be increased. The increased ABA level further promotes the closure of stomata and thus improves the drought tolerance of soybean plants.

Figure 9.
A molecular model of the GmERF113 in the soybean response to drought stress. When soybean plants are subjected to drought stress, GmERF113 is rapidly activated and transcribed, and then GmERF113 activates the expression of PR10-1 by binding to the GCC-box in the PR10-1 promoter, thereby enhancing the drought resistance of soybean plants. In addition, GmERF113 promotes the expression of two genes which are in the ABA signaling pathway, GmPP2C37 and GmRGLG1, also both GmERF113 and GmPR10 decrease the expression of an ABA catabolic gene, GmABA8′-OH 3, thereby the ABA content in the plants can be increased. The increased ABA level further promotes the closure of stomata and thus improves the drought tolerance of soybean plants.
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Treatments
‘Dongnong 50’, a popular soybean cultivar in Heilongjiang, China, was obtained from the Key Laboratory of Soybean Biology at the Chinese Ministry of Education, Harbin, China, and used for expression analysis, gene transformation experiments, and gene isolation. The seeds were sown in pots containing a vermiculite: soil (1:1) mixture and placed in a growth chamber under a 16 h light/8 h dark photoperiod with 70% relative humidity at 25 °C until the V2 stage [110]. The seedlings were treated with 20% PEG-6000, and the first trifoliate leaves were collected at 0, 1, 3, 6, 9, 12, and 24 h [111]. All the samples were immediately frozen in liquid nitrogen and stored at −80 °C prior to analysis. For the dual-luciferase assays, Nicotiana benthamiana seeds were grown in half-strength Murashige and Skoog (1/2MS) medium for 7 days, and the 7-day-old seedlings were transferred to sterile soil and cultured for up to 21 days (vermiculite:soil, 1:1). The plants were maintained at 25 °C under a 16 h light/8 h dark photoperiod.
4.2. RNA Isolation and Quantitative Reverse-Transcription PCR
The total RNA was extracted from the samples using TRIzol reagent (Invitrogen, Shanghai, China) following the manufacturer’s protocol. Reverse transcription was conducted using a ReverTra Ace qPCR RT Kit (TOYOBO, Japan). qRT-PCR was employed to measure the gene expression levels using a real-time RT-PCR kit (Toyobo, Osaka, Japan) with a LightCycler 96 System (Roche, Pleasant, CA, USA). The soybean housekeeping gene GmActin4 (GenBank accession no. AF049106) and GmTubulin4 (GenBank accession no. XM_003554060) were used as internal reference for normalization of qRT-PCR CT values. The qPCR analyses were performed on three biological replicates, each with three technical replicates. All the qPCR primers that were used in this study are listed in Supplementary Table S1.
4.3. Construction of the GmERF113 Recombinant Plasmids, Genetic Transformation of Soybean
The GmERF113-PMD18T recombinant vector was constructed in our previous study [61]. To generate the 35S:GmERF113 construct, the full-length coding sequence of GmERF113 was amplified with specific primers (Supplementary Table S1) and inserted into the NcoI and BglII cleavage sites of plant expression vector pCAMBIA3301 with the Bar gene and 4× Myc tag as the selectable marker. To obtain the GmERF113 RNAi vector, a 325-bp fragment of GmERF113 was amplified using the primer set ERF1 RNAi-F/R (Supplementary Table S1). After the fragment was inserted into the BamHI and XbaI sites of the pFGC5941 vector, the reverse repeat fragment was cloned into the XhoI and NcoI sites of the vector with the Bar gene as the selectable marker [112].
The overexpression construct and the RNAi silencing construct were transferred into Agrobacterium tumefaciens strain LBA4404 and used to transform the soybean as described by Paz et al. [113]. GmERF113-OE transgenic plants (T3, self-crossing) were identified using a QuickStix Kit for Bar (EnviroLogix, Portland, ME, USA), by qPCR and immunoblotting using Myc antibody (Abmart, code number M20002M). GmERF113-RNAi T3 transgenic plants were tested using bar strips and qPCR.
4.4. Drought Treatment, Measuring Water Loss Rate, and Stomatal Aperture
A total of three independent GmERF113-overexpression T3 seedlings, three independent GmERF113-RNAi T3 seedlings, and three wild-type (WT) seedlings were grown in the same volume of soil (1:1 vermiculite: soil) and incubated in an incubator at 25 °C under a long daylight cycle (16 h light/8 h dark) and 60% relative humidity until V3 stage [110].
To measure water loss, the second trifoliate leaf was cut from the plant, weighed immediately, and periodically at one-hour intervals following the procedure that was described by Wang et al. [114].
For the drought treatment and the measurement of the stomatal aperture assays, the WT, GmERF113-OE, and GmERF113-RNAi seedlings were withheld watering for 7 days. The plants were then re-watered for 2 days. The seedling phenotypes were photographed after 0, 5, and 7 days of drought treatment and after 2 days of rehydration using a camera (Canon IXUS 860IS). The second trifoliate leaves of plants were collected at the same time point as mentioned above. The lower epidermis of the leaves was immediately taped and placed under a microscope to observe the morphology of the stomata. The stomata apertures in the images were measured using ViewPoint software. The stomatal aperture results are reflected by the width/length of the stomata.
4.5. Measurement of Proline Content, MDA Content, and Superoxide Dismutase and Peroxidase Activities
The second trifoliate leaves of GmERF113 transgenic and WT soybean plants were collected after 0, 5, and 7 days of drought treatment and after 2 days of rehydration to measure the physiological indexes. Malondialdehyde (MDA) content, proline content, and the superoxide dismutase (SOD) and peroxidase (POD) activities were measured using an MDA Assay kit (Comin, MDA-2-Y, Suzhou, China), Proline Assay kit (Comin, PRO-2-Y, Suzhou, China), SOD Assay kit (Comin, SOD-2-Y, Suzhou, China), and POD Assay kit (Comin, POD-2-Y, Suzhou, China), respectively, according to the manufacturers’ protocols. Each experiment included three replicates, each with three technical replicates.
4.6. Chromatin Immunoprecipitation (ChIP) Assay
For the ChIP assays, WT and p35S:GmERF113-myc transgenic plants were subjected to chromatin extraction and immunoprecipitation as described by Saleh et al. [115]. The second and third trifoliate leaves of the WT and 35S-GmERF113-myc transgenic plants were harvested at approximately 1 g, respectively. The leaves were fixed in formaldehyde solution for 30 min under a vacuum, and the reaction was terminated by adding 0.15 M of glycine powder. The chromatin complex was isolated and sonicated to about 500 bp fragments. The protein-DNA crosslinks were incubated with anti-Myc-Tag Mouse mAb (Agarose Conjugated) (Abmart, Shanghai, China, code number M20012). The combined DNAs that were eluted, purified, dissolved, and analyzed by quantitative PCR. The ChIP-qPCR results are reported as relative binding units (IP/input). The primers that were used are listed in Supplementary Table S1.
4.7. Cloning of the GmPR10-1 Promoter and Dual-Luciferase Assay
The GmPR10-1 promoter sequence was isolated from the genome of ‘Dongnong 50’ and cloned into the pGreenII 0800-LUC vector, which was used as a reporter. The primers pGmPR10-1F/R that were used to clone the GmPR10-1 promoter are listed in Supplementary Table S1. p35S:GmERF113-myc was used as an effector construct. The effector and reporter constructs were co-transfected into healthy leaves of 21-day-old N. benthamiana plants by agroinfiltration. After infiltration, the plants were incubated in the dark for 3 days. The leaves were then sprayed with luciferin (1 mM luciferin and 0.01% Triton X-100) and incubated in the dark for 20 min before being cut off and placed on Chemiluminescence imaging (Tanon 5200) for photographing. Luciferase (LUC) and the Renilla luciferase (REN) activities of the leaf samples were determined using a commercial kit (Promega; PR-E1910). The 35S promoter- driven Renilla luciferase (REN) gene in the pGreenII 0800-LUC vector was used as an internal control. The LUC activity was reacted by LUC/REN ratio. The data are the averages of at least three independent replicates, each with three technical replicates.
4.8. Agrobacterium Rhizogenes-Mediated Transformation of Soybean Hairy Roots
The full-length cDNA (termed GmPR10-1) was isolated from the cDNA of ‘Dongnong 50′ by RT-PCR using primers GmPR10-1-F/R (Supplementary Table S1). To generate the 35S:GmPR10-1 construct, the Bar gene in the left arm of the plant expression vector pCAMBIA3301 was replaced with the full-length coding sequence of GmPR10-1 that was amplified with specific primers (Supplementary Table S1), and the GUS gene was used as a selectable marker. To generate the GmPR10-1 RNAi vector, a 300-bp fragment of GmPR10-1 was amplified using the GmPR10-1 RNAi-F/R primer set (Supplementary Table S1) and inserted in forward orientation into the BamHI and XbaI sites of pFGC5941 and in reverse orientation into the XhoI and NcoI sites of this vector. Transgenic soybean hairy roots were generated by Agrobacterium rhizogenes-mediated transformation following the methods as described by Kereszt et al. [116].
4.9. RNA-Seq Analysis
RNA was extracted from the leaves of three independent 30-day-old GmERF113-OE transgenic soybean plants and three WT plants under non-stress conditions using TRIzol reagent and a Spectrum Plant Total RNA Kit (Sigma-Aldrich, St. Louis, MO, USA). Sequencing libraries were generated using a NEB Next Ultra RNA Library Prep Kit for Illumina (NEB, Ipswich, MA, USA) following the manufacturer’s recommendations, and index codes were added to each sample. RNA-seq was performed using the Illumina HiSeq 2500 platform. Each sample generated more than 6 gigabytes of data. The sequencing data were compared with the soybean reference genome (https://phytozome.jgi.doe.gov/pz/portal.html, accessed on 20 December 2019) using TopHat2 [117] to obtain positional information on the reference gene and information about the characteristics of the sequence. Differential expression analysis between the sample groups was performed using DESeq [118] to identify differentially expressed genes (DEGs) between the two biological conditions. During DEG detection, fold change ≥ 1, FDR < 0.05 was used as a screening standard. The functions of the DEGs were annotated by GO enrichment analysis [119]. The experiment was performed on three biological replicates, each with three technical replicates.
4.10. Measuring ABA Contents
The ABA contents were measured as described previously [120] using a High Performance Liquid Chromatography kit (COMIN, Suzhou, China) according to the manufacturer’s protocol.
4.11. Statistical Analysis
The experiments were performed on three biological replicates, each with three technical replicates, and the results were statistically analyzed using Student’s t-test after test for normal distribution with the Shapiro–Wilk tests [121]. IBM SPSS26 package for Windows (IBM, New York, NY, USA) was used for the Shapiro–Wilk tests. A difference was considered to be statistically significant when * P < 0.05 or ** P < 0.01. The bars indicate the standard deviation of the mean.
5. Conclusions
In this study, we found that GmERF113 positively regulates the drought response in soybean. We further confirmed that GmERF113 activates the expression of the downstream target gene GmPR10-1. The GmERF113-GmPR10-1 pathway improves drought resistance and affects the ABA content in soybean. In addition, we demonstrated that GmERF113 involves the drought response by positively regulating various drought-related genes that are involved in regulating stomatal opening, antioxidant pathways, cell wall modification, and ABA signaling pathways. Our findings indicate the role of GmERF113 in the regulating mechanism of soybean response to drought stress and provide a theoretical basis for the molecular breeding of drought-resistant soybean.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms23158159/s1.
Author Contributions
P.X., S.Z. and J.W. designed the experiments. X.F., J.M., F.G., D.Q., M.Z. and L.W. performed the experiments. C.Z., B.S., S.L., S.H. and Y.L. analyzed the data. X.F., P.X., S.Z. and J.W. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NSFC Projects (31671719, 31971972), Natural Science Foundation of Heilongjiang Province (ZD2019C001), Key Research and Development Program of Heilongjiang Province (GX18B032).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request. Raw RNA sequencing data are available at the NCBI Sequence ReadArchive (SRA) under accession PRJNA760544 and can be accessed via the following link: https://dataview.ncbi.nlm.nih.gov/object/PRJNA760544?reviewer=1jlefrlk5q0np5tv574n095a38, accessed on 5 September 2021. Genes sequences in this research were obtained from National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/, accessed on 5 January 2017) and Phytozome (https://phytozome.jgi.doe.gov/, accessed on 16 May 2020). The accession numbers of genes are as follows: GmERF113 (XM_003548806), GmPR10-1 (NM_001251335), GmActin4 (AF049106), GmTubulin4 (XM_003554060), Galactinol-sucrose galactosyltransferase 2-like (Glyma.03G137900), ABC transporter C family member 9-like (Glyma.03G101000), Beta-galactosidase 1-like (Glyma.08G193500), Tyrosine aminotransferase (Glyma.06G235500), UDP-glycosyltransferase 76E11-like (Glyma.12G058300), Calcium-binding protein CML38-like (Glyma.04G194800), Peroxidase 15-like (Glyma.15G128800), Zinc finger CCCH domain-containing protein 20-like (Glyma.08G031400), ABA 8’-hydroxylase 3 (Glyma.16G110700), Protein phosphatase 2C 37-like (Glyma.18G035000), E3 ubiquitin-protein ligase RGLG1-like (Glyma.06G155100).
Acknowledgments
We thank the support from the Outstanding Talents and Innovative Team of Agricultural Scientific Research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gupta, A.; Rico-Medina, A.; Cao-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, H.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Zhou, Y.Y.; Zhang, Y.; Wang, X.W.; Han, X.; An, Y.; Lin, S.W.; Shen, C.; Wen, J.; Liu, C.; Yin, W.; et al. The root-specific NF-Y family transcription factor, PdNF-YB21, positively regulates root growth and drought resistance by ABA-mediated IAA transport in Populus. New Phytol. 2020, 227, 407–426. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van, B.F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Fujita, M. Roles of osmolytes in plant adaptation to drought and salinity. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: New Delhi, India, 2016; pp. 37–68. [Google Scholar] [CrossRef]
- Furlan, A.L.; Bianucci, E.; Giordano, W.; Castro, S.; Becker, D.F. Proline metabolic dynamics and implications in drought tolerance of peanut plants. Plant Physiol. Biochem. 2020, 151, 566–578. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Lynch, T.J. Abscisic acid inhibition of radicle emergence but not seedling growth is suppressed by sugars. Plant Physiol. 2000, 122, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Molina, L.; Mongrand, S.; Chua, N.H. A postgermination developmental arrest checkpoint is mediated by abscisic acid and requires the ABI5 transcription factor in Arabidopsis. Proc. Natl. Acad. Sci. USA 2001, 98, 4782–4787. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Tang, W.; Wang, C.; Ge, L.; Sun, J.; Qi, X.; He, Z.; Zhou, Y.; Chen, J.; Xu, Z.; et al. SiMYB56 confers drought stress tolerance in transgenic rice by regulating lignin biosynthesis and ABA signaling pathway. Front. Plant Sci. 2020, 11, 785. [Google Scholar] [CrossRef]
- Long, H.T.; Zheng, Z.; Zhang, Y.J.; Xing, P.Z.; Wan, X.R.; Zheng, Y.X.; Li, L. An abscisic acid (ABA) homeostasis regulated by its production, catabolism and transport in peanut leaves in response to drought stress. PLoS ONE 2019, 14, e0213963. [Google Scholar] [CrossRef] [Green Version]
- Scarpeci, T.E.; Frea, V.S.; Zanor, M.I.; Valle, E.M. Overexpression of AtERF019 delays plant growth and senescence and improves drought tolerance in Arabidopsis. J. Exp. Bot. 2017, 68, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Joshi, R.; Singh, B.; Bohra, A.; Chinnusamy, V.; Wani, S.; Hossain, M. Salt stress signaling pathways: Specificity and crosstalk. In Managing Salinity Tolerance in Plants: Molecular and Genomic Perspectives; CRC Press: Boca Raton, FL, USA, 2015; pp. 51–78. [Google Scholar] [CrossRef]
- Singh, K.B.; Foley, R.C.; Oate-Sanchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. An apple NAC transcription factor enhances salt stress tolerance by modulating the ethylene response. Physiol. Plant. 2018, 164, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Manna, M.; Thakur, T.; Chirom, O.; Mandlik, R.; Deshmukh, R.; Salvi, P. Transcription factors as key molecular target to strengthen the drought stress tolerance in plants. Physiol. Plant. 2021, 172, 847–868. [Google Scholar] [CrossRef]
- Kabange, R.N.; Imran, Q.M.; Imran, M.; Shahid, M.; Imran, M.; Khan, M.; Lee, S.U.; Hussain, A.; Lee, I.J.; Yun, B.W. Drought-induced AtbZIP62 transcription factor regulates drought stress response in Arabidopsis. Plant Physiol. Biochem. 2020, 156, 384–395. [Google Scholar] [CrossRef]
- Shim, J.S.; Oh, N.; Chung, P.J.; Kim, Y.S.; Yang, D.; Kim, J.K. Overexpression of OsNAC14 improves drought tolerance in rice. Front. Plant Sci. 2018, 9, 310. [Google Scholar] [CrossRef]
- Tiwari, P.; Indoliya, Y.; Chauhan, A.S.; Singhac, P.; Singhac, P.K.; Singhd, P.C.; Srivastava, S.; Pande, V.; Chakrabarty, D. Auxin-salicylic acid cross-talk ameliorates OsMYB-R1 mediated defense towards heavy metal, drought and fungal stress. J. Hazard. Mater. 2020, 399, 122811. [Google Scholar] [CrossRef]
- Yang, S.Q.; Xu, K.; Chen, S.J.; Li, T.F.; Xia, H.; Chen, L.; Liu, H.; Luo, L. A stress-responsive bZIP transcription factor OsbZIP62 improves drought and oxidative tolerance in rice. BMC Plant Biol. 2019, 19, 260. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, Y.; Qiang, L.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef]
- Okamuro, J.K.; Caster, B.; Villarroel, R.; Van-Montagu, M.; Jofuku, K.D. The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 7076–7081. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K.; Seki, M. Regulatory network of gene expression in the drought and cold stress responses. Curr. Opin. Plant Biol. 2003, 6, 410–417. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Leon-Reyes, A.; Vander, E.S.; Van, W.S. Networking by small- molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugano, S.; Sugimoto, T.; Takatsuji, H.; Jianga, C.J. Induction of resistance to Phytophthora sojae in soybean (Glycine max) by salicylic acid and ethylene. Plant Pathol. 2013, 62, 1048–1056. [Google Scholar] [CrossRef]
- Jin, J.H.; Wang, M.; Zhang, H.X.; Khan, A.; Wei, A.M.; Luo, D.X.; Gong, Z.H. Genome-wide identification of the AP2/ERF transcription factor family in pepper (Capsicum annuum L.). Genome 2018, 61, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Faraji, S.; Filiz, E.; Kazemitabar, S.K.; Vannozzi, A.; Palumbo, F.; Barcaccia, G.; Heidari, P. The AP2/ERF gene family in Triticum durum: Genome-wide identification and expression analysis under drought and salinity stresses. Genes 2020, 11, 1464. [Google Scholar] [CrossRef] [PubMed]
- Lv, K.; Li, J.; Zhao, K.; Chen, S.; Nie, J.; Zhang, W.L.; Liu, G.; Wei, H. Overexpression of an AP2/ERF family gene, BpERF13, in birch enhances cold tolerance through upregulating CBF genes and mitigating reactive oxygen species. Plant Sci. 2020, 292, 110375. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, J.; Liu, C.X.; Chen, Z.P.; Yao, Z.C.; Cao, S.Q. Overexpression of ethylene response factor ERF96 gene enhances selenium tolerance in Arabidopsis. Plant Physiol. Biochem. 2020, 149, 294–300. [Google Scholar] [CrossRef]
- Hao, L.; Shi, S.; Guo, H.; Li, M.; Hu, P.; Wei, Y.; Feng, Y. Genome-wide identification and expression profiles of ERF subfamily transcription factors in Zea mays. PeerJ. 2020, 8, e9551. [Google Scholar] [CrossRef]
- Jin, X.; Yin, X.F.; Ndayambaza, B.; Zhang, Z.S.; Min, X.Y.; Lin, X.; Wang, Y.R.; Liu, W. Genome-wide identification and expression profiling of the ERF Gene family in Medicago sativa L. under various abiotic stresses. DNA Cell Biol. 2019, 38, 1056–1068. [Google Scholar] [CrossRef]
- Wan, L.Y.; Wu, Y.S.; Huang, J.Q.; Dai, X.; Lei, Y.; Yan, L.; Jiang, H.; Zhang, J.; Varshney, R.K.; Liao, B. Identification of ERF genes in peanuts and functional analysis of AhERF008 and AhERF019 in abiotic stress response. Funct. Integr. Genomics. 2014, 14, 467–477. [Google Scholar] [CrossRef]
- Rong, W.; Qi, L.; Wang, A.; Ye, X.G.; Du, L.P.; Liang, H.X.; Xin, Z.; Zhang, Z. The ERF transcription factor TaERF3 promotes tolerance to salt and drought stresses in wheat. Plant Biotechnol. J. 2014, 12, 468–479. [Google Scholar] [CrossRef]
- Fujimoto, S.Y.; Ohta, M.; Usui, A.; Shinshi, H.; Ohme-Takagi, M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell. 2000, 12, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Guo, X.; Zhang, M.H.; Wang, X.; Zhao, Y.; Yin, Z.; Zhang, Z.; Wang, Y.; Xiong, H.; Zhang, H.; et al. OsERF71 confers drought tolerance via modulating ABA signaling and proline biosynthesis. Plant Sci. 2018, 270, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.D.; Cheng, Y.X.; Wu, J.J.; Cheng, Q.; Li, W.B.; Fan, S.J.; Jiang, L.; Xu, Z.; Kong, F.; Zhang, D.; et al. Overexpression of GmERF5, a new member of the soybean EAR motif containing ERF transcription factor, enhances resistance to Phytophthora sojae in soybean. J. Exp. Bot. 2015, 66, 2635–2647. [Google Scholar] [CrossRef]
- Lee, D.K.; Yoon, S.; Kim, Y.S.; Kim, J.K. Rice OsERF71-mediated root modification affects shoot drought tolerance. Plant Signal. Behav. 2016, 12, e1268311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, H.Y.; Yu, J.P.; Miao, J.L.; Li, J.J.; Zhang, H.L.; Wang, X.; Liu, P.; Zhao, Y.; Jiang, C.; Yin, Z.; et al. Natural variation in OsLG3 increases drought tolerance in rice by inducing ROS scavenging. Plant Physiol. 2018, 178, 451–467. [Google Scholar] [CrossRef] [Green Version]
- An, J.P.; Zhang, X.W.; Bi, S.Q.; You, C.X.; Wang, X.F.; Hao, Y.J. The ERF transcription factor MdERF38 promotes drought stress-induced anthocyanin biosynthesis in apple. Plant J. 2020, 101, 573–589. [Google Scholar] [CrossRef]
- Ohme-Takagi, M.; Shinshi, H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell. 1995, 7, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Jofuku, K.D.; Omidyar, P.K.; Gee, Z.; Okamuro, J.K. Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc. Natl. Acad. Sci. USA 2005, 102, 3117–3122. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Yu, J.-P.; Chen, F.; Zhao, T.-J.; Fang, X.-H.; Li, Y.-Q.; Sui, S.-F. TINY, a dehydration-responsive element (DRE)-binding protein-like transcription factor connecting the DRE- and ethylene-responsive element-mediated signaling pathways in Arabidopsis. J. Biol. Chem. 2008, 283, 6261–6271. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Zhang, Z.; Zhang, H.; Wang, X.C.; Huang, R. Transcriptional modulation of ethylene response factor protein JERF3 in the oxidative stress response enhances tolerance of tobacco seedlings to salt, drought, and freezing. Plant Physiol. 2008, 148, 1953–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Hu, R.H.; Liu, D.; Liu, X.; Wang, J.; Xiang, X.; Li, Y. The AP2 transcription factor NtERF172 confers drought resistance by modifying NtCAT. Plant Biotechnol. J. 2020, 18, 2444–2455. [Google Scholar] [CrossRef]
- Cheng, M.C.; Hsieh, E.J.; Chen, J.H.; Chen, H.Y.; Lin, T.P. Arabidopsis RGLG2, functioning as a RING E3 ligase, interacts with AtERF53 and negatively regulates the plant drought stress response. Plant Physiol. 2012, 158, 363–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Ekramoddoullah, A.K.M. The family 10 of plant pathogenesis-related proteins their structure, regulation, and function in response to biotic and abiotic stresses. Physiol. Mol. Plant Pathol. 2006, 68, 3–13. [Google Scholar] [CrossRef]
- Ali, M.A.; Abbas, A.; Kreil, D.P.; Bohlmann, H. Overexpression of the transcription factor RAP2.6 leads to enhanced callose deposition in syncytia and enhanced resistance against the beet cyst nematode Heterodera schachtii in Arabidopsis roots. BMC Plant Biol. 2013, 13, 47. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Kim, S.G.; Kang, K.; Kim, J.; Park, S.; Gupta, R.; Kim, Y.H.; Wang, Y.; Kim, S.T. Overexpression of a pathogenesis-related protein 10 enhances biotic and abiotic stress tolerance in rice. Plant Pathol. J. 2016, 32, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, P.; Bhatt, V.; Singh, R.; Das, M.; Sopory, S.; Chikara, J. Pathogenesis-related gene, JcPR-10a from Jatropha curcas exhibit RNase and antifungal activity. Mol. Biotechnol. 2013, 54, 412–425. [Google Scholar] [CrossRef]
- Xu, P.; Jiang, L.; Wu, J.; Li, W.; Fan, S.; Zhang, S. Isolation and characterization of a pathogenesis-related protein 10 gene (GmPR10) with induced expression in soybean (Glycine max) during infection with Phytophthora sojae. Mol. Biol. Rep. 2014, 41, 4899–4909. [Google Scholar] [CrossRef]
- Li, S.; Wang, Z.; Tang, B.F.; Zheng, L.L.; Chen, H.J.; Cui, X.M.; Ge, F.; Liu, D. A pathogenesis-related protein-like gene is involved in the Panax notoginseng defense response to the root rot pathogen. Front. Plant Sci. 2021, 11, 610176. [Google Scholar] [CrossRef]
- Zhang, X.; Menard, R.; Li, Y.; Coruzzi, G.; Heitz, T.; Shen, W.; Berr, A. Arabidopsis SDG8 potentiates the sustainable transcriptional induction of the Pathogenesis-Related genes PR1 and PR2 during plant defense response. Front. Plant Sci. 2020, 11, 277. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Cheng, Q.; Wang, H.Y.; Gao, H.; Fang, X.; Chen, X.; Zhao, M.; Wei, W.; Song, B.; Liu, S.; et al. GmBTB/POZ promotes the ubiquitination and degradation of LHP1 to regulate the response of soybean to Phytophthora sojae. Commun. Biol. 2021, 4, 372. [Google Scholar] [CrossRef]
- Jain, S.; Kumar, D.; Jain, M.; Chaudhary, P.; Deswal, R.; Sarin, N. Ectopic overexpression of a salt stress-induced pathogenesis-related class 10 protein (PR10) gene from peanut (Arachis hypogaea L.) affords broad spectrum abiotic stress tolerance in transgenic tobacco. Plant Cell Tiss. Org. Cult. 2012, 10, 19–31. [Google Scholar] [CrossRef]
- Hashimoto, M.; Kisseleva, L.; Sawa, S.; Furukawa, T.; Komatsu, S.; Koshiba, T. A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses possibly via the jasmonic acid signaling pathway. Plant Cell Physiol. 2004, 45, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, O.R.; Pulla, R.K.; Kim, Y.J.; Baluswamy, R.D.; Khorolragchaa, A.; Yang, D.C. Expression of the ginseng PgPR10-1 in Arabidopsis confers resistance against fungal and bacterial infection. Gene 2012, 506, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, Y.; Liu, G.; Li, H. Cloning and characterization of a Pathogenesis-Related gene (ThPR10) from Tamarix hispida. Acta Biol. Cracov. Bot. 2010, 52, 17–25. [Google Scholar] [CrossRef]
- Li, R.; Liu, C.X.; Zhao, R.R.; Wang, L.; Chen, L.Y.; Yu, W.; Zhang, S.; Sheng, J.; Shen, L. CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant Biol. 2019, 19, 38. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.X.; Zhang, F.C.; Zhang, W.Z.; Song, L.F.; Wu, W.H.; Chen, Y.F. Arabidopsis Di19 functions as a transcription factor and modulates PR1, PR2, and PR5 expression in response to drought stress. Mol. Plant. 2013, 6, 1487–1502. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.-J.; Yin, L.-J.; Liu, Y.; Ma, J.; Zheng, J.-C.; Lan, J.-H.; Fu, J.-D.; Chen, M.; Xu, Z.-S.; Ma, Y.-Z. The ABA-induced soybean ERF transcription factor gene GmERF75 plays a role in enhancing osmotic stress tolerance in Arabidopsis and soybean. BMC Plant Biol. 2019, 19, 506. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.Y.; Chang, X.; Qi, D.Y.; Zhang, S.; Dong, L.; Wang, G.; Fan, S.; Jiang, L.; Cheng, Q.; Chen, X.; et al. A novel soybean ERF transcription factor, GmERF113, increases resistance to Phytophthora sojae infection in soybean. Front. Plant Sci. 2017, 8, 299. [Google Scholar] [CrossRef] [Green Version]
- Ruth, G. Oxidative stress and acclimation mechanisms in plants. Arab. Book 2002, 1, e0036. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S.; Mackay, A. Ion transport in Halophytes. Adv. Bot. Res. 2011, 57, 151–187. [Google Scholar] [CrossRef]
- Jiang, M.Y.; Jing, J.H.; Wang, S.T. Water stress and membrane-lipid peroxidation in plants. Acta. Univ. Agric. Boreali-Occidentalis. 1991, 19, 88–94. [Google Scholar]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell. 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Rustagi, A.; Jain, S.; Kumar, D.; Shekhar, S.; Jain, M.; Bhat, V.; Sarin, N.B. High efficiency transformation of Banana [Musa acuminata L. cv. Matti (AA)] for enhanced tolerance to salt and drought stress through overexpression of a peanut salinity-induced Pathogenesis-Related class 10 protein. Mol. Biotechnol. 2015, 57, 27–35. [Google Scholar] [CrossRef]
- Koberg, M.; Abu-Much, R.; Gedanken, A. Optimization of bio-diesel production from soybean and wastes of cooked oil: Combining dielectric microwave irradiation and a SrO catalyst. Bioresour. Technol. 2011, 102, 1073–1078. [Google Scholar] [CrossRef]
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 2006, 9, 189–195. [Google Scholar] [CrossRef]
- Chen, W.; Yao, Q.; Patil, G.B.; Agarwal, G.; Deshmukh, R.K.; Lin, L.; Wang, B.; Wang, Y.; Prince, S.J.; Song, L.; et al. Identification and comparative analysis of differential gene expression in soybean leaf tissue under drought and flooding stress revealed by RNA-Seq. Front. Plant Sci. 2016, 7, 1044. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.G.; Shi, J.L.; Xu, W.R.; Li, H.E.; He, M.Y.; Xu, Y.; Xu, T.; Yang, Y.; Cao, J.; Wang, Y. Three ERF transcription factors from Chinese wild grapevine Vitis pseudoreticulata participate in different biotic and abiotic stress-responsive pathways. J. Plant Physiol. 2013, 170, 923–933. [Google Scholar] [CrossRef]
- Xing, L.P.; Di, Z.C.; Yang, W.W.; Liu, J.Q.; Li, M.N.; Wang, X.J.; Cui, C.; Wang, X.; Wang, X.; Zhang, R.; et al. Overexpression of ERF1-V from Haynaldia villosa can enhance the resistance of wheat to powdery mildew and increase the tolerance to salt and drought stresses. Front. Plant Sci. 2017, 8, 1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.J.; Tian, Y.S.; Xu, J.; Fu, X.Y.; Gao, J.J.; Wang, B.; Han, H.; Wang, L.; Peng, R.; Yao, Q. A tomato ERF transcription factor, SlERF84, confers enhanced tolerance to drought and salt stress but negatively regulates immunity against Pseudomonas syringae pv. tomato DC3000. Plant Physiol. Biochem. 2018, 132, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Y.; Chen, M.; Li, L.; Xu, Z.; Chen, X.; Guo, J.; Ma, Y. Overexpression of the soybean GmERF3 gene, an AP2/ERF type transcription factor for increased tolerances to salt, drought, and diseases in transgenic tobacco. J. Exp. Bot. 2009, 60, 3781–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Chen, M.; Chen, X.; Xu, Z.; Li, L.; Guo, J.; Ma, Y. Isolation and characterization of a novel EAR-motif-containing gene GmERF4 from soybean (Glycine max L.). Mol. Biol. Rep. 2010, 37, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Li, J.-W.; Li, X.-W.; Lei, T.-T.; Yan, F.; Zhao, Y.; Li, Y.-J.; Su, L.-T.; Wang, Y.; Wang, Q.-Y. Isolation and characterization of a novel transcriptional repressor GmERF6 from soybean. J. Ind. Eng. Chem. 2012, 57, 624–629. [Google Scholar] [CrossRef]
- Peng, Q.; Su, Y.C.; Ling, H.; Ahmad, W.; Gao, S.W.; Guo, J.L. A sugarcane pathogenesis-related protein, ScPR10, plays a positive role in defense responses under Sporisorium scitamineum, SrMV, SA, and MeJA stresses. Plant Cell Rep. 2017, 36, 1427–1440. [Google Scholar] [CrossRef]
- Besbes, F.; Habegger, R.; Schwab, W. Induction of PR-10 genes and metabolites in strawberry plants in response to Verticillium dahliae infection. BMC Plant Biol. 2019, 19, 128. [Google Scholar] [CrossRef] [Green Version]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis related protein-1 (PR-1) genes in tomato (Solanum lycopersicum L.): Bioinformatics analyses and expression profiles in response to drought stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef]
- Zribi, I.; Ghorbel, M.; Brini, F. Pathogenesis related proteins (PRs): From cellular mechanisms to plant defense. Curr. Protein Pept. Sci. 2021, 22, 396–412. [Google Scholar] [CrossRef]
- Takeuchi, K.; Hasegawa, H.; Gyohda, A.; Komatsu, S.; Okamoto, T.; Okada, K.; Terakawa, T.; Koshiba, T. Overexpression of RSOsPR10, a root-specific rice PR10 gene, confers tolerance against drought stress in rice and drought and salt stresses in bentgrass. Plant Cell Tiss. Org. 2016, 127, 35–46. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Ni, Z.Y.; Chen, Q.J.; Guo, Z.J.; Gao, W.W.; Su, X.J.; Qu, Y. Proteomic responses of drought-tolerant and drought-sensitive cotton varieties to drought stress. Mol. Genet. Genomics 2016, 291, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Han, X.J.; He, X.L.; Qiu, W.M.; Lu, Z.C.; Zhang, Y.X.; Chen, S.S.; Liu, M.; Qiao, G.; Zhuo, R. Pathogenesis-related protein PR10 from Salix matsudana Koidz exhibits resistance to salt stress in transgenic Arabidopsis thaliana. Environ. Exp. Bot. 2017, 141, 74–82. [Google Scholar] [CrossRef]
- Eramalingam, A.; Kudapa, H.; Pazhamala, L.T.; Egarg, V.; Varshney, R.K. Gene expression and yeast two-hybrid studies of transcription factors mediating drought stress response in root tissues of chickpea (Cicer arietinum L.). Front. Plant Sci. 2015, 6, 1117. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.; Geisler, M.; Su, J.; Kolukisaoglu, H.; Azevedo, L.; Plaza, S.; Curtis, M.D.; Richter, A.; Weder, B.; Schulz, B.; et al. Disruption of AtMRP4, a guard cell plasma membrane ABCC-type ABC transporter, leads to deregulation of stomatal opening and increased drought susceptibility. Plant J. 2004, 39, 219–236. [Google Scholar] [CrossRef]
- Ramalingam, J.; Pathan, M.; Feril, O.; Miftahudin; Ross, K.; Ma, X.; Mahmoud, A.; Layton, J.; Rodriguez-Milla, M.; Chikmawati, T.; et al. Structural and functional analyses of the wheat genomes based on expressed sequence tags (ESTs) related to abiotic stresses. Genome 2006, 49, 1324–1340. [Google Scholar] [CrossRef]
- Hou, F.; Du, T.; Qin, Z.; Xu, T.; Li, A.; Dong, S.; Ma, D.; Li, Z.; Wang, Q.; Zhang, L. Genome-wide in silico identification and expression analysis of beta-galactosidase family members in sweetpotato [Ipomoea batatas (L.) Lam]. BMC Genomics 2021, 22, 140. [Google Scholar] [CrossRef]
- Wang, H.; Dong, Q.; Duan, D.; Zhao, S.; Li, M.; van Nocker, M.; Ma, F.; Mao, K. Comprehensive genomic analysis of the TYROSINE AMINOTRANSFERASE (TAT) genes in apple (Malus domestica) allows the identification of MdTAT2 conferring tolerance to drought and osmotic stresses in plants. Plant Physiol. Biochem. 2018, 133, 81–91. [Google Scholar] [CrossRef]
- Li, Y.; Wang, B.; Dong, R.; Hou, B. AtUGT76C2, an Arabidopsis cytokinin glycosyltransferase is involved in drought stress adaptation. Plant Sci. 2015, 236, 157–167. [Google Scholar] [CrossRef]
- Wu, X.; Qiao, Z.; Liu, H.; Acharya, B.; Li, C.; Zhang, W. CML20, an Arabidopsis Calmodulin-like protein, negatively regulates guard cell ABA signaling and drought stress tolerance. Front. Plant Sci. 2017, 8, 824. [Google Scholar] [CrossRef] [Green Version]
- Seong, S.; Shim, S.; Bang, W.; Kim, J. Overexpression of OsC3H10, a CCCH-Zinc Finger, improves drought tolerance in rice by regulating stress-related genes. Plants. 2020, 9, 1298. [Google Scholar] [CrossRef]
- Zhai, Y.; Wang, Y.; Li, Y.J.; Lei, T.T.; Yan, F.; Su, L.T.; Li, X.; Zhao, Y.; Sun, X.; Li, J.; et al. Isolation and molecular characterization of GmERF7, a soybean ethylene-response factor that increases salt stress tolerance in tobacco. Gene 2013, 513, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N.; Gill, S.; Trivedi, D. Abscisic Acid (ABA): Biosynthesis, regulation, and role in abiotic stress tolerance. Abiotic Stress Response Plants 2016, 15, 311–322. [Google Scholar] [CrossRef]
- Kondo, S.; Sugaya, S.; Sugawa, S.; Ninomiya, M.; Kittikorn, M.; Okawa, K.; Ohara, H.; Ueno, K.; Todoroki, Y.; Mizutani, M.; et al. Dehydration tolerance in apple seedlings is affected by an inhibitor of ABA 8′-hydroxylase CYP707A. J. Plant Physiol. 2012, 169, 234–241. [Google Scholar] [CrossRef]
- Shang, X.; Yu, Y.; Zhu, L.; Liu, H.; Chai, Q.; Guo, W. A cotton NAC transcription factor GhirNAC2 plays positive roles in drought tolerance via regulating ABA biosynthesis. Plant Sci. 2020, 296, 110498. [Google Scholar] [CrossRef]
- Qu, L.; Sun, M.; Li, X.; He, R.; Zhong, M.; Luo, D.; Liu, X.; Zhao, X. The Arabidopsis F-box protein FOF2 regulates ABA-mediated seed germination and drought tolerance. Plant Sci. 2020, 301, 110643. [Google Scholar] [CrossRef]
- Takeuchi, J.; Okamoto, M.; Mega, R.; Kanno, Y.; Ohnishi, T.; Seo, M.; Todoroki, Y. Abscinazole-E3M, a practical inhibitor of abscisic acid 8′-hydroxylase for improving drought tolerance. Sci. Rep. 2016, 6, 37060. [Google Scholar] [CrossRef]
- Belmiro, V.; Alicia, M.; Montserrat, C.; Montserrat, P.; Victoria, L. ZmSnRK2.8 responds to ABA through the SnRK2-PP2C complex. Maydica 2012, 57, 11–18. [Google Scholar]
- Yang, Q.; Liu, K.; Niu, X.; Wang, Q.; Wan, Y.Q.; Yang, F.Y.; Li, G.; Wang, Y.; Wang, R. Genome-wide identification of PP2C genes and their expression profiling in response to drought and cold stresses in Medicago truncatula. Sci. Rep. 2018, 8, 12841. [Google Scholar] [CrossRef] [Green Version]
- Belda-Palazon, B.; Julian, J.; Coego, A.; Wu, Q.; Zhang, X.; Batistic, O.; AlQuraishi, S.A.; Kudla, J.; An, C.; Rodriguez, P.L. ABA inhibits myristoylation and induces shuttling of the RGLG1 E3 ligase to promote nuclear degradation of PP2CA. Plant J. 2019, 98, 813–825. [Google Scholar] [CrossRef]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; MacLean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell. 2004, 16, 3460–3479. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Pajerowska-Mukhtar, K.; Culler, A.H.; Dong, X.N. Salicylic acid inhibits pathogen growth in plants through repression of the auxin signaling pathway. Curr. Biol. 2007, 17, 1784–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, F.Y.; Yoshioka, K.; Desveaux, D. The roles of ABA in plant-pathogen interactions. J. Plant Res. 2011, 124, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.X.; Hu, Y.J.; Tang, X.K.; Zhou, P.L.; Deng, X.B.; Wang, H.H.; Guo, Z. Constitutive expression of rice WRKY30 gene increases the endogenous jasmonic acid accumulation, PR gene expression and resistance to fungal pathogens in rice. Planta 2012, 236, 1485–1498. [Google Scholar] [CrossRef]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef]
- Ford, R.; Tan, D.; Vaghefi, N.; Mustafa, B. Abscisic Acid Activates Pathogenesis-Related Defense Gene Signaling in Lentils; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 243–270. [Google Scholar]
- Chatterjee, M.; Chakraborty, J.; Das, S. Abscisic acid-responsive 18 (CaABR18) protein from chickpea inhibits the growth of the wilt-causing Fusarium oxysporum f. sp. ciceri race1. Plant Mol. Biol. Rep. 2019, 37, 170–185. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Ma, Q.B.; Xia, Z.L.; Cai, Z.D.; Li, L.; Cheng, Y.B.; Liu, J.; Nian, H. GmWRKY16 enhances drought and salt tolerance through an ABA-mediated pathway in Arabidopsis thaliana. Front. Plant Sci. 2019, 9, 1979. [Google Scholar] [CrossRef] [Green Version]
- Kerschen, A.; Napoli, C.A.; Jorgensen, R.A.; Muller, A.E. Effectiveness of RNA interference in transgenic plants. FEBS Lett. 2004, 566, 223–228. [Google Scholar] [CrossRef]
- Paz, M.M.; Shou, H.; Guo, Z.; Zhang, Z.; Banerjee, A.K.; Wang, K. Assessment of conditions affecting Agrobacterium-mediated soybean transformation using the cotyledonary node explants. Euphytica 2004, 136, 167–179. [Google Scholar] [CrossRef]
- Wang, K.; Bu, T.T.; Cheng, Q.; Dong, L.D.; Su, T.; Chen, Z.M.; Kong, F.; Gong, Z.; Liu, B.; Li, M. Two homologous LHY pairs negatively control soybean drought tolerance by repressing the abscisic acid responses. New Phytol. 2021, 229, 2660–2675. [Google Scholar] [CrossRef]
- Saleh, A.; Alvarez-Venegas, R.; Avramova, Z. An efficient chromatin immunoprecipitation (ChIP) protocol for studying histone modifications in Arabidopsis plants. Nat. Protoc. 2008, 3, 1018–1025. [Google Scholar] [CrossRef]
- Kereszt, A.; Li, D.; Indrasumunar, A.; Nguyen, C.D.; Nontachaiyapoom, S.; Kinkema, M.; Gresshoff, P.M. Agrobacterium rhizogenes-mediated transformation of soybean to study root biology. Nat. Protoc. 2007, 2, 948–952. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshla, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.; Luo, Z.L.; Li, Y.P.; Ren, G.X.; Liu, C.S.; Ma, X.J. Effect of abscisic acid on accumulation of five active components in root of Glycyrrhiza uralensis. Molecules 2017, 22, 1982. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality. Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
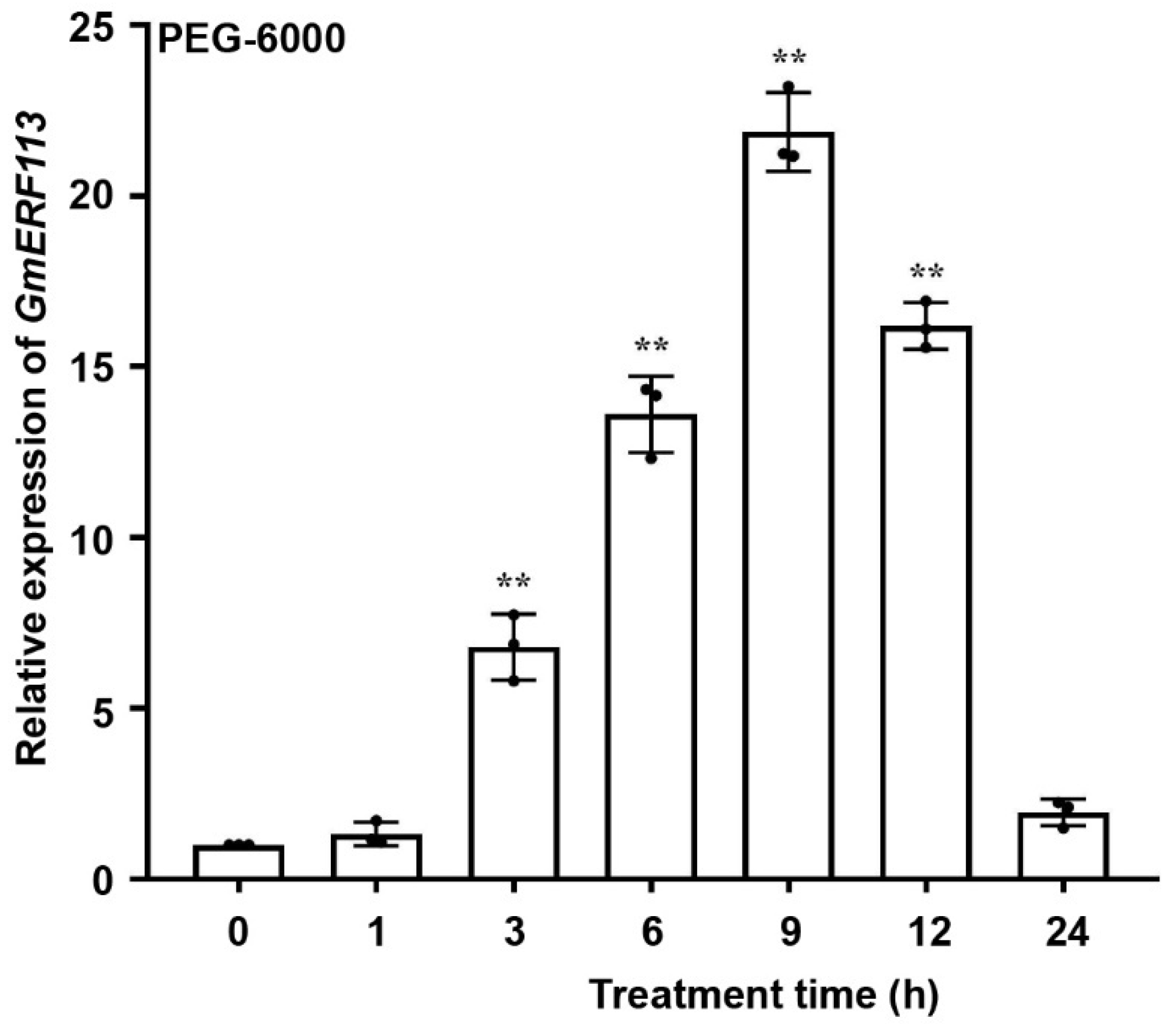
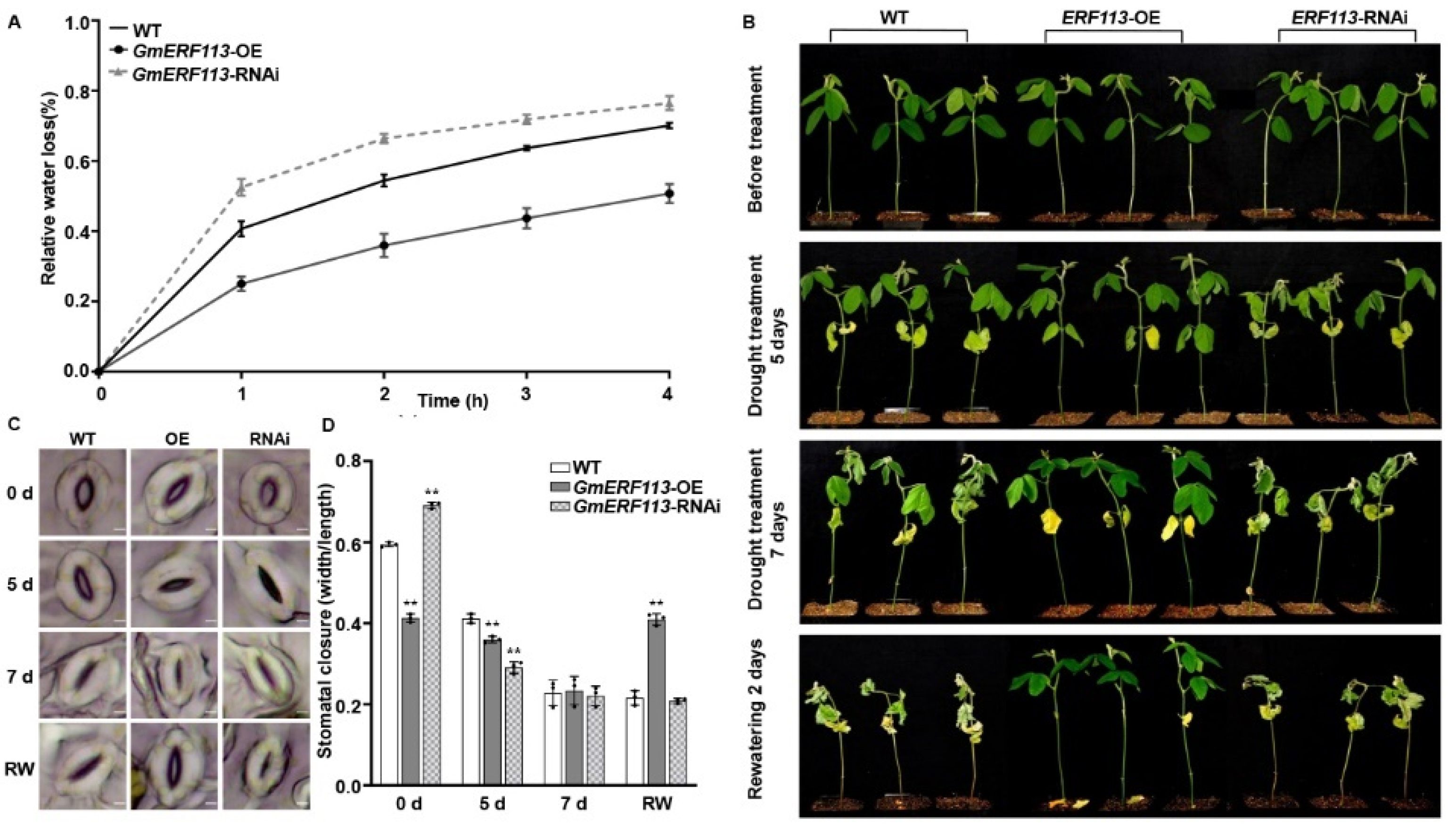
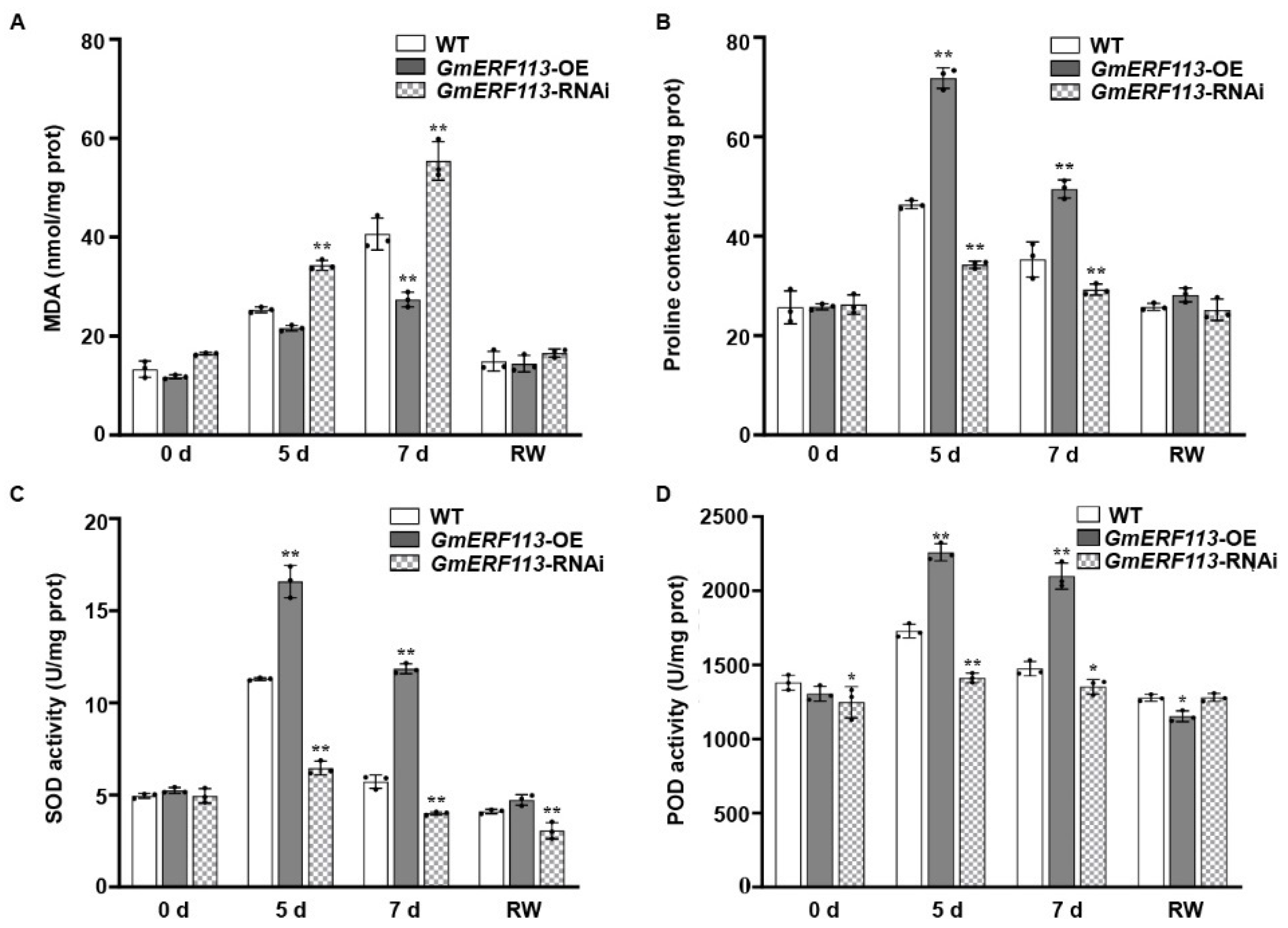
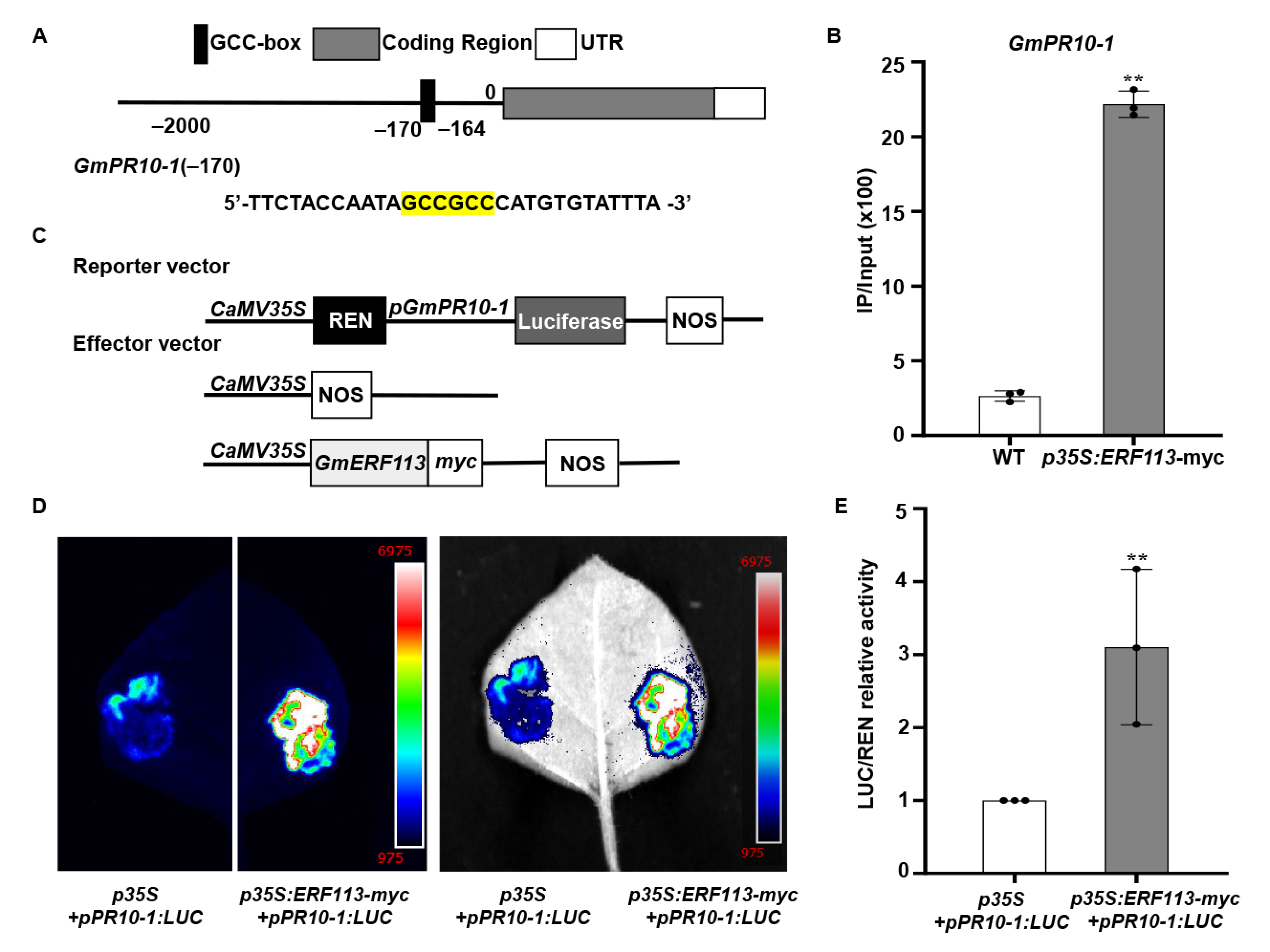
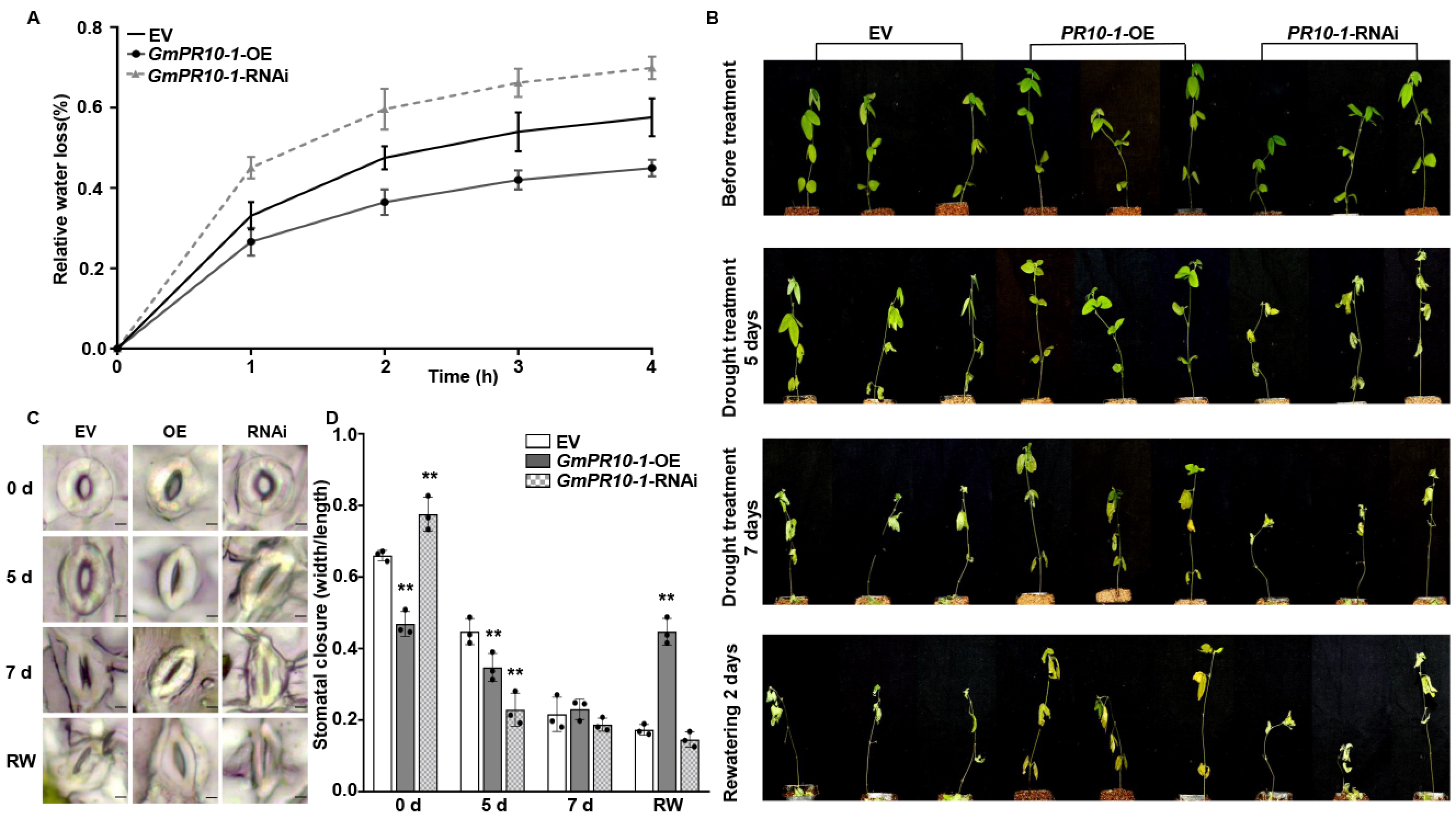
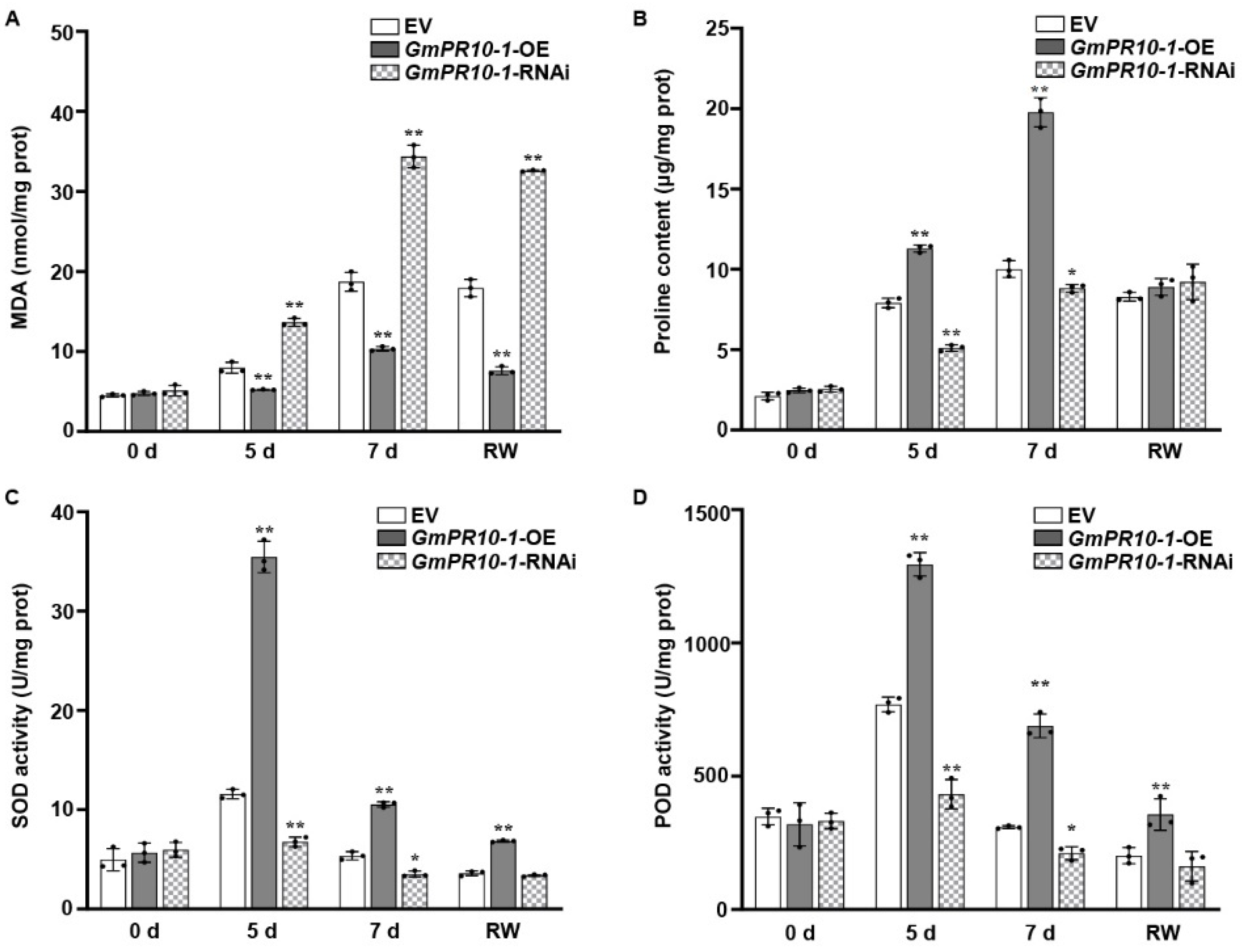
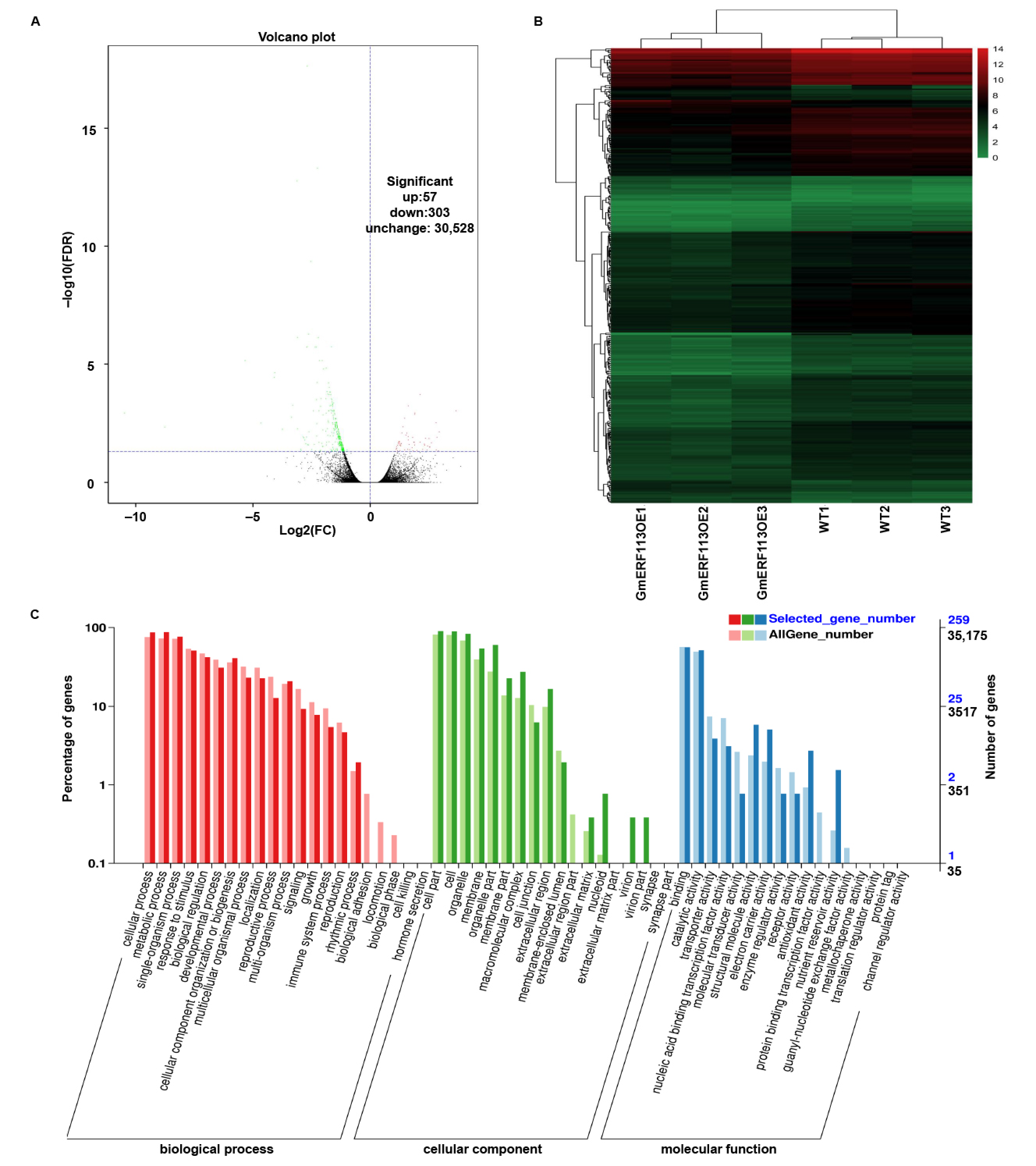
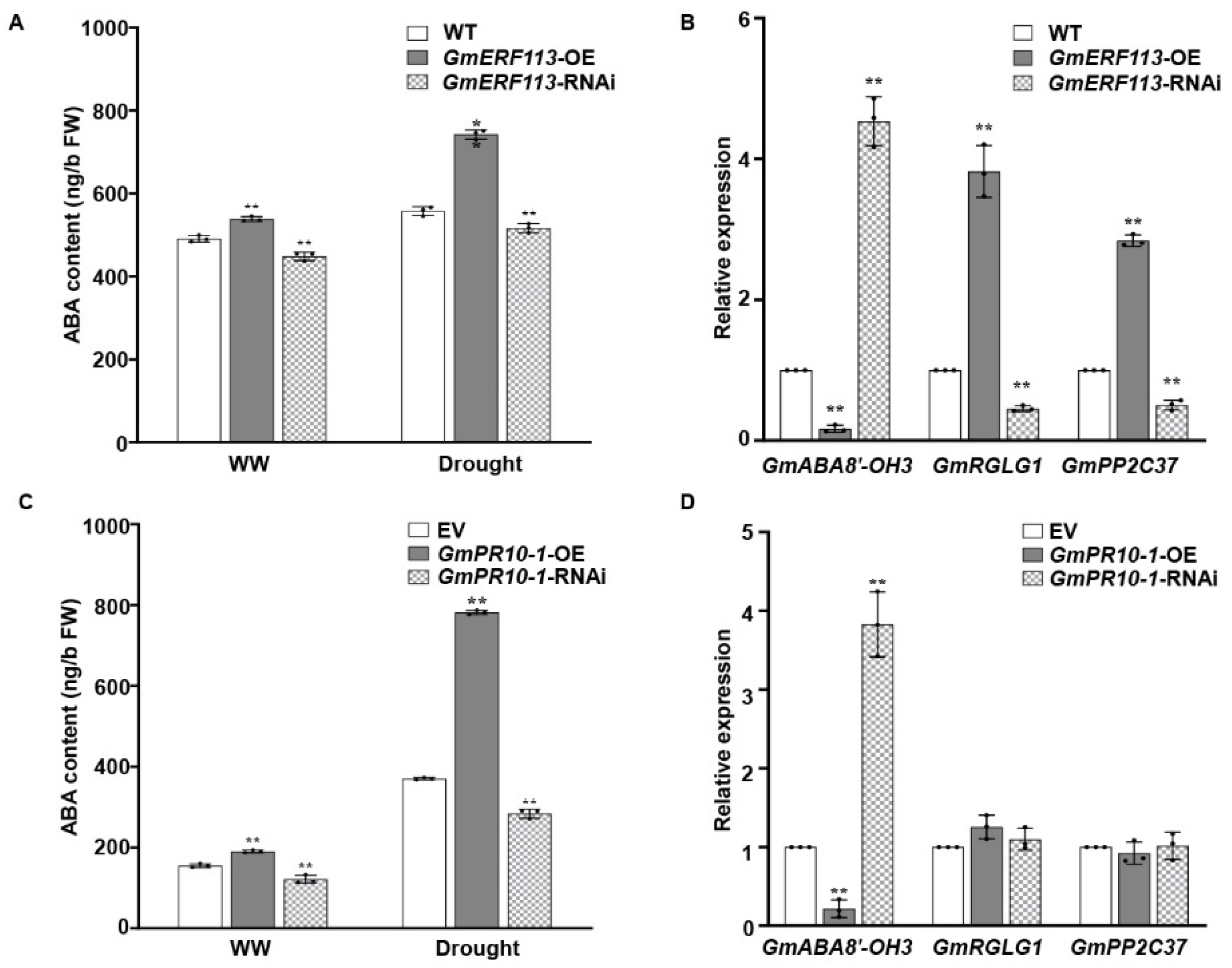
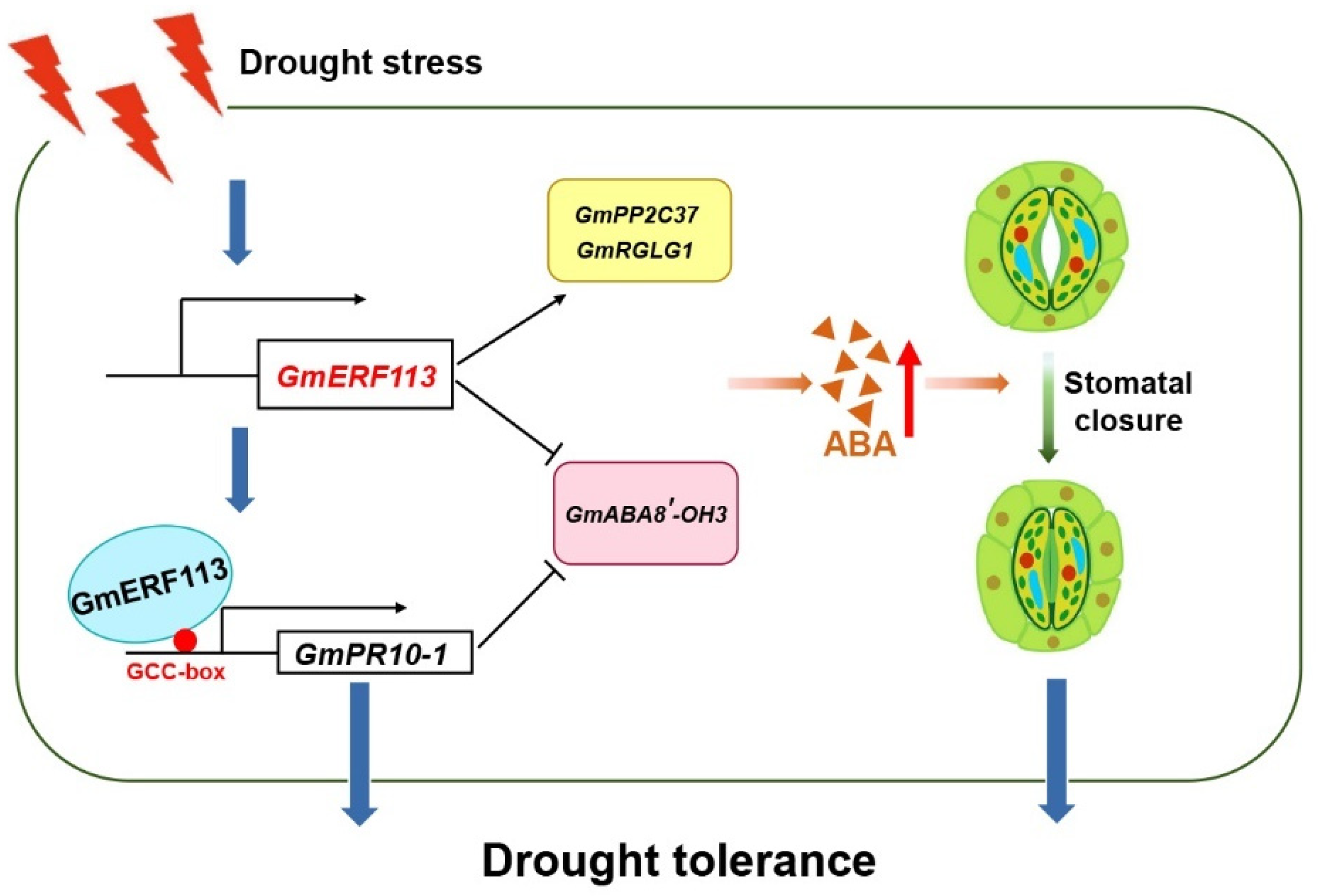
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).