
Carbamate Pesticides: Shedding Light on Their Impact on the Male Reproductive System

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. The Role of Acetylcholine in Male Reproductive Function
3. Carbamates as Endocrine-Disrupting Chemicals (EDCs)
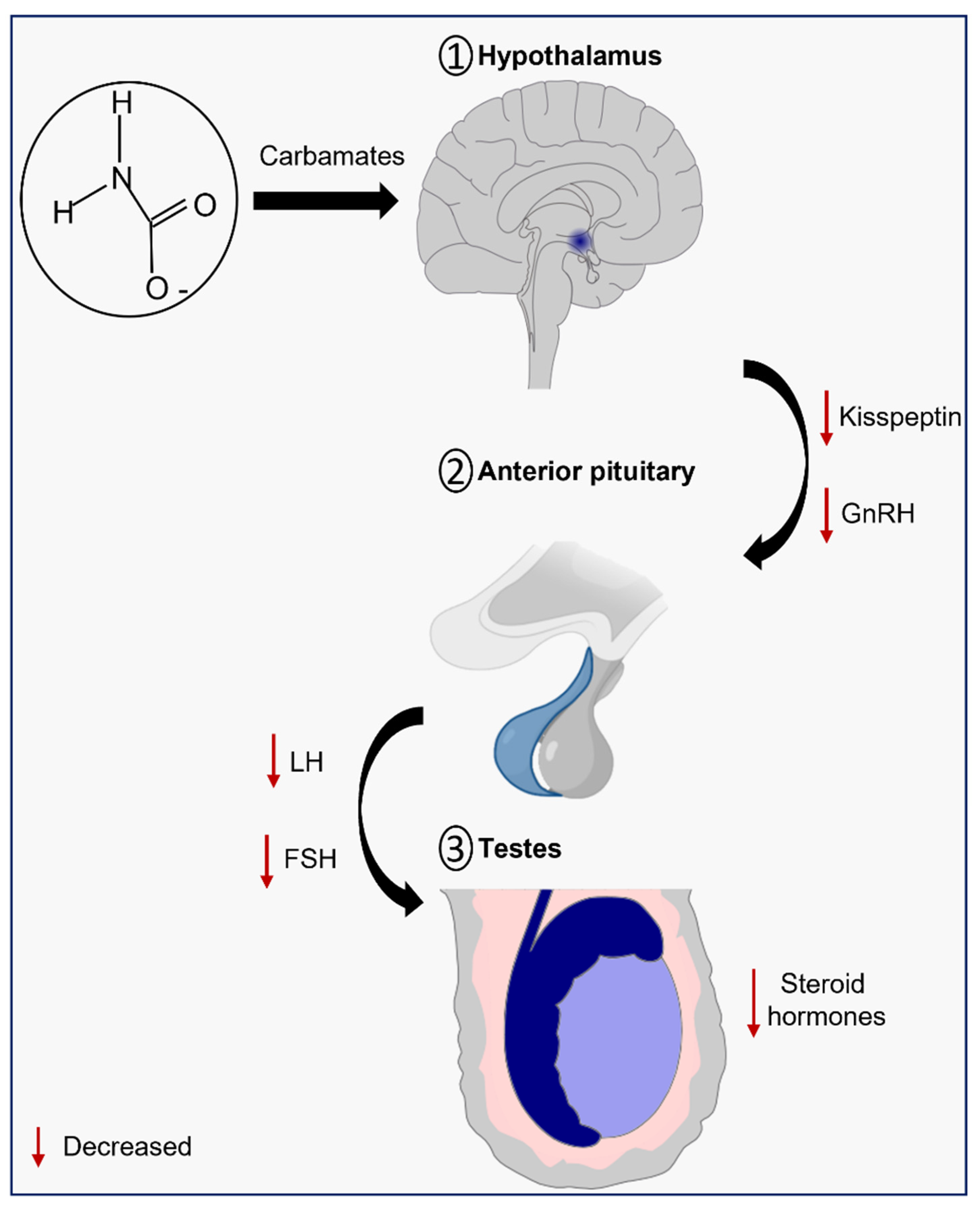
4. Carbamates and Kisspeptin
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- King, A.M.; Aaron, C.K. Organophosphate and carbamate poisoning. Emerg. Med. Clin. N. Am. 2015, 33, 133–151. [Google Scholar] [CrossRef] [PubMed]
- Martin-Reina, J.; Duarte, J.A.; Cerrillos, L.; Bautista, J.D.; Moreno, I. Insecticide Reproductive Toxicity Profile: Organophosphate, Carbamate and Pyrethroids. J. Toxins 2017, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- WHO. Carbamate Pesticides: A General Introduction; World Health Organization: Geneva, Switzerland, 1986. [Google Scholar]
- Gupta, R.C.; Mukherjee, I.R.; Doss, R.B.; Malik, J.K.; Milatovic, D. Organophosphates and Carbamates. In Reproductive and Developmental Toxicology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 609–631. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Brindisi, M. Organic carbamates in drug design and medicinal chemistry. J. Med. Chem. 2015, 58, 2895–2940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzal, S.; Shah, S.S.; Iqbal, M.F.; Noureen, A. Review on effect of carbamate pesticide on male reproductive system of mammals. Int. J. Entomol. Res. 2018, 3, 31–33. [Google Scholar]
- Liu, J.; Zhang, P.; Zhao, Y.; Zhang, H. Low dose carbendazim disrupts mouse spermatogenesis might Be through estrogen receptor related histone and DNA methylation. Ecotoxicol. Environ. Saf. 2019, 176, 242–249. [Google Scholar] [CrossRef]
- Li, H.; Zhang, P.; Zhao, Y.; Zhang, H. Low doses of carbendazim and chlorothalonil synergized to impair mouse spermatogenesis through epigenetic pathways. Ecotoxicol. Environ. Saf. 2020, 188, 109908. [Google Scholar] [CrossRef]
- Combarnous, Y. Endocrine Disruptor Compounds (EDCs) and agriculture: The case of pesticides. Comptes Rendus Biol. 2017, 340, 406–409. [Google Scholar] [CrossRef]
- Mor, I.; Soreq, H. Cholinergic Toxicity and the Male Reproductive System. Reprod. Dev. Toxicol. 2011, 66, 863–870. [Google Scholar] [CrossRef]
- Dajas-Bailador, F.; Wonnacott, S. Nicotinic acetylcholine receptors and the regulation of neuronal signalling. Trends Pharmacol. Sci. 2004, 25, 317–324. [Google Scholar] [CrossRef]
- Harvey, R.D. Muscarinic receptor agonists and antagonists: Effects on cardiovascular function. Muscarinic Recept. 2012, 208, 299–316. [Google Scholar]
- Abrams, P.; Andersson, K.E.; Buccafusco, J.J.; Chapple, C.; De Groat, W.C.; Fryer, A.D.; Kay, G.; Laties, A.; Nathanson, N.M.; Pasricha, P.J. Muscarinic receptors: Their distribution and function in body systems, and the implications for treating overactive bladder. Br. J. Pharmacol. 2006, 148, 565–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, M.O.; Abreu, M.L.; Porto, C.S.; Avellar, M.C.W. Characterization of muscarinic acetylcholine receptor in rat Sertoli cells. Endocrinology 2001, 142, 4701–4710. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, C.; Ma, X.; Yan, X.; Xiong, B.; Shen, W.; Yin, S.; Zhang, H.; Sun, Q.; Zhao, Y. Muscarinic acetylcholine receptor M5 is involved in spermatogenesis through the modification of cell-cell junctions. Reproduction 2021, 162, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Porto, C.; Lucas, T.; Siu, E.; Royer, C.; Trindade, E.; Nader, H.; Lazari, M. Muscarinic Acetylcholine Receptors: Relevance to Infertility and Male Contraception. Immunol. Endocr. Metab. Agents Med. Chem. 2008, 8, 42–50. [Google Scholar] [CrossRef]
- Avellar, M.C.; Siu, E.R.; Yasuhara, F.; Marostica, E.; Porto, C.S. Muscarinic acetylcholine receptor subtypes in the male reproductive tract: Expression and function in rat efferent ductules and epididymis. J. Mol. Neurosci. 2010, 40, 127–134. [Google Scholar] [CrossRef]
- Avellar, M.C.; Lazari, M.F.; Porto, C.S. Expression and function of G-protein-coupled receptors in the male reproductive tract. Acad. Bras. Cienc. 2009, 81, 321–344. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.J.; Ishikawa, Y.; Yagi, H.; Iguchi, T.; Oka, Y.; Kuroda, K.; Iwata, K.; Kiyonari, H.; Matsuda, S.; Matsuzaki, H.; et al. PIP3-Phldb2 is crucial for LTP regulating synaptic NMDA and AMPA receptor density and PSD95 turnover. Sci. Rep. 2019, 9, 4305. [Google Scholar] [CrossRef]
- Albuquerque, E.X.; Pereira, E.F.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [Green Version]
- Zoli, M.; Pucci, S.; Vilella, A.; Gotti, C. Neuronal and Extraneuronal Nicotinic Acetylcholine Receptors. Curr. Neuropharmacol. 2018, 16, 338–349. [Google Scholar] [CrossRef]
- Bray, C.; Son, J.H.; Kumar, P.; Meizel, S. Mice deficient in CHRNA7, a subunit of the nicotinic acetylcholine receptor, produce sperm with impaired motility. Biol. Reprod. 2005, 73, 807–814. [Google Scholar] [CrossRef]
- Arican, E.Y.; Gokceoglu Kayali, D.; Ulus Karaca, B.; Boran, T.; Ozturk, N.; Okyar, A.; Ercan, F.; Ozhan, G. Reproductive effects of subchronic exposure to acetamiprid in male rats. Sci. Rep. 2020, 10, 8985. [Google Scholar] [CrossRef] [PubMed]
- Wessler, I.K.; Kirkpatrick, C.J. Non-neuronal acetylcholine involved in reproduction in mammals and honeybees. J. Neurochem. 2017, 142 (Suppl. S2), 144–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, M.R.; Rama Sastry, B.V.; Stavinoha, W.B. Identification of acetylcholine and propionylcholine in bull spermatozoa by integrate pyrolysis, gas chromatography and mass spectrometry. Biochim. Biophys. Acta (BBA)—Gen. Subj. 1977, 500, 440–444. [Google Scholar] [CrossRef]
- Saiko, A. Physiological importance of acetylcholine in sperm cytoplasm. Fiziol. Zhurnal 1969, 15, 537–542. [Google Scholar]
- Schirmer, S.U.; Eckhardt, I.; Lau, H.; Klein, J.; DeGraaf, Y.C.; Lips, K.S.; Pineau, C.; Gibbins, I.L.; Kummer, W.; Meinhardt, A.; et al. The cholinergic system in rat testis is of non-neuronal origin. Reproduction 2011, 142, 157–166. [Google Scholar] [CrossRef]
- Ramirez-Reveco, A.; Villarroel-Espindola, F.; Rodriguez-Gil, J.E.; Concha, I.I. Neuronal signaling repertoire in the mammalian sperm functionality. Biol. Reprod. 2017, 96, 505–524. [Google Scholar] [CrossRef] [Green Version]
- Sliwa, L. Chemotaction of mouse spermatozoa induced by certain hormones. Arch. Androl. 1995, 35, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Sastry, B.V.; Janson, V.E.; Chaturvedi, A.K. Inhibition of human sperm motility by inhibitors of choline acetyltransferase. J. Pharmacol. Exp. Ther. 1981, 216, 378–384. [Google Scholar]
- Sato, T.; Ban, Y.; Uchida, M.; Gondo, E.; Yamamoto, M.; Sekiguchi, Y.; Sakaue, A.; Kemi, M.; Nakatsuka, T. Atropine-induced inhibition of sperm and semen transport impairs fertility in male rats. J. Toxicol. Sci. 2005, 30, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Ban, Y.; Sato, T.; Nakatsuka, T.; Kemi, M.; Samura, K.; Matsumoto, H.; Cukierski, M.; Zwieten, M. Impairment of male fertility induced by muscarinic receptor antagonists in rats. Reprod. Toxicol. 2002, 16, 757–765. [Google Scholar] [CrossRef]
- da Silva Junior, E.D.; de Souza, B.P.; Rodrigues, J.Q.; Caricati-Neto, A.; Jurkiewicz, A.; Jurkiewicz, N.H. Functional characterization of acetylcholine receptors and calcium signaling in rat testicular capsule contraction. Eur. J. Pharmacol. 2013, 714, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Ceron, S.; Vargas-Lopez, H.; Perez-Albacete, M.; Tovar-Zapata, I.; Martinez-Hernandez, P.; Rodriguez-Lopez, J.N.; Cabezas-Herrera, J. Analysis of cholinesterases in human prostate and sperm: Implications in cancer and fertility. Chem. Biol. Interact. 2010, 187, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.K.; Ravichandran, S.; Sukesh, A.; Kadalmani, B.; Kandasamy, M. Single injection of very mild dose botulinum toxin in the vastus lateralis improves testicular spermatogenesis and sperm motility in ageing experimental mice. Lab. Anim. Res. 2022, 38, 7. [Google Scholar] [CrossRef]
- Dressler, D.; Saberi, F.A. Botulinum toxin: Mechanisms of action. Eur. Neurol. 2005, 53, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babakhanzadeh, E.; Nazari, M.; Ghasemifar, S.; Khodadadian, A. Some of the Factors Involved in Male Infertility: A Prospective Review. Int. J. Gen. Med. 2020, 13, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.M.; Yang, W.X. Molecular regulation of hypothalamus-pituitary-gonads axis in males. Gene 2014, 551, 15–25. [Google Scholar] [CrossRef]
- Kaprara, A.; Huhtaniemi, I.T. The hypothalamus-pituitary-gonad axis: Tales of mice and men. Metabolism 2018, 86, 3–17. [Google Scholar] [CrossRef]
- Aghazadeh, Y.; Zirkin, B.R.; Papadopoulos, V. Pharmacological regulation of the cholesterol transport machinery in steroidogenic cells of the testis. Vitam. Horm. 2015, 98, 189–227. [Google Scholar]
- Carrageta, D.F.; Guerra-Carvalho, B.; Spadella, M.A.; Yeste, M.; Oliveira, P.F.; Alves, M.G. Animal models of male reproductive ageing to study testosterone production and spermatogenesis. Rev. Endocr. Metab. Disord. 2022, 1–20. [Google Scholar] [CrossRef]
- Tilbrook, A.; Clarke, I.J. Negative feedback regulation of the secretion and actions of gonadotropin-releasing hormone in males. Biol. Reprod. 2001, 64, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; Ismail, R.S.; Zaki, H.F.; Arafa, H.M.M.; El-Khatib, A.S.N. L-carnitine extenuates endocrine disruption, inflammatory burst and oxidative stress in carbendazim-challenged male rats via upregulation of testicular StAR and FABP9, and downregulation of P38-MAPK pathways. Toxicology 2021, 457, 152808. [Google Scholar] [CrossRef] [PubMed]
- Elsharkawy, E.E.; El-Nasser, M.A.; Bakheet, A.A. Mancozeb impaired male fertility in rabbits with trials of glutathione detoxification. Regul. Toxicol. Pharm. 2019, 105, 86–98. [Google Scholar] [CrossRef]
- Mohanty, B.; Pandey, S.P.; Tsutsui, K. Thyroid disrupting pesticides impair the hypothalamic-pituitary-testicular axis of a wildlife bird, Amandava amandava. Reprod. Toxicol. 2017, 71, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Qiu, L.; Hu, G.; Fan, L.; Song, C.; Zheng, Y.; Wu, W.; Qu, J.; Li, D.; Chen, J.; et al. Effects of methomyl on steroidogenic gene transcription of the hypothalamic-pituitary-gonad-liver axis in male tilapia. Chemosphere 2016, 165, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhou, S.; Chen, Y.; Chen, X.; Liu, J.; Ge, F.; Lian, Q.; Chen, X.; Ge, R.S. Ziram Delays Pubertal Development of Rat Leydig Cells. Toxicol. Sci. 2017, 160, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Abbara, A.; Clarke, S.A.; Dhillo, W.S. Clinical Potential of Kisspeptin in Reproductive Health. Trends Mol. Med. 2021, 27, 807–823. [Google Scholar] [CrossRef]
- Leon, S.; Talbi, R.; McCarthy, E.A.; Ferrari, K.; Fergani, C.; Naule, L.; Choi, J.H.; Carroll, R.S.; Kaiser, U.B.; Aylwin, C.F.; et al. Sex-specific pubertal and metabolic regulation of Kiss1 neurons via Nhlh2. eLife 2021, 10, 69765. [Google Scholar] [CrossRef]
- Padda, J.; Khalid, K.; Moosa, A.; Syam, M.; Kakani, V.; Imdad, U.; Ismail, D.; Cooper, A.C.; Jean-Charles, G. Role of Kisspeptin on Hypothalamic-Pituitary-Gonadal Pathology and Its Effect on Reproduction. Cureus 2021, 13, e17600. [Google Scholar] [CrossRef]
- Salehi, S.; Adeshina, I.; Chen, H.; Zirkin, B.R.; Hussain, M.A.; Wondisford, F.; Wolfe, A.; Radovick, S. Developmental and endocrine regulation of kisspeptin expression in mouse Leydig cells. Endocrinology 2015, 156, 1514–1522. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Li, Z.; Jiang, W.; Ling, Y.; Kuang, H. Reproductive functions of Kisspeptin/KISS1R Systems in the Periphery. Reprod. Biol. Endocrinol. 2019, 17, 65. [Google Scholar] [CrossRef]
- Novaira, H.J.; Sonko, M.L.; Hoffman, G.; Koo, Y.; Ko, C.; Wolfe, A.; Radovick, S. Disrupted kisspeptin signaling in GnRH neurons leads to hypogonadotrophic hypogonadism. Mol. Endocrinol. 2014, 28, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meccariello, R.; Fasano, S.; Pierantoni, R. Kisspeptins, new local modulators of male reproduction: A comparative overview. Gen. Comp. Endocrinol. 2020, 299, 113618. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, C.M.; Montagna, E.; de Oliveira, R.; Christofolini, D.M.; Barbosa, C.P.; Crandall, K.A.; Bianco, B. Kisspeptin/GPR54 System: What Do We Know About Its Role in Human Reproduction? Cell Physiol. Biochem. 2018, 49, 1259–1276. [Google Scholar] [CrossRef]
- Hsu, M.C.; Wang, J.Y.; Lee, Y.J.; Jong, D.S.; Tsui, K.H.; Chiu, C.H. Kisspeptin modulates fertilization capacity of mouse spermatozoa. Reproduction 2014, 147, 835–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Jin, Y.H.; Oh, S.Y.; Kwon, J.Y. Kisspeptin2 stimulates the HPG axis in immature Nile tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 202, 31–38. [Google Scholar] [CrossRef]
- Gloria, A.; Contri, A.; Mele, E.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptin Receptor on the Sperm Surface Reflects Epididymal Maturation in the Dog. Int. J. Mol. Sci. 2021, 22, 120. [Google Scholar] [CrossRef]
- Yue, Z.; Yu, M.; Zhang, X.; Wang, J.; Ru, S. The anti-androgenic effect of chronic exposure to semicarbazide on male Japanese flounder (Paralichthys olivaceus) and its potential mechanisms. Comp. Biochem. Physiol. C Toxicol. Pharm. 2018, 210, 30–34. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Reference | Formulation | Dose | Duration of Treatment | Administration | Animal Model | Main Findings |
|---|---|---|---|---|---|---|
| Liu et al. [7] | Carbendazim | 0.1, 1, and 10 mg/kg | 5 weeks | Oral gavage | ICR mice (n = 30 per group) | Decreased sperm concentration and motility Impaired spermatogenesis Decreased estrogen signaling Alterations in histone and DNA methylation |
| Salem et al. [43] | Carbendazim | 100 mg/kg | 8 weeks | Oral gavage | Swiss albino rats (n = 10 per group) | Decreased sperm concentration, motility, and viability Increased percentage of morphological abnormal sperm Decreased concentrations of serum testosterone, gonadotropins, and inhibin B Increased oxidative stress Alterations on the seminiferous tubules structure |
| Elsharkawy et al. [44] | Mancozeb | 100 mg/kg | 12 weeks | Oral gavage (twice per week) | White New Zealand rabbit (Oryctolagus cuniculus) (n = 9 per group) | Decreased concentrations of serum testosterone and gonadotropins Decreased sperm viability Increased percentage of morphological abnormal sperm Disruption of the germinal epithelium Vacuolization of Leydig cells |
| Mohanty et al. [45] | Mancozeb | 0.14 and 0.28 mg/day | 30 days | Oral (mixed with food) | Red Avadavat (Amandava amandava) (n = 9 per group) | Decreased concentrations of serum testosterone and gonadotropins Impaired gonadal development Altered hypothalamic expression of GnRH Disruption of the HPT axis |
| Meng et al. [46] | Methomyl | 0.2, 2, 20, and 200 μg/L | 30 days | Dissolved in water | Nile tilapia (Oreochromis niloticus) (n = 30 per tank, n = 3 per condition) | Altered expression of HPT-related genes in the hypothalamus, pituitary, and testis at 20 and 200 μg/L The effects of 200 μg/L were considered irreversible |
| Yue et al. [59] | Semicarbazide | 1, 10, and 100 μg/L | 130 days | Dissolved in water | Japanese flounder (Paralichthys olivaceus) (n = 6 per group) | Decreased expression of genes involved in steroidogenesis Decreased concentration of serum testosterone and estradiol Disruption of the HPT axis Alterations in the kiss/gpr54 system and GABA synthesis |
| Guo et al. [47] | Ziram | 2 and 4 mg/kg/day | 4 weeks | Oral gavage | Sprague–Dawley rats (n = 6 per group) | Decreased concentrations of serum testosterone and FSH Decreased Leydig cell number |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, S.; Silva, R.; Carrageta, D.F.; Alves, M.G.; Seco-Rovira, V.; Oliveira, P.F.; de Lourdes Pereira, M. Carbamate Pesticides: Shedding Light on Their Impact on the Male Reproductive System. Int. J. Mol. Sci. 2022, 23, 8206. https://doi.org/10.3390/ijms23158206
Moreira S, Silva R, Carrageta DF, Alves MG, Seco-Rovira V, Oliveira PF, de Lourdes Pereira M. Carbamate Pesticides: Shedding Light on Their Impact on the Male Reproductive System. International Journal of Molecular Sciences. 2022; 23(15):8206. https://doi.org/10.3390/ijms23158206
Chicago/Turabian StyleMoreira, Sílvia, Ricardo Silva, David F. Carrageta, Marco G. Alves, Vicente Seco-Rovira, Pedro F. Oliveira, and Maria de Lourdes Pereira. 2022. "Carbamate Pesticides: Shedding Light on Their Impact on the Male Reproductive System" International Journal of Molecular Sciences 23, no. 15: 8206. https://doi.org/10.3390/ijms23158206
APA StyleMoreira, S., Silva, R., Carrageta, D. F., Alves, M. G., Seco-Rovira, V., Oliveira, P. F., & de Lourdes Pereira, M. (2022). Carbamate Pesticides: Shedding Light on Their Impact on the Male Reproductive System. International Journal of Molecular Sciences, 23(15), 8206. https://doi.org/10.3390/ijms23158206