Abstract
Chronic inflammation caused by liver damage or infection plays an important role in the development and progression of hepatocellular carcinoma (HCC). The activation of Toll-like receptors 4 (TLR4) is involved in HCC tumorigenesis. Moreover, high TLR4 expression in HCC has been linked to poor prognosis. Although the expression of TLR4 in HCC is relatively low compared to hematopoietic cells, it is important to explore the molecular mechanism leading to the elevation of TLR4 in HCC. In this study, we aimed to investigate the positive regulating loop for TLR4 expression in HCC in response to chronic inflammation. Our results confirm that the mRNA expression of TLR4 and proinflammatory cytokines, including interleukin 6 (IL6) and C-C motif chemokine ligand 2 (CCL2), positively correlate in human HCC samples. High TLR4 expression in HCC is more susceptible to lipopolysaccharide (LPS); TLR4 activation in HCC provides growth and survival advantages and thus promotes tumorigenesis. It has been shown that the LIN28/let-7 microRNA (miRNA) axis is a downstream effector of the TLR4 signal pathway, and let-7 miRNA is a potential post-transcriptional regulator for TLR4. Thus, we investigated the correlation between TLR4 and LIN28A mRNA and let-7g miRNA in HCC clinical samples and found that the expression of TLR4 was positively correlated with LIN28A and negatively correlated with let-7g miRNA. Moreover, by culturing PLC/PRF5 (PLC5) HCC cells in low-dose LPS-containing medium to mimic chronic inflammation for persistent TLR4 activation, the mRNA and protein levels of TLR4 and LIN28A were elevated, and let-7g miRNA was decreased. Furthermore, the 3’ untranslated region (3’UTR) of TLR4 mRNA was shown to be the target of let-7g miRNA, suggesting that inhibition of let-7g miRNA is able to increase TLR4 mRNA. While parental PLC5 cells have a low susceptibility to LPS-induced cell growth, long-term LPS exposure for PLC5 cells leads to increased proliferation, cytokine expression and stemness properties. In conclusion, our studies demonstrate positive feedback regulation for chronic TLR4 activation in the modulation of TLR4 expression level through the LIN28A/let-7g pathway in HCC and suggest a connection between chronic inflammation and TLR4 expression level in HCC for promoting tumorigenesis.
1. Introduction
Hepatocellular carcinoma (HCC) is among the most common cancers worldwide, and major risk factors include chronic liver damage, which is mainly due to chronic infection with hepatitis B virus (HBV) and hepatitis C virus (HCV); alcohol abuse; metabolic syndrome; high calorie intake; diabetes; and obesity [1,2,3]. Under chronic damage and infection, inflammatory response is activated to produce pro-inflammatory cytokines or inflammation-related genes through transcription factors such as nuclear factor-κB (NF-κB) and signal transducer and activator of transcription 3 (STAT3). Additionally, several signaling pathways activated by inflammation have been indicated as being involved in carcinogenesis, such as the mitogen-activated protein kinases (MAPKs), NF-κB and phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathways [4].
Toll-like receptors (TLRs) are primary sensors of exogenous pathogen-associated molecular patterns (PAMPs) and endogenous damage-associated molecular patterns (DAMPs), and thereby play important roles in inflammatory responses [5]. As the first member of the TLR family to be discovered, TLR4 has been indicated to be involved in mediating innate immunity and inflammation and is linked to tumorigenesis. In general, TLR4 signaling is mediated by the recruitment of adaptor proteins, such as MyD88 and TRIF, to TIR domains in the cytoplasmic portion of the receptors. The dimerized adaptors, in turn, recruit a variety of signaling proteins that activate different effector pathways, including IκB kinase (IKK), p38 MAPK, c-Jun N-terminal kinase (JNK), PI3K and IKK-like kinases, such as TANK-binding kinase 1 (TBK1) and IKK [6]. In cancer cells such as lung, liver, gastric, pancreatic, ovarian, and colon cancer, TLR4 expression is elevated [7,8,9,10,11]. The activation of TLR4 enhances tumor growth or metastasis and protects cancer cells from apoptosis or immune cell-induced lysis [12,13].
MicroRNAs (miRNAs) are able to regulate the 3′ untranslated regions (3’UTRs) of target mRNA through post-transcriptional modification and participate in numerous physiological and pathological processes. In recent years, miRNAs have been identified as important regulators for TLR pathways [14]. For instance, miR-105 inhibits TLR2 expression in human oral keratinocytes [15], and miR-203 in pancreatic cancer-secreted exosomes inhibits TLR4 in dendritic cells [16]. As the largest and the most studied of all miRNA families, the let-7 family has been indicated as being involved in the regulation of TLR4 expression. In cultured human cholangiocytes, let-7i regulates TLR4 expression via post-transcriptional suppression [17]. let-7e is positively regulated by Akt1 in LPS-treated macrophages and targets TLR4 [18]. In human gastric epithelial cells with Helicobacter pylori infection, the TLR4 signaling pathway is activated and promotes LIN28 expression to inhibit let-7b, which is complementary to the 3′UTR of TLR4 mRNA and regulates TLR4 expression [19].
LIN28, which has two homologs, LIN28A and LIN28B, is an RNA-binding protein and mediates biological functions by binding to the target mRNA or miRNA. LIN28 has been shown to bind to the loop portion of pri-let-7 and pre-let-7 to inhibit the activities of Drosha and Dicer, and mediate the terminal uridylation of pre-let-7, leading to the blockage of matured let-7 [20,21,22,23,24,25]. As the main downstream effector of TLR4-mediated IKK signal, NF-κB directly activates the transcription of LIN28 [26,27]. Meanwhile, it has also been shown that interleukin 6 (IL6), an inflammatory cytokine induced by TLR4, mediates STAT3 activation to promote the transcription of LIN28 [28].
TLR4 is a critical receptor responsible for inflammatory response and is associated with HCC progression [29,30]. The expression of TLR4 has previously been associated with enhanced invasion and metastasis abilities and poor prognosis in HCC [31,32]. Although the expression of TLR4 in HCC cells is relatively low compared to expression levels in hematopoietic cells, we are interested in investigating the molecular mechanism that leads to the elevation in TLR4 expression in HCC under chronic inflammation. As human HCC cells are extremely heterogeneous, the correlation of TLR4 with inflammatory cytokines was investigated; we confirmed that the expression of TLR4 is positively correlated with inflammatory cytokines such as IL6 and C-C motif chemokine ligand 2 (CCL2), and that TLR4 activation promotes HCC. Next, to explore the post-transcriptional regulation of TLR4 expression, the LIN28/let7 axis, which is involved in tumor development and is associated with inflammation, is a potential molecular target. Thus, the relationship between TLR4 and LIN28/let-7 was investigated. In HCC tissues, a positive correlation was found between TLR4 and LIN28A, while a negative correlation was found between TLR4 with let-7g. With long-term TLR4 activation in PLC5 cells, LIN28A is increased and let-7g miRNA is decreased. Moreover, we identified that let-7g directly targets TLR4 mRNA and inhibits TLR4 expression. Our results suggest that long-term inflammation leads to the upregulation of TLR4 expression and downstream effectors through a positive feedback loop and contributes to HCC development.
2. Results
2.1. The Expression of TLR4 Is Positively Correlated with IL6 and CCL2 in Clinical Samples
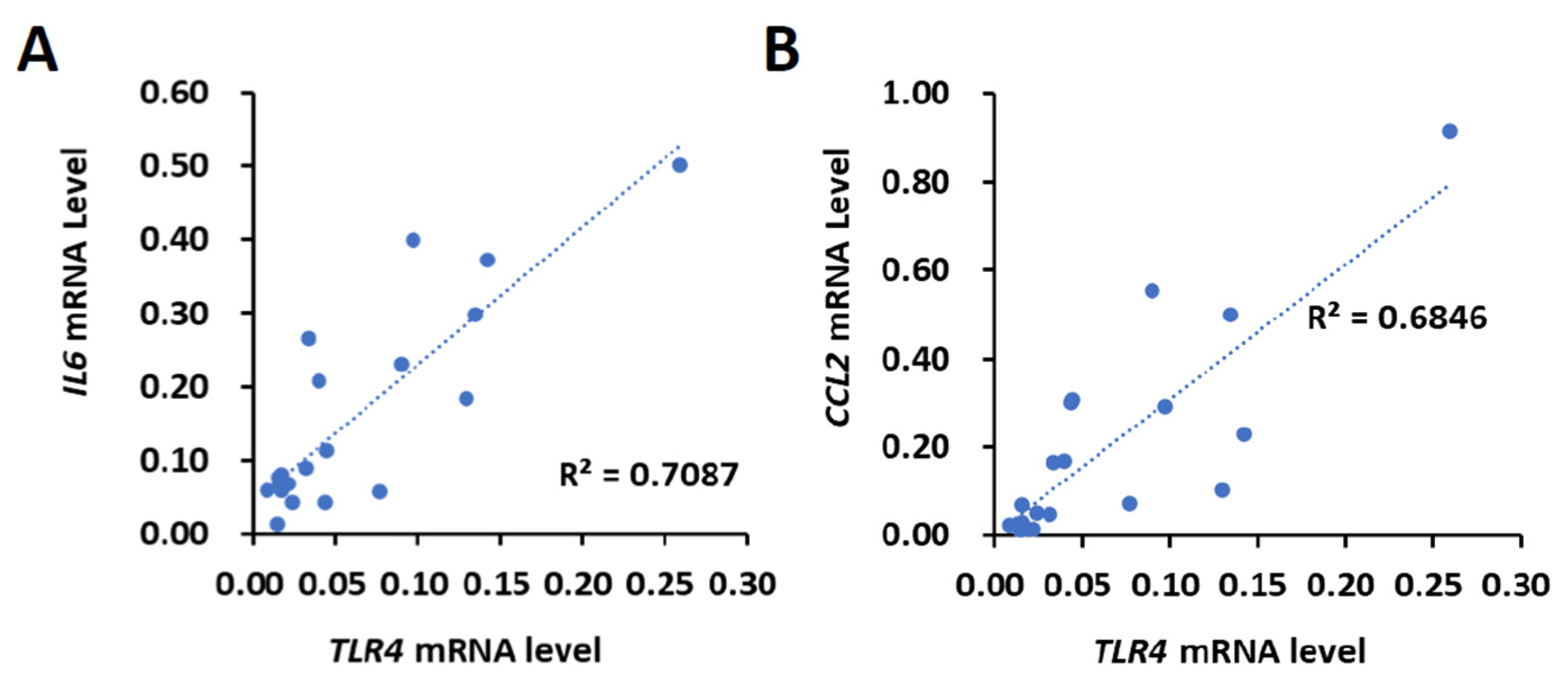
First, the correlation between TLR4 and IL6 or CCL2 levels was investigated. Using the 20 HCC tissues that were collected from HBV- and HCV-negative male HCC patients by the Taiwan Liver Cancer Network, the mRNA expression of TLR4, IL6 and CCL2 was determined using the quantitative polymerase chain reaction (qPCR). The results show that the expression levels of TLR4, IL6 and CCL2 varied between human HCC samples, indicating that the upregulation of TLR4, IL6 and CCL2 might not be a typical event in the early stages of HCC (stage I or II) (Supplementary Figure S1). We then used Pearson’s correlation analysis to test whether there was a significant relationship between the mRNA expression levels of TLR4, IL6 and CCL2. Our results show a positive correlation between the mRNA expression levels of TLR4 and IL6 (R = 0.8419, p < 0.0001) and CCL2 (R = 0.8274, p < 0.0001) (Figure 1).
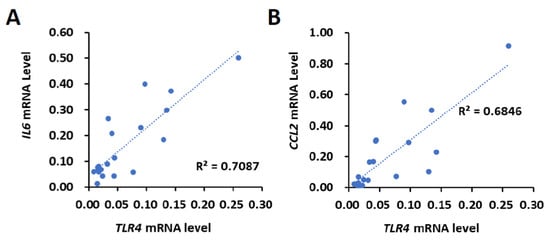
Figure 1.
Positive correlation of TLR4 with IL6 or CCL2 mRNA in HCC clinical samples. Correlation between the mRNA expression of TLR4 with (A) IL6 and (B) CCL2 in HCC patients. Pearson’s correlation analyses were performed. Data are presented as each value (R2, correlation coefficient).
2.2. Activation of TLR4 in HCC Leads to Survival Advantage
Previous studies have shown that TLR4 is critical for promoting HCC tumorigenesis [26,28]. By screening the expression levels of TLR4 mRNA and protein in five different liver cancer cells, Huh7, PLC/PRF5 (PLC5), Hep3B, HepG2 and SK-Hep1, HepG2 and Hep3B were examined. Huh7, PLC5, Hep3B and HepG2 cell lines were derived from biopsies of HCC patients, and SK-Hep1 cells were derived from the ascitic fluid of a liver adenocarcinoma patient. Five HCC cell lines with different genetic backgrounds, such as p53 expression and HBV infection, exhibited different levels of TLR4 expression. Huh7 and Hep3B cells expressed high levels of TLR4 mRNA, while PLC5, HepG2 and SK-Hep1 cells had relatively low TLR4 mRNA expression. For protein levels, Huh7 cells expressed high levels of TLR4, while PLC5, Hep3B, HepG2 and SK-Hep1 cells demonstrated low TLR4 expression levels (Supplementary Figure S2). Hep3B cells show inconsistent expression levels between mRNA and protein; thus, Huh7 cells were used to represent cells with high TLR4 expression levels, while PLC5 cells were used to represent cells with low TLR4 expression levels.
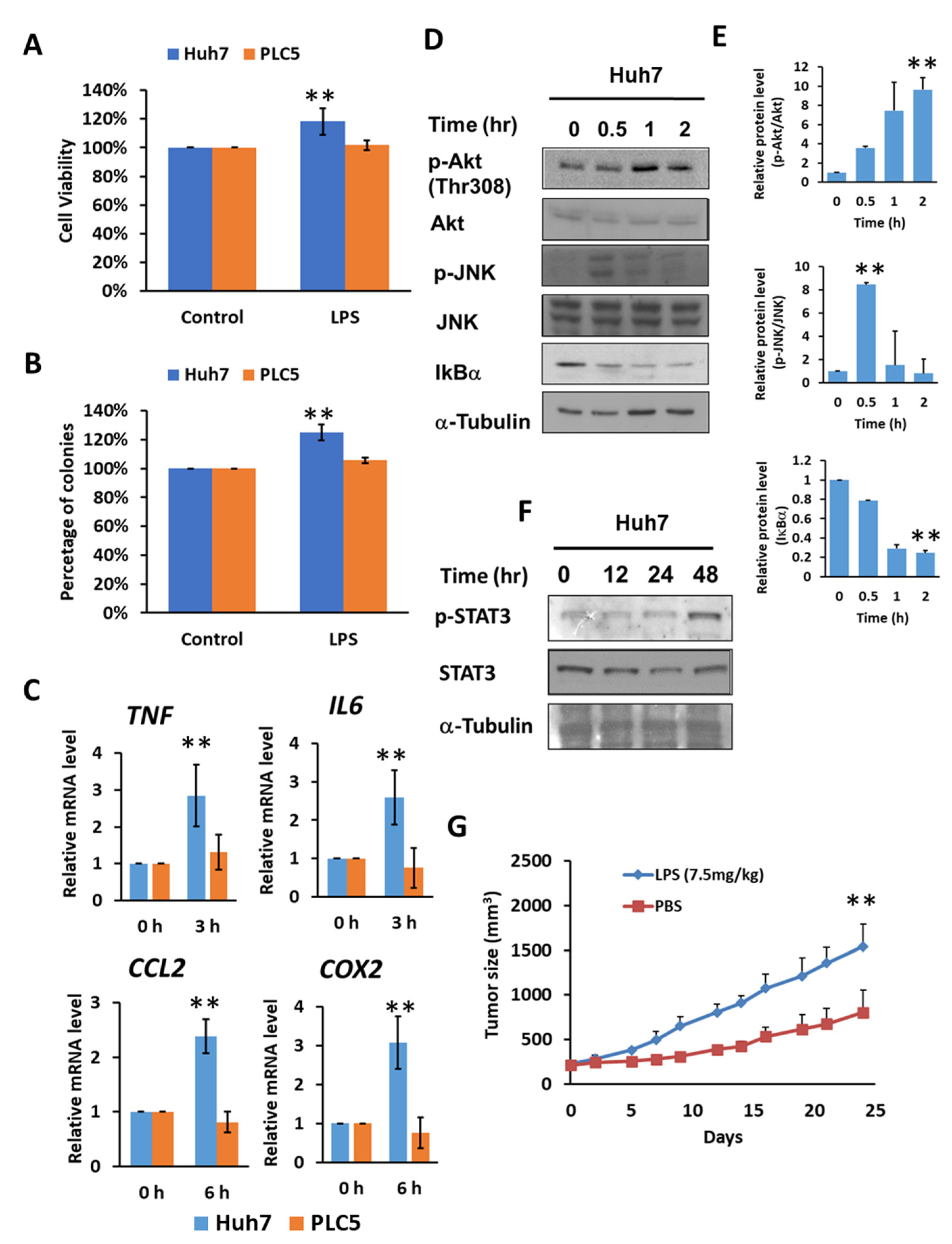
By activating TLR4 signaling with lipopolysaccharide (LPS) treatment, Huh7 cells demonstrated approximately 20 percent higher cell viability and a 25 percent increase in colony formation; however, no significant difference in cell viability or colony formation was found in PLC5 cells (Figure 2A,B). Meanwhile, Huh7 cell growth promoted by LPS was diminished in TLR4-knockdown cells (Supplementary Figure S3), indicating an LPS-mediated enhancement in cell viability and TLR4-dependent colony formation in HCC cells. The downstream signaling pathways of TLR4, such as NF-κB, MAPKs and PI3K pathways, can further help cell growth and proliferation, and may also assist in the progression of cancer. We confirmed that TLR4 activation by LPS in Huh7 cells with high TLR4 expression, not PLC5 cells, was able to mediate the production of inflammatory cytokines and chemokines, including tumor necrosis factor (TNF), IL6, and CCL2 and Cyclooxygenase 2 (COX2) (Figure 2C), and the activation of Akt, JNK, IKK (Figure 2D,E) and STAT3 signals (Figure 2F), but this was not the case in PLC5 cells with low TLR4 expression. Moreover, LPS treatment is able to promote tumor growth in vivo in a Huh7 xenograft mice model (Figure 2G).
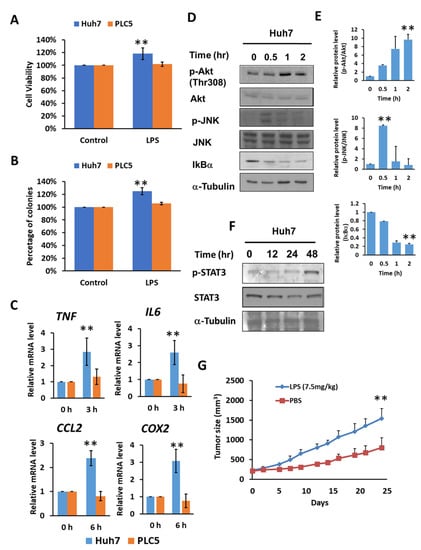
Figure 2.
TLR4 activation in Huh7 HCC cells leads to growth and survival advantages. (A) Cell viability and (B) clonogenic ability of Huh7 and PLC5 cells with TLR4 engagement. Huh7 and PLC5 cells incubated with or without LPS (10 ng/mL) were placed on 96-well plates for 4 days or on 6-well plates for 14 days. Cell viability was measured with MTT assays. Clonogenic ability was determined by colony formation, which was visualized by crystal violet. The relative viability and clonogenic ability of cells without LPS treatment were set as 100 percent. Results are mean ± SD for six separate experiments. (C) Induction of TNF, IL6, CCL2 and COX2 mRNA in Huh7 and PLC5 cells with LPS stimulation. Huh7 or PLC5 cells were treated with or without LPS for indicated time periods. RNA was extracted, and the expression of the indicated mRNAs was measured by qPCR. The relative mRNA level in Huh7 cells with LPS treatment at 0 h was set as 1.0. Results are mean ± SD for three separate experiments. (D) Activation of Akt, JNK and IKK pathways in Huh7 cells in response to LPS. Cells were treated with LPS and collected at the indicated time points. Akt and JNK phosphorylation and IκBα degradation were analyzed by means of immunoblotting. The results are represented by three independent experiments. (E) The quantification of relative protein levels for p-Akt/Akt, p-JNK/JNK and IκBα in each condition. Error bars show standard deviation. (F) Activation of STAT3 in Huh7 cells with LPS treatment. Cells were treated with LPS for the indicated times. Cell lysates were analyzed by immunoblotting for STAT3 phosphorylation. The results are represented by three independent experiments. (G) Effect of LPS on HCC cell growth in vivo. SCID mice were inoculated with 2 × 106 Huh7 cells s.c. in the right flank. Phosphate buffered saline (PBS) or LPS (2.5 mg/kg) was administrated i.p. once weekly, and treatment was started on day 1, when tumors were measurable (n = 6 for each group). Caliper measurements were performed to estimate the tumor volume (mean ± SD mm3). (** p < 0.01).
2.3. Analysis of the Correlation between TLR4 and LIN28A/Let-7g Axis in HCC Clinical Samples
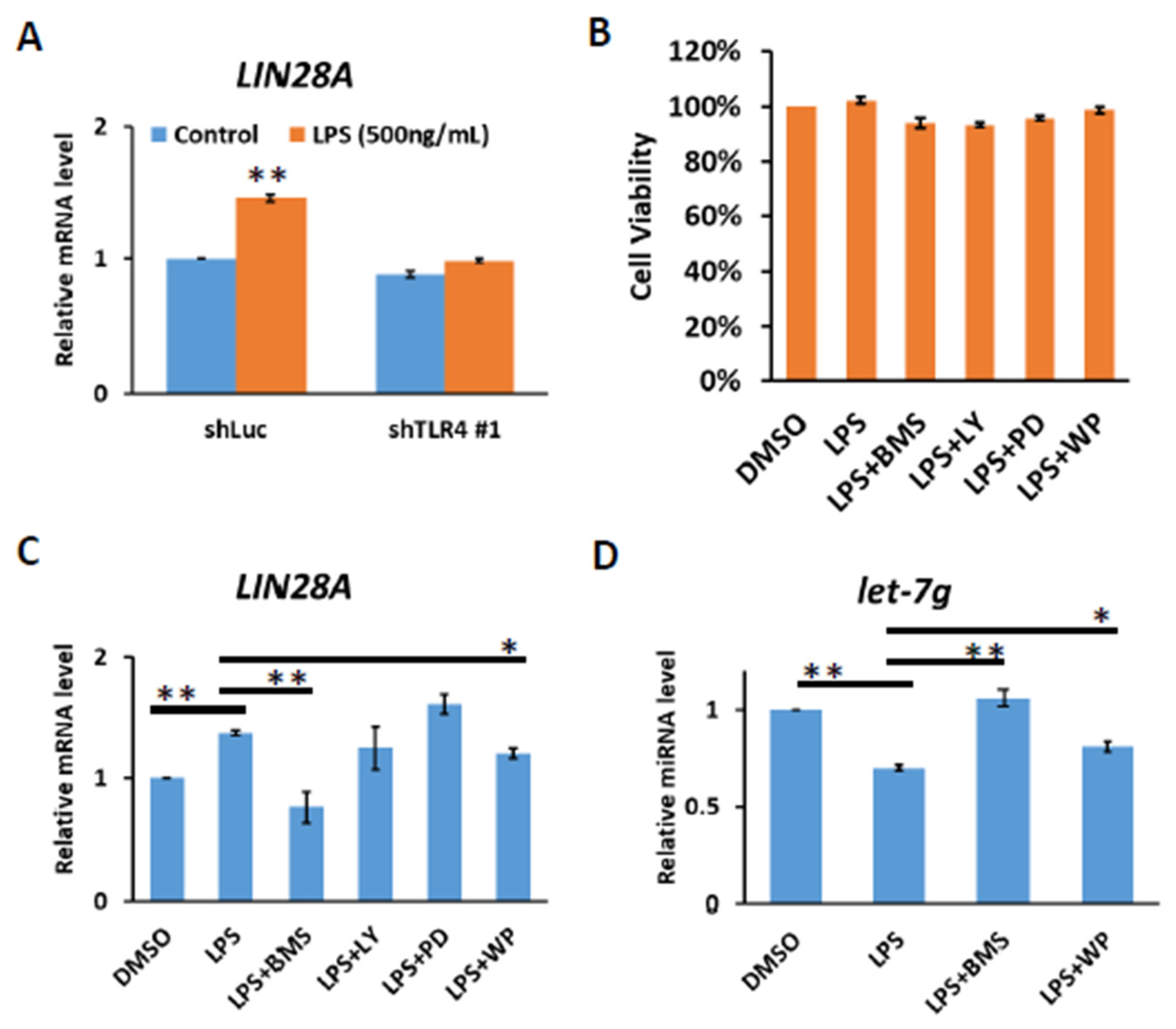
It has previously been demonstrated that the LIN28 family is transcriptionally regulated via direct promoter binding by transcription factors, such as NF-κB and STAT3, the downstream effectors of TLR4 signaling [26,27,28]. We confirmed that TLR4 activation by LPS in Huh7 cells leads to increases in LIN28A mRNA and that LPS-mediated induction of LIN28A expression is diminished in TLR4-knockdown cells (Figure 3A). To examine whether NF-κB and STAT3 were responses to the induction of LIN28A mRNA, Huh7 cells were treated with LPS in the presence of various inhibitors to block the IKK, PI3K, MAPK and STAT3 pathways. After 4 hours of treatment with dimethyl sulfoxide (DMSO) or inhibitors, no significant effect on cell viability was found (Figure 3B). The results show that the induction of LIN28A mRNA was blocked by an IKK inhibitor, BMS345541, and a STAT3 inhibitor, WP1066, indicating that LPS-mediated LIN28A expression is NF-κB- and STAT3-dependent (Figure 3C). Previous studies suggested that Lin28A inhibits biogenesis of let-7 miRNA by the direct binding to pre-let-7 or pri-let-7 and inhibits its processing [20,21,22,23,24,25]. Therefore, the inhibition of IKK or STAT3 pathways with BMS345541 or WP1066 leads to the elevation of let-7g miRNA (Figure 3D).
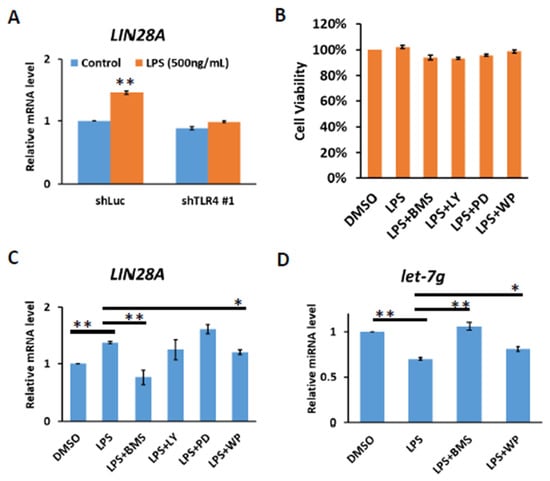
Figure 3.
TLR4 activation induces IKK-dependent LIN28A expression. (A) Induction of LIN28A mRNA of Huh7 cells with or without TLR4 silencing in response to LPS. The cells were treated with LPS (500 ng/mL) for 4 h and RNA was extracted to be measured by qPCR. (B) Cell viability of Huh7 cells. Huh7 incubated with LPS (10 ng/mL) in the presence of indicated inhibitors was placed on 96-well plates for 4 h. Cell viability was measured with MTT assays. (C) LIN28A mRNA and (D) let-7g miRNA expression regulated by LPS depends on IKK and STAT3 pathways. Huh7 cells were treated with LPS (500 ng/mL) in the presence of indicated inhibitors, including IKK inhibitor (BMS345541 (20 μM)), PI3K inhibitor (LY294002 (20 μM)), MEK inhibitor (PD98059 (50 μM)) and STAT3 inhibitor (WP1066 (10 μM)), for 4 h. RNA was extracted, and the expression levels of the indicated mRNAs and miRNA were measured by qPCR. The relative mRNA and miRNA level in Huh7 cells without LPS treatment was set as 1.0. Results are mean ± SD for two separate experiments. (* p < 0.05, ** p < 0.01).
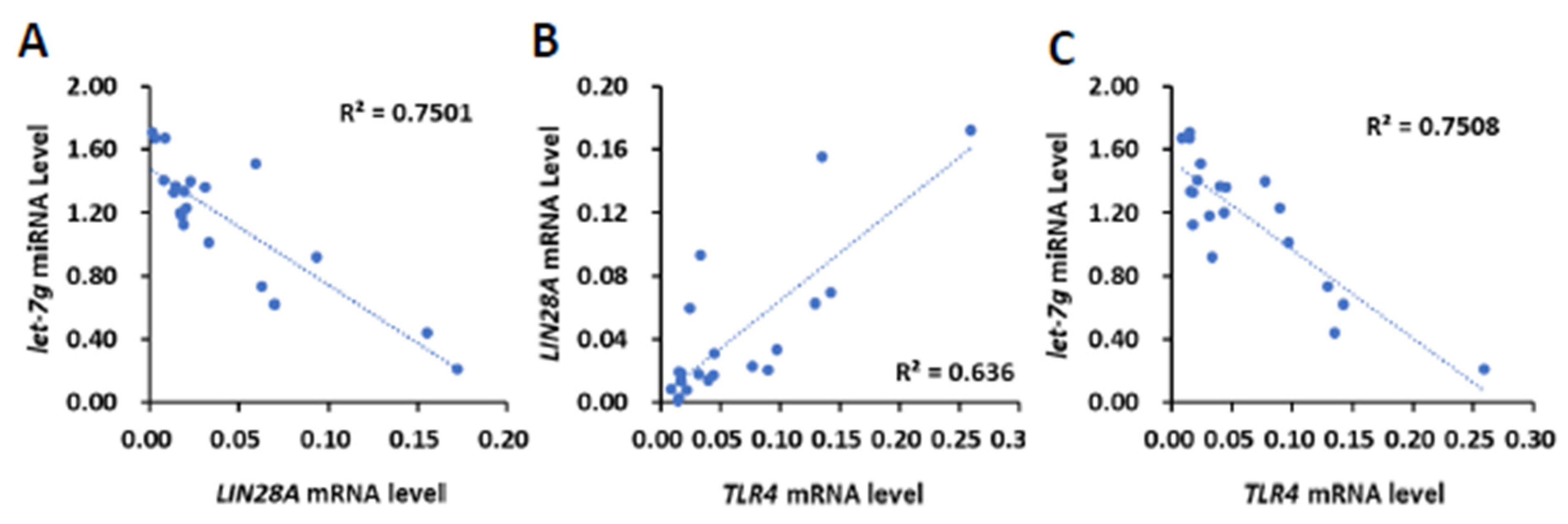
To clinically investigate whether TLR4 mRNA expression is associated with the expression LIN28 mRNA or let-7 miRNA, 20 human HCC tissues (Figure 1) were further analyzed for the expression of LIN28A and LIN28B mRNA and let-7b, let-7i and let-7g miRNA (Supplementary Figure S4). We focused on LIN28A for the correlation analysis due to the low expression level of LIN28B in clinical samples. By employing Pearson’s correlation analysis, we found that the expression level of LIN28A mRNA was also inversely related to let-7g miRNA (R = −0.7259, p < 0.001; Figure 4A), but not let-7b and let-7i miRNA (R = −0.1402 and −0.1213, respectively). By analyzing TLR4’s association with LIN28A mRNA and let-7g miRNA, we found that there was a positive correlation between TLR4 and LIN28A mRNA level (R = 0.7975, p < 0.0001), and an inverse correlation between TLR4 mRNA and let-7g miRNA (R = −0.6662, p < 0.005; Figure 4B,C). In addition to human HCC samples, tumor cells from a model with DEN-induced HCC mice were isolated and analyzed for expression of Tlr4 and Lin28a mRNA and let-7g miRNA level (Supplementary Figure S5A–C). The results show that Tlr4 mRNA level was positively correlated with Lin28a (R = 0.8972, p < 0.001), and inversely correlated with let-7g miRNA (R = −0.7836, p < 0.01; Supplementary Figure S5D–F). Based on these results, we demonstrated that TLR4 and LIN28A mRNA are positively correlated, while TLR4 and let-7g miRNA are negatively correlated, confirming the association between TLR4 and the LIN28A/let-7g axis.
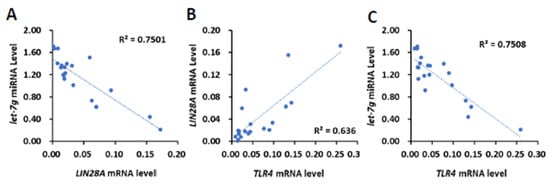
Figure 4.
Positive correlation of TLR4 with LIN28A mRNA and inverse correlation of TLR4 and LIN28A mRNA with let-7g miRNA in clinical HCC samples. (A) Correlation between the expression of LIN28A mRNA with let-7g miRNA in HCC patients, and correlation between the expression of TLR4 mRNA and (B) LIN28A mRNA and (C) let-7g miRNA. Pearson’s correlation analyses were performed. Data are presented as each value (R2, correlation coefficient).
2.4. HCC with Long-Term TLR4 Activation Leads to Upregulation of TLR4 and LIN28A and Downregulation of Let-7g miRNA
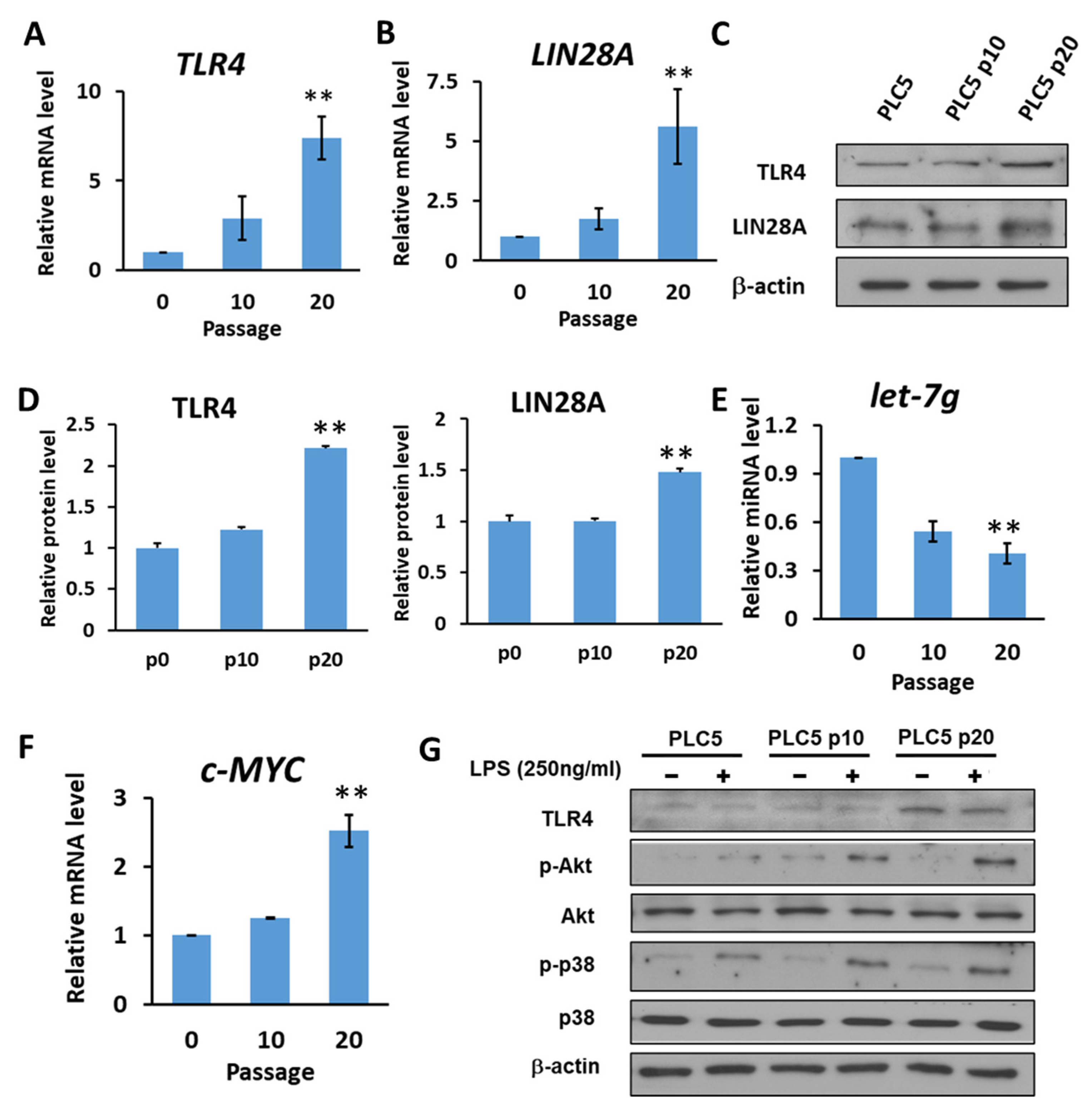
It has also been shown that let-7 miRNA, which is inhibited by LIN28, might target TLR4 mRNA. Based on the correlation between TLR4, LIN28A and let-7g clinically, we investigated whether a positive feedback loop links chronic inflammatory response with upregulation of TLR4 expression in HCC cells. Human PLC5 cancer cells with low TLR4 expression were cultured in the presence of LPS (10 ng/mL) to mimic chronic inflammatory response. After 10 and 20 generations of cultivation, the results show that the expression of TLR4 and LIN28A mRNA was increased in PLC5 cells cultured with LPS-containing medium (Figure 5A,B). The protein expression levels of TLR4 and LIN28A were detected by Western blotting (Figure 5C). The results show that TLR4 and LIN28A expression levels were increased with long-term LPS exposure (Figure 5D). Next, the expression level of let-7g miRNA was examined. Our results show that PLC5 cells under chronic inflammation increased the expression of TLR4 and LIN28A and caused a reduction in let-7g miRNA (Figure 5E). Additionally, c-MYC, which is an important transcription factor for stemness and is inhibited by let-7 miRNA, was analyzed to confirm the downregulation of let-7 miRNA. The data show that c-MYC increased in PLC5 cells after long-term LPS exposure (Figure 5F). As the expression of TLR4 gradually increased under the influence of long-term LPS treatment, the activation of TLR4-induced signaling was examined. The experimental results show that, compared with PLC5 cells that had not undergone long-term LPS exposure, TLR4-mediated signaling, including Akt and p38, in PLC5 cells with long-term LPS exposure was activated in response to LPS stimulation for 30 min (Figure 5G), indicating that long-term LPS stimulation to mimic the chronic inflammatory environment is able to promote TLR4 expression and sensitize TLR4-induced signals in HCC cells.
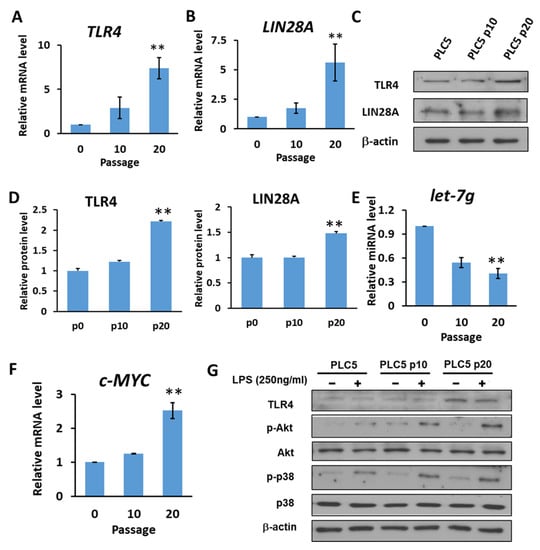
Figure 5.
HCC with long-term LPS exposure leads to the upregulation of TLR4 and LIN28A, and the downregulation of let-7g miRNA. (A) TLR4 and (B) LIN28A mRNA levels in PLC5 cells with or without long-term LPS exposure for indicated passages. (C) Protein level of TLR4 and LIN28A in PLC5 cells with LPS exposure for indicated passages. The expression levels of indicated proteins were detected with immunoblotting. The results are represented by three independent experiments. (D) The quantification of TLR4 and LIN28A protein levels compared to the α-actin control in each condition. Error bars show standard deviation. (E) The expression of let-7g miRNA level in PLC5 cells with or without LPS exposure for indicated passages. (F) c-Myc mRNA level in PLC5 cells with or without LPS exposure for indicated passages. The mRNA and miRNA levels were detected with qPCR. The relative mRNA or miRNA level in PLC5 cells without LPS treatment was set as 1.0. Results are mean ± SD for three separate experiments. (G) Activation of Akt and p38 pathways in PLC5 cells with or without long-term LPS exposure. Cells were treated without or with LPS (250 ng/mL) for 30 min. Akt and p38 phosphorylation was analyzed by means of immunoblotting. The results are represented by two independent experiments. (** p < 0.01).
2.5. TLR4 Is the Direct Target to Be Regulated by MicroRNA Let-7g
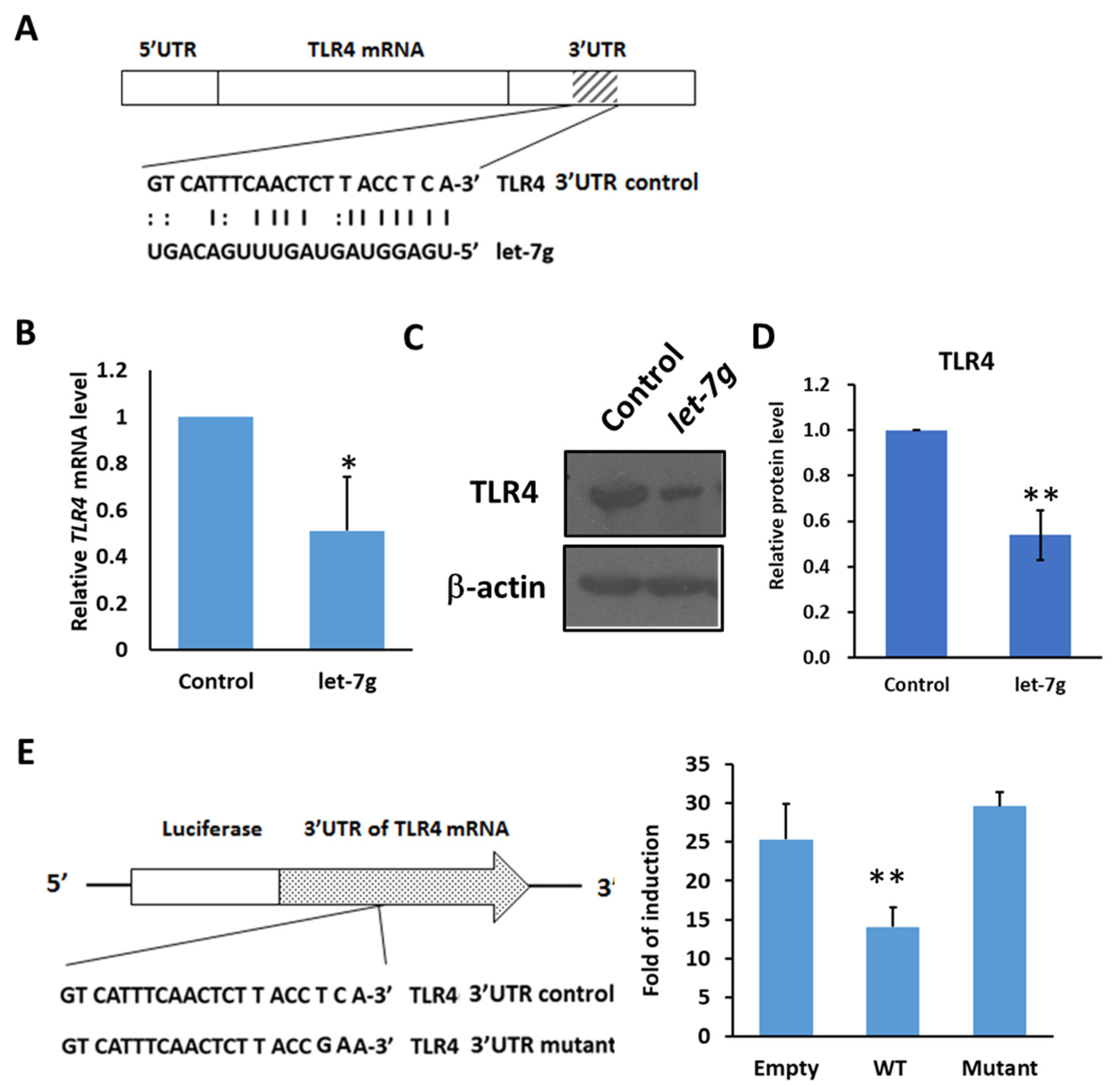
In epithelial cells, it was found that let-7i or let-7b miRNAs target TLR4 mRNA and regulate the expression of TLR4 [17,19]. With the high sequence similarity of let-7g with let-7b and let-7i, it is likely that let-7g miRNA can regulate the expression of TLR4 mRNA. With microRNA.org (http://www.microrna.org) (accessed on 20 September 2018), the 3′UTR of TLR4 mRNA was predicted as a potential seed region for let-7g miRNA (Figure 6A). To determine the relationship between let-7g miRNA and TLR4 mRNA, let-7g was transfected into Huh7 cells, and the expression level of TLR4 mRNA and protein level were examined. It can be seen that TLR4 mRNA was inhibited by let-7g miRNA (Figure 6B). Meanwhile, the protein expression level of TLR4 decreased with the overexpression of let-7g miRNA (Figure 6C,D). To further verify that let-7g regulates the 3′UTR of TLR4 mRNA through post-transcriptional modification, a luciferase reporter assay was performed. It was found that luciferase activity in the construct with the 3′UTR of TLR4 mRNA was reduced by about 50% compared with the empty construct without the 3′UTR of TLR4 mRNA in the presence of pre-let-7g miRNA (Figure 6E). With the mutated 3′UTR of TLR4 in the binding site of let-7g, luciferase activity was not decreased by pre-let-7g miRNA (Figure 6E). These data indicate that let-7g inhibits TLR4 mRNA through post-transcriptional modification.
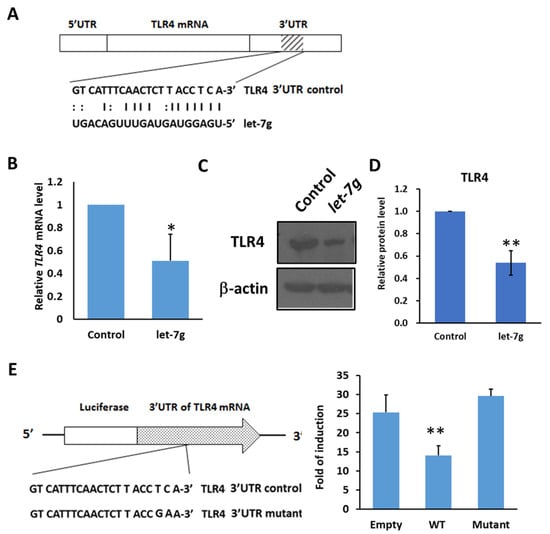
Figure 6.
TLR4 mRNA is targeted by let-7g miRNA. (A) Schematic of the seed region match between hsa-let-7g and the putative TLR4 3′UTR. MicroRNA.org was used to analyze and predict the possible seed region of has-let-7g on the 3′UTR of human TLR4 mRNA. (B) Huh7 cells were transfected with control or hsa-let-7g precursor-expressed vector and incubated for 1 to 3 days as indicated. The levels of endogenous TLR4 mRNA were quantified with qPCR. (C) Huh7 cells were transfected with control or hsa-let-7g miRNA precursor-expressed vector, incubated for 2 days, and harvested for the Western blotting of TLR4 protein levels. The results are represented by two independent experiments. (D) The quantification of relative protein levels for TLR4 in each condition. Error bars show standard deviation. (E) The post-transcriptional regulation of TLR4 mRNA by let7-g miRNA with luciferase reporter assay. The positions of three seed match sites for let-7g and TLR4 3′UTR in the luciferase reporter construct pmirGLO-TLR4-3′UTR are indicated (left). Huh7 cells in 96-well plates were co-transfected with pre-let-7g miRNA-expressed vector and the indicated reporter constructs, including empty, wild-type TLR4 3′UTR, and mutated TLR4 3′UTR, respectively, in the downstream of Firefly luciferase gene. Firefly and Renilla luciferase activities were measured at 24 h post-transfection. The data represent the means from three independent experiments. (* p < 0.05, ** p < 0.01).
2.6. HCC with Long-Term LPS Exposure Leads to Increase Proliferation and Stemness Properties
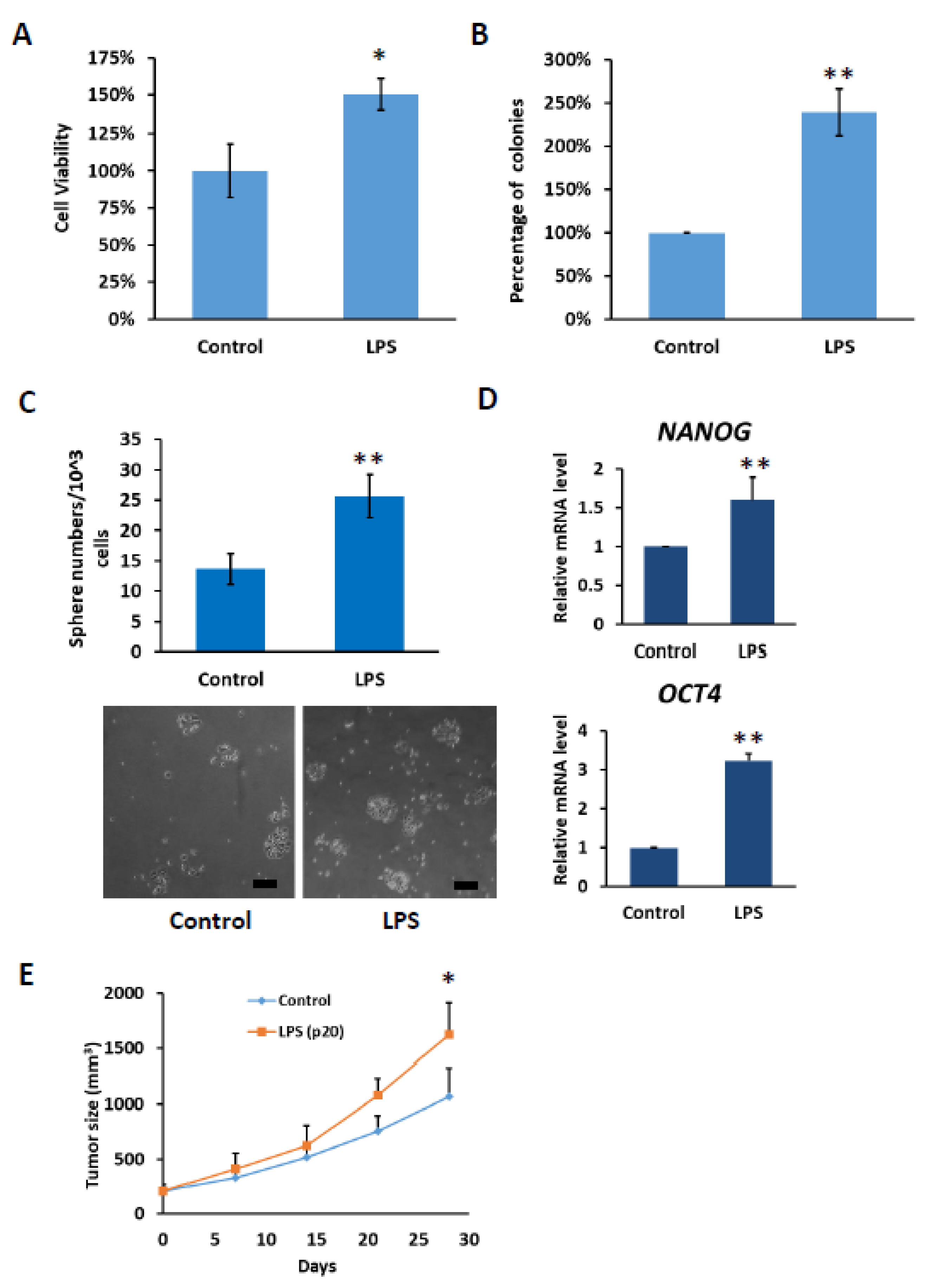
Based on our results, it was observed that under the influence of chronic inflammation, the expression of TLR4 gradually increased. Therefore, we investigated whether the increase in TLR4 expression promoted HCC tumorigenesis. Compared with control PLC5 cells, PLC5 cells with long-term LPS exposure exhibited higher cell viability in the MTT assay and promoted colony formation ability (Figure 7A,B). Furthermore, TLR4 activation leads to the downregulation of let-7 miRNA, which is responsible for controlling stemness properties. Elevation of TLR4 expression in LPS-treated PLC5 cells promotes the formation of spheroid cells, indicating an increase in cancer precursor cells (Figure 7C). The spheres were collected and the mRNA levels of stemness-associated genes, including NANOG and OCT4, were analyzed. Consistent with the sphere formation assay, NANOG and OCT4 mRNA levels were increased in the spheres of LPS-exposed PLC5 cells (Figure 7D). Next, tumor growth in the xenograft mice model was examined. From the results, it can be seen that tumor growth increased with PLC5 cells cultured in LPS-containing medium for 20 passages compared with tumors with PLC5 cells cultured in control medium for 20 passages, suggesting that HCC cancer cells cultured under chronic inflammation promote tumorigenesis (Figure 7E).
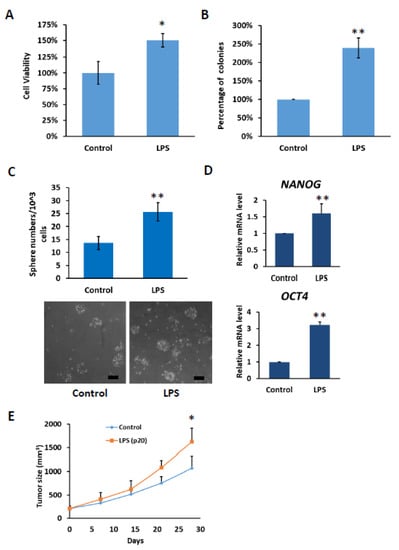
Figure 7.
Cell growth and stemness properties were increased in PLC5 cells with long-term LPS exposure. (A) Cell viability and (B) clonogenic ability of PLC5 cells under long-term LPS exposure. PLC5 cells with or without LPS exposure for 20 passages were placed on 96-well plates for 4 days or on 6-well plates for 14 days. Cell viability was measured with MTT assays. Clonogenic ability was determined by colony formation, which was visualized by crystal violet. The relative viability and clonogenic ability of PLC5 cells without LPS treatment were set as 100 percent. Results are mean ± SD for three separate experiments. (C) Non-adherent sphere formation of PLC5 cells under long-term LPS exposure. PLC5 cells with or without LPS exposure for 20 passages were plated on 96-well plates and incubated for 4 days. Bright-field images are shown. Scale bar = 100 μm. Number of large spheres generated from 1 × 103 PLC5 cells with LPS exposure for indicated passages was quantified. Results are mean ± SD for three separate experiments. (D) The levels of OCT4 and NANOG mRNA in spheres from PLC5 cells with or without LPS exposure. The mRNA levels were detected with qPCR. (E) Effect of long-term LPS exposure on HCC cell growth in vivo. SCID mice were inoculated with 5 × 106 PLC5 cells with or without LPS exposure for 20 passages s.c. in the right flank (n = 6 for each group). Caliper measurements were performed to estimate the tumor volume (mean ± SD mm3). (* p < 0.05, ** p < 0.01).
3. Discussion
The inflammatory response is an important defense mechanism that impacts every single step of tumorigenesis [4]. It is known that chronic inflammatory response is a risk factor for liver cancer progression. TLR4 plays important roles in innate and inflammatory responses and is a crucial receptor to sense PAMPs and DAMPs. As the first line of defense against gut-derived antigens, emerging evidence has shown that activation of TLR4 signaling in liver contributed to inflammation- and injury-induced HCC promotion [33]. Based on our results from human HCC samples, the expression of TLR4 is positively correlated with IL-6 or CCL2 level, indicating that the expression of TLR4 is associated with inflammatory effectors. In line with published results, we showed that the activation of TLR4 signaling in HCC provides growth advantages by promoting cytokine expression and proliferation through the Akt and STAT3 pathways.
Signaling pathways that mediate the protumorigenic effects of inflammation are often subject to a feed-forward loop [2]. A recent study has shown that TLR4 activation promotes HCC progression through a COX-2/prostaglandin E2 (PGE2)/STAT3 positive feedback loop [34]. This study focused on investigating the mechanism that regulates TLR4 expression level in HCC in response to chronic inflammation. With our results, the activation of TLR4 triggers NF-κB-mediated inflammatory response, while LIN28 and let-7 miRNA are indicated in tumor development and are associated with inflammation. A positive correlation was found between TLR4 and LIN28A, while a negative correlation was demonstrated between let-7g miRNA and TLR4 or LIN28A. To mimic chronic inflammation with persistent TLR4 activation, we cultured PLC5 cells in low-dose LPS-containing medium; in this way, we demonstrated that mRNA and protein levels of TLR4 and LIN28A were elevated. The increase in LIN28A led to a decrease in let-7g miRNA. Furthermore, the 3′UTR of TLR4 mRNA was shown to be the target of let-7g miRNA, suggesting that the inhibition of let-7g miRNA was able to increase TLR4 mRNA.
The LIN28/let-7 axis has previously been highlighted in cancer progression [35]. It can be found in many cancers (e.g., breast cancer, lung cancer, or prostate cancer) that LIN28-positive tumors are more aggressive, and the inhibition of let-7 miRNAs promotes the development of cancer [24]. The LIN28/let-7 molecular switch has emerged as a central regulator of liver diseases, including liver injury by viral infection, hepatitis, cirrhosis, and HCC [25]. Increased LIN28 and decreased let-7 expression in HCC are pathologically associated with poor prognosis [36,37]. At present, 13 members of the let-7 family have been identified in humans, including let-7a-1, let-7a-2, let-7a-3, let-7b, let-7c, let-7d, let-7e, let-7f-1, let-7f-2, let-7g, let-7i, miR-98, and miR-202 [38]. Oncogenic signaling pathways, such as c-Myc, Ras, high-mobility group A (HMGA), and signal transducer and activator of transcription 3 (STAT3), which are critical in tumorigenesis, proliferation and invasion, are targeted by let-7 and, thus, let-7 is associated with stem cell differentiation and tumor suppression [39,40,41,42].
In this study, we demonstrated that HCC with long-term LPS exposure leads to increased cell proliferation and stemness. It has been reported previously that LPS could also affect tumor immunity. In addition, it has been observed that LPS leads to the differentiation of hepatic progenitors into myofibroblasts and increases the production of IL6 and TNF [43]. LPS is a trigger for macrophage-mediated inflammation, and LPS-mediated TLR4 activation induces pro-inflammatory cytokines and interferon (IFN)/IFN inducible genes [44,45].
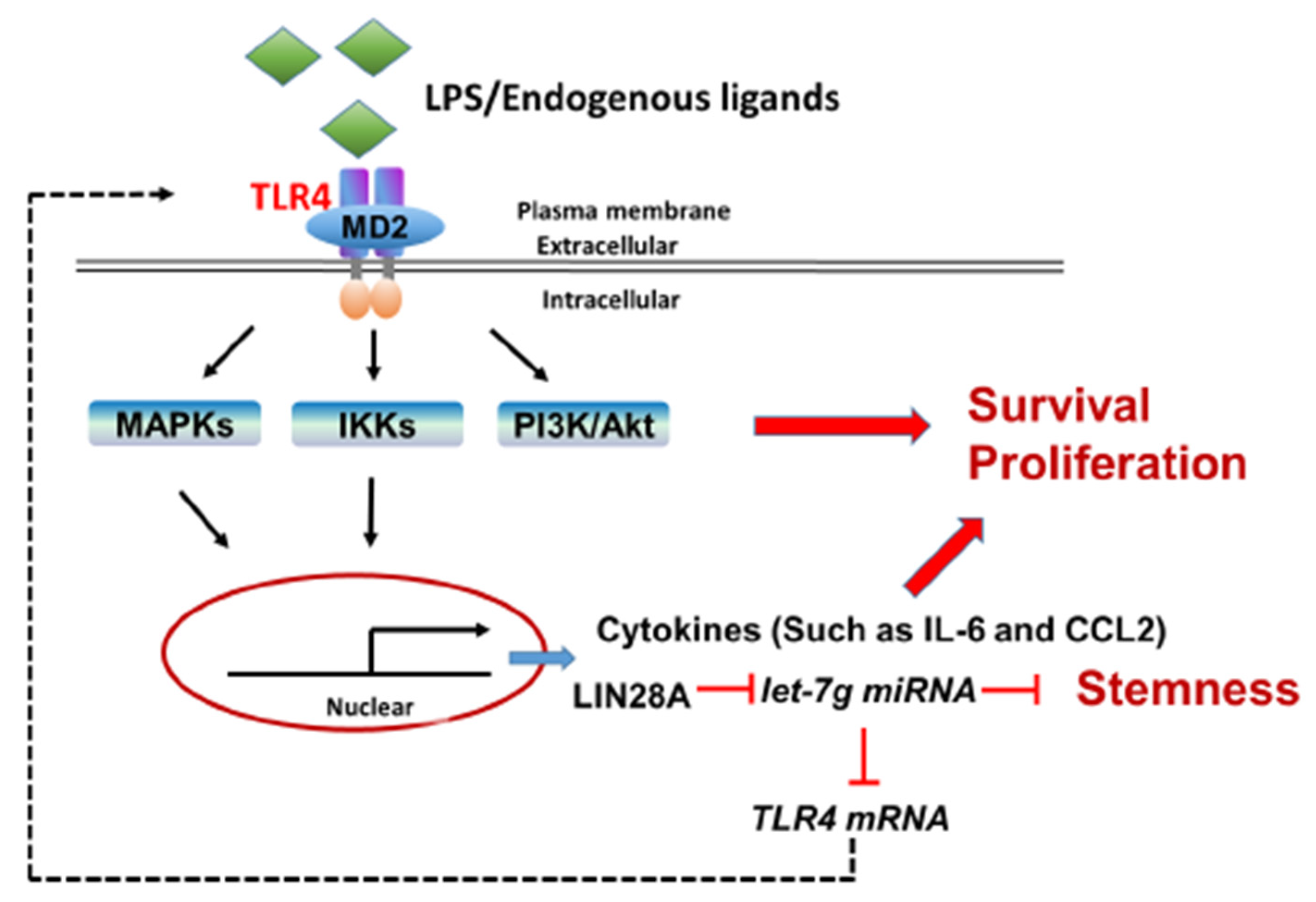
In addition to the chronic inflammatory microenvironment, the genes of interest in this study, including TLR4, LIN28A and let-7g miRNA, have been demonstrated to be involved in HCC tumorigenesis in several studies in the literature. However, this study elucidates the potential molecular mechanisms that modulate TLR4 expression in HCC through a positive feedback loop by the TLR4 and LIN28A/let-7g pathways in response to persistent TLR4 activation. Importantly, HCC cells with elevated TLR4 expression are sensitive to TLR4-mediated tumor promotion through the activation of Akt, NF-κB and STAT3 signaling and the downregulation of let-7 miRNA (Figure 8). These findings help to shape new concepts for the regulation of TLR4 expression in HCC through chronic inflammation and provide new insights for HCC clinical treatment. Furthermore, the genotype and phenotype of human HCC are variable and heterogenous; thus, it would be of interest to further investigate the clinicopathological features of HCC patients with elevated TLR4 in the future.
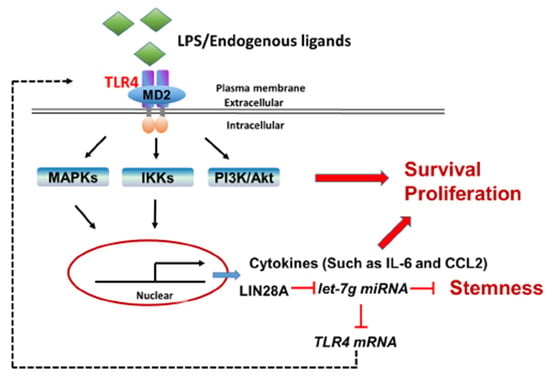
Figure 8.
Proposed model for the positive feedback loop of TLR4-mediated survival signaling in liver cancer.
4. Materials and Methods
4.1. Reagents and Antibodies
The following antibodies were used: anti-TLR4 (sc-10741), anti-JNK (sc-7345), anti-IκBα (sc-371), anti-α-tubulin (sc-23948) and anti-α-actin (sc-69879) from Santa Cruz (Dallas, TX, USA); HRP-conjugated anti-rabbit light chain (MAB201P) and HRP-conjugated anti-mouse light chain (AP200P) from EMD Millipore (Burlington, MA, USA); and anti-Akt (#9272), anti-LIN28A (#3978), anti-phospho-Akt (Thr308) (#2965), and anti-phospho-SAPK/JNK (Thr183/Tyr185) (#9251) from Cell Signaling (Danvers, MA, USA).
4.2. Study Subjects
The tumor tissues were collected from Stage I or II (based on The American Joint Committee on Cancer (AJCC) TNM system), HBV- and HCV-negative male patients that had not received chemotherapy or radiotherapy in the Taiwan Liver Cancer Network (TLCN). The HCC samples were used to clinically examine the expression of TLR4, IL6, CCL2, LIN28A and LIN28B mRNA and let-7b, let-7g and let-7i miRNA.
4.3. Cell Culture and Treatment
Human HEK293T and human PLC/PRF5 (PLC5) cells were obtained from the Bioresource Collection and Research Center (BCRC, Hsinchu, Taiwan). Human Huh7 cells were obtained from the Japanese Collection of Research Bioresources (JCRB, Tokyo, Japan). The cells were grown in Dulbecco’s modified Eagle medium (DMEM) containing 10% fetal bovine serum (FBS), 1 mM sodium pyruvate, and 1% (v/v) penicillin-streptomycin solution. DMEM, sodium pyruvate and penicillin–streptomycin solution were obtained from Biological Industries (Kibbutz Beit Haemek, Israel), and FBS was purchased from Invitrogen (Waltham, MA, USA).
4.4. Plasmids
The lentivirus-based pLKO.1-shTLR4 (TRCN0000056894 and TRCN0000056897) for human TLR4 shRNA-expression, pLKO.1-shLuc (TRCN0000072243) for luciferase shRNA-expression, pLKO-AS1010 for expressing indicated microRNA, and the packaging plasmids, pCMVΔ8.91 and pMD.G, were obtained from the National RNAi Core Facility (Institute of Molecular Biology/Genomics Research Center, Academia Sinica, Taipei, Taiwan). pRL-TK-Renilla luciferase and pGL3-control vector, used for the luciferase reporter assay, were from Promega (Madison, WI, USA). All constructs were confirmed by DNA sequencing.
4.5. Real-Time Quantitative Polymerase Chain Reaction
RNA samples were isolated with TRIzol (Invitrogen), and reverse transcribed to synthesize cDNA with 100 nM of each of the RT oligonucleotides (for RT of miRNA) or 10 µM of random hexamer (for RT of mRNA) by the iScript cDNA or iScriptTM select cDNA synthesis kit (Bio-Rad, Hercules, CA, USA). Target mRNA was quantified by real-time quantitative polymerase chain reaction (RT-qPCR) with the StepOnePlus system (Applied Biosystems, Waltham, MA, USA). Primer sequences are available in Supplementary Table S1. All values were normalized to the level of human GAPDH or mouse Cph mRNA expression.
4.6. Cell Viability Assay
The cell viability was determined using the MTT assay. Cells were seeded onto a 96-well plate at 5 × 103 cells per well. Twenty-four hours later, the culture medium was replaced with LPS for the indicated concentration. After incubating for 4 days, 30 μL of MTT solution (4 mg MTT/mL PBS) was added into each well and incubated for 4 h. The medium was removed, and Formazan crystals formed by the cells were dissolved using 200 μL of DMSO. The absorbance was read at 560 nm using 630 nm as the reference wavelength on a Multiwell plate reader.
4.7. Colony Formation Assay
Huh7 or PLC5 cells were seeded onto a 6-well plate at 1 × 103 cells per well. After incubating for two weeks, the colonies formed were fixed in 3.7% formaldehyde solution and visualized by staining with 0.2% crystal violet. The number of colonies was counted under the microscope. Data were obtained from 3 independent experiments.4.8. Lentivirus Production and Infection
HEK293T cells were transfected with the lentivirus-based constructs along with the packaging plasmids, pCMVΔ 8.91 and pMD.G, using T-pro NTRII (T-Pro Biotechnology, New Taipei City Taiwan). Virus-containing medium was collected at 48 and 72 h post-transfection. Cells were infected with lentivirus-containing medium at a multiplicity of infection (MOI) of 10 to 25 in the presence of 10 μg/mL polybrene (Sigma-Aldrich, St. Louis, MO, USA). After 24 h, the virus-containing medium was replaced with selection medium containing 5 μg/mL puromycin (EMD Millipore). After cell growth was stable, cells were used in subsequent experiments.
4.8. Mouse Mammary Tumor Model and Xenograft Tumor Formation
For the DEN-induced HCC model, three-week-old male C57/BL6 mice were injected intraperitoneally (i.p.) with 5 mg/kg of DEN (Sigma-Aldrich). After 8 months on normal chow, mice were sacrificed, and HCC cells were isolated as described by He et al. [46]. For the xenograft tumor model, six-week-old male SCID mice were obtained from the National Laboratory Animal Center (Taipei, Taiwan). Each mouse was inoculated s.c. in the right dorsal flank with indicated cancer cells suspended in 50 μL PBS and an equal volume of Matrigel (BD Biosciences, San Jose, CA, USA). Tumors were measured with Vernier calipers. Tumor volumes were calculated with the following formula: width2 × length × 0.52. Whenever tumor size reached 2000 mm3 before the end of treatment, mice were sacrificed based on the Animal Care and Use Guidelines.
4.9. Immunoblotting
Cells were lysed in ice-cold RIPA lysis buffer containing 50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1% NP40, 0.5% deoxycholate, 0.1% SDS, 1 mM PMSF, 1 mM Na3VO4 and 1 mM NaF. After centrifugation, the proteins in each cell extract were separated by SDS-PAGE, electro-transferred to polyvinylidene difluoride (PVDF) membrane and analyzed by immunoblotting. The membranes were stained with ponceau S to confirm the efficiency and consistency of the protein, blocked with 5% nonfat dry milk in TBST buffer, and then probed with the indicated antibodies. The blots were visualized using a chemiluminescence reagent (Thermo, Rockford, IL, USA) and exposure of an X-ray film (Fujifilm, Taipei, Taiwan). Quantification was carried out with Image J 1.53.
4.10. Regrowth Protocol for PLC5 Cells with or without LPS Exposure
PLC5 cells were seeded in a 6 cm dish at a density of 1 × 106 cells per 5 mL of culture medium with or without 10 ng/mL LPS. After 3 days, the cells were trypsinized, counted and subcultured with the indicated density for indicated passages.
4.11. Sphere-Formation Assay
PLC5 cells (1 × 105) were seeded in a 10 cm dish and cultured in tumor sphere medium consisting of serum-free DMEM/F12 medium (GIBCO, Waltham, MA, USA), N-2 supplement (GIBCO), 10 ng/mL human recombinant bFGF-basic (PeproTech, Rehovot, Israel), and 10 ng/mL EGF (PeproTech). The medium was changed every other day until tumor sphere formation was achieved in approximately 3 weeks [47].
4.12. Luciferase Reporter Assay
As previously described [48], HEK293T cells were seeded onto a 96-well plate at 1 × 104 cells per well, and co-transfected with 10 ng of luciferase reporter and 50 ng of miRNA-expressing construct using T-pro NTRII. After transfection for 24 h, luciferase activity was monitored using the Dual-Glo® luciferase assay system and a luminometer. Renilla luciferase activity was normalized against Firefly luciferase activities and presented as the percentage of inhibition.
4.13. Statistical Analysis
Correlation analysis was completed using the Pearson method with SPSS statistical software (IBM SPSS Statistics 20.0, IBM, Armonk, NY, USA). All data were reported as mean ± SD. The comparison of means was performed with multiple t-tests using GraphPad Prism version 6.01 (GraphPad Software, San Diego, CA, USA). The tumor volume data satisfied the assumptions of normality and homogeneity required for parametric analysis and, thus, group means at the indicated days were compared with a one-way analysis of variance followed by Fisher’s least significant difference method for multiple comparisons using GraphPad Prism (version 6.01). A p value of less than 0.05 (* p < 0.05, ** p < 0.01) was considered statistically significant.
Supplementary Materials
The following are available online at: https://www.mdpi.com/article/10.3390/ijms23158419/s1.
Author Contributions
Conceptualization, P.-H.T.; methodology, C.Y. and I.-T.C.; software, C.Y.; validation, I.-T.C., A.-C.C. and Y.-C.C.; formal analysis, I.-T.C. and C.Y.; investigation, A.-C.C., Y.-T.L., C.Y. and Y.-C.C.; resources, P.-H.T. and C.-F.C.; writing—original draft preparation, I.-T.C.; writing—review and editing, P.-H.T. and C.-F.C.; supervision, P.-H.T. and C.-F.C.; funding acquisition, C.-F.C. and P.-H.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants from National Health Research Institutes, Taiwan (NHRI-EX103-10111BC) to P.-H.T., and National Yang-Ming University-Far Eastern Memorial Hospital Joint Research Program (NYMU-FEMH 111DN24, 110DN29; 109DN28) to P.-H.T. and C.-F.C. The funding institutions of this study had no further role in the study design, the collection, analysis, and interpretation of data, the writing of this paper, or the decision to submit it for publication.
Institutional Review Board Statement
The use of human samples and animals was in accordance with national approved law (Human Subjects Research Act and Animal Protection Act) and institutionally approved guidelines. Archived human HCC tumor samples were obtained from the Taiwan Liver Cancer Network (TLCN) and approved by the Institutional Review Board of National Yang-Ming University (YM103007E). All animal studies were performed according to the animal study protocol approved by the Institutional Animal Care and Use Committee (IACUC) of National Yang-Ming University (IACUC #1000503r) and the recommendation of “A Guidebook for Care and Use of Laboratory Animals” (Third Edition). All efforts were made to minimize animal discomfort and suffering.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding authors.
Acknowledgments
We would like to thank the Taiwan Liver Cancer Network (TLCN) for providing the liver tissue samples and related clinical data (all are anonymous) for our research. TLCN is supported by grants from Ministry of Science and Technology and National Health Research Institute, Taiwan. RNAi reagents were obtained from the National RNAi Core Facility, Academia Sinica, which is supported by the National Core Facility Program for Biotechnology Grants of the NSC (NSC 100-2319-B-001-002). We are grateful to Jeng-Fan Lo for their helpful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sengupta, R.; Honey, K. AACR Cancer Progress Report 2019: Transforming Lives Through Innovative Cancer Science. Clin. Cancer Res. 2019, 25, 5431. [Google Scholar] [CrossRef] [Green Version]
- Anstee, Q.M.; Reeves, H.L.; Kotsiliti, E.; Govaere, O.; Heikenwalder, M. From NASH to HCC: Current concepts and future challenges. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Marengo, A.; Jouness, R.I.; Bugianesi, E. Progression and Natural History of Nonalcoholic Fatty Liver Disease in Adults. Clin. Liver Dis. 2016, 20, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, Q.; Wang, L.; Chen, W.; Li, N.; Cao, X. TLR4 signaling promotes immune escape of human lung cancer cells by inducing immunosuppressive cytokines and apoptosis resistance. Mol. Immunol. 2007, 44, 2850–2859. [Google Scholar] [CrossRef] [PubMed]
- Eiro, N.; Altadill, A.; Juarez, L.M.; Rodriguez, M.; Gonzalez, L.O.; Atienza, S.; Bermudez, S.; Fernandez-Garcia, B.; Fresno-Forcelledo, M.F.; Rodrigo, L.; et al. Toll-like receptors 3, 4 and 9 in hepatocellular carcinoma: Relationship with clinicopathological characteristics and prognosis. Hepatol. Res. 2014, 44, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, Z.; Ye, J.; Zhang, X.; Wu, H.; Peng, J.; Song, W.; Chen, C.; Cai, S.; He, Y.; et al. Uncontrolled inflammation induced by AEG-1 promotes gastric cancer and poor prognosis. Cancer Res. 2014, 74, 5541–5552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.J.; Wu, H.S.; Wang, L.; Tian, Y.; Zhang, J.H.; Wu, H.L. Expression and significance of TLR4 and HIF-1alpha in pancreatic ductal adenocarcinoma. World J. Gastroenterol. 2010, 16, 2881–2888. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.G.; Alvero, A.B.; Chen, R.; Silasi, D.A.; Abrahams, V.M.; Chan, S.; Visintin, I.; Rutherford, T.; Mor, G. TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer. Cancer Res. 2006, 66, 3859–3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oblak, A.; Jerala, R. Toll-like receptor 4 activation in cancer progression and therapy. Clin. Dev. Immunol. 2011, 2011, 609579. [Google Scholar] [CrossRef] [Green Version]
- Pradere, J.P.; Dapito, D.H.; Schwabe, R.F. The Yin and Yang of Toll-like receptors in cancer. Oncogene 2014, 33, 3485–3495. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Shi, X. MicroRNAs in the regulation of TLR and RIG-I pathways. Cell Mol. Immunol. 2013, 10, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Benakanakere, M.R.; Li, Q.; Eskan, M.A.; Singh, A.V.; Zhao, J.; Galicia, J.C.; Stathopoulou, P.; Knudsen, T.B.; Kinane, D.F. Modulation of TLR2 protein expression by miR-105 in human oral keratinocytes. J. Biol. Chem. 2009, 284, 23107–23115. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Chen, J.; Zhou, L.; Chen, W.; Ding, G.; Cao, L. Pancreatic cancer derived exosomes regulate the expression of TLR4 in dendritic cells via miR-203. Cell Immunol. 2014, 292, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Splinter, P.L.; O’Hara, S.P.; LaRusso, N.F. A cellular micro-RNA, let-7i, regulates Toll-like receptor 4 expression and contributes to cholangiocyte immune responses against Cryptosporidium parvum infection. J. Biol. Chem. 2007, 282, 28929–28938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Androulidaki, A.; Iliopoulos, D.; Arranz, A.; Doxaki, C.; Schworer, S.; Zacharioudaki, V.; Margioris, A.N.; Tsichlis, P.N.; Tsatsanis, C. The kinase Akt1 controls macrophage response to lipopolysaccharide by regulating microRNAs. Immunity 2009, 31, 220–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, G.G.; Wang, W.H.; Dai, Y.; Wang, S.J.; Chu, Y.X.; Li, J. Let-7b is involved in the inflammation and immune responses associated with Helicobacter pylori infection by targeting Toll-like receptor 4. PLoS ONE 2013, 8, e56709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viswanathan, S.R.; Daley, G.Q.; Gregory, R.I. Selective blockade of microRNA processing by Lin28. Science 2008, 320, 97–100. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.A.; Thomson, J.M.; Hammond, S.M. Lin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processing. RNA 2008, 14, 1539–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, I.; Joo, C.; Cho, J.; Ha, M.; Han, J.; Kim, V.N. Lin28 mediates the terminal uridylation of let-7 precursor MicroRNA. Mol. Cell 2008, 32, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Piskounova, E.; Polytarchou, C.; Thornton, J.E.; LaPierre, R.J.; Pothoulakis, C.; Hagan, J.P.; Iliopoulos, D.; Gregory, R.I. Lin28A and Lin28B inhibit let-7 microRNA biogenesis by distinct mechanisms. Cell 2011, 147, 1066–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/let-7 Pathway in Cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniel, K.; Hall, C.; Sato, K.; Lairmore, T.; Marzioni, M.; Glaser, S.; Meng, F.; Alpini, G. Lin28 and let-7: Roles and regulation in liver diseases. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G757–G765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. An epigenetic switch involving NF-kappaB, Lin28, Let-7 MicroRNA, and IL6 links inflammation to cell transformation. Cell 2009, 139, 693–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, Y.; Zhang, D.; Jiang, S.; Li, A.; Guo, A.; Wu, X.; Xia, X.; Cheng, H.; Tao, T.; Gu, X. LIN28 expression in rat spinal cord after injury. Neurochem. Res. 2014, 39, 862–874. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Chen, C.; Shi, M.; Wang, F.; Chen, X.; Diao, D.; Hu, M.; Yu, M.; Qian, L.; Guo, N. Stat3-coordinated Lin-28-let-7-HMGA2 and miR-200-ZEB1 circuits initiate and maintain oncostatin M-driven epithelial-mesenchymal transition. Oncogene 2013, 32, 5272–5282. [Google Scholar] [CrossRef] [PubMed]
- Dapito, D.H.; Mencin, A.; Gwak, G.Y.; Pradere, J.P.; Jang, M.K.; Mederacke, I.; Caviglia, J.M.; Khiabanian, H.; Adeyemi, A.; Bataller, R.; et al. Promotion of hepatocellular carcinoma by the intestinal microbiota and TLR4. Cancer Cell 2012, 21, 504–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Martin, R.C.; Shi, X.; Pandit, H.; Yu, Y.; Liu, X.; Guo, W.; Tan, M.; Bai, O.; Meng, X.; et al. Lack of FGF21 promotes NASH-HCC transition via hepatocyte-TLR4-IL-17A signaling. Theranostics 2020, 10, 9923–9936. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Jing, Y.Y.; Yu, G.F.; Han, Z.P.; Yu, D.D.; Fan, Q.M.; Ye, F.; Li, R.; Gao, L.; Zhao, Q.D.; et al. Toll like receptor 4 facilitates invasion and migration as a cancer stem cell marker in hepatocellular carcinoma. Cancer Lett. 2015, 358, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.Y.; Han, Z.P.; Sun, K.; Zhang, S.S.; Hou, J.; Liu, Y.; Li, R.; Gao, L.; Zhao, X.; Zhao, Q.D.; et al. Toll-like receptor 4 signaling promotes epithelial-mesenchymal transition in human hepatocellular carcinoma induced by lipopolysaccharide. BMC Med. 2012, 10, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiziltas, S. Toll-like receptors in pathophysiology of liver diseases. World J. Hepatol. 2016, 8, 1354–1369. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Wang, G.; Zhao, H.; Zhang, Y.; Han, Q.; Zhang, C.; Tian, Z.; Zhang, J. TLR4 signaling promotes a COX-2/PGE2/STAT3 positive feedback loop in hepatocellular carcinoma (HCC) cells. Oncoimmunology 2016, 5, e1074376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wang, G.; Hao, D.; Liu, X.; Wang, D.; Ning, N.; Li, X. Aberrant regulation of the LIN28A/LIN28B and let-7 loop in human malignant tumors and its effects on the hallmarks of cancer. Mol. Cancer 2015, 14, 125. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, S.R.; Powers, J.T.; Einhorn, W.; Hoshida, Y.; Ng, T.L.; Toffanin, S.; O’Sullivan, M.; Lu, J.; Phillips, L.A.; Lockhart, V.L.; et al. Lin28 promotes transformation and is associated with advanced human malignancies. Nat. Genet. 2009, 41, 843–848. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Zhao, L.; Budhu, A.; Forgues, M.; Jia, H.L.; Qin, L.X.; Ye, Q.H.; Yu, J.; Shi, X.; Tang, Z.Y.; et al. Let-7g targets collagen type I alpha2 and inhibits cell migration in hepatocellular carcinoma. J. Hepatol. 2010, 52, 690–697. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cao, L.; Wang, Y.; Wang, X.; Liu, N.; You, Y. Regulation of let-7 and its target oncogenes (Review). Oncol. Lett. 2012, 3, 955–960. [Google Scholar] [CrossRef]
- Johnson, S.M.; Grosshans, H.; Shingara, J.; Byrom, M.; Jarvis, R.; Cheng, A.; Labourier, E.; Reinert, K.L.; Brown, D.; Slack, F.J. RAS is regulated by the let-7 microRNA family. Cell 2005, 120, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Sampson, V.B.; Rong, N.H.; Han, J.; Yang, Q.; Aris, V.; Soteropoulos, P.; Petrelli, N.J.; Dunn, S.P.; Krueger, L.J. MicroRNA let-7a down-regulates MYC and reverts MYC-induced growth in Burkitt lymphoma cells. Cancer Res. 2007, 67, 9762–9770. [Google Scholar] [CrossRef] [Green Version]
- Mayr, C.; Hemann, M.T.; Bartel, D.P. Disrupting the pairing between let-7 and Hmga2 enhances oncogenic transformation. Science 2007, 315, 1576–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lu, Y.; Toh, S.T.; Sung, W.K.; Tan, P.; Chow, P.; Chung, A.Y.; Jooi, L.L.; Lee, C.G. Lethal-7 is down-regulated by the hepatitis B virus x protein and targets signal transducer and activator of transcription 3. J. Hepatol. 2010, 53, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Jing, Y.Y.; Gao, L.; Li, R.; Yang, X.; Pan, X.R.; Yang, Y.; Meng, Y.; Hou, X.J.; Zhao, Q.D.; et al. Lipopolysaccharide induces the differentiation of hepatic progenitor cells into myofibroblasts constitutes the hepatocarcinogenesis-associated microenvironment. Cell Death Differ. 2020, 27, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Aravalli, R.N. Role of innate immunity in the development of hepatocellular carcinoma. World J. Gastroenterol. 2013, 19, 7500–7514. [Google Scholar] [CrossRef]
- He, G.; Yu, G.Y.; Temkin, V.; Ogata, H.; Kuntzen, C.; Sakurai, T.; Sieghart, W.; Peck-Radosavljevic, M.; Leffert, H.L.; Karin, M. Hepatocyte IKKbeta/NF-kappaB inhibits tumor promotion and progression by preventing oxidative stress-driven STAT3 activation. Cancer Cell 2010, 17, 286–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.W.; Chen, Y.S.; Chou, S.H.; Han, C.L.; Chen, Y.J.; Yang, C.C.; Huang, C.Y.; Lo, J.F. Distinct subpopulations of head and neck cancer cells with different levels of intracellular reactive oxygen species exhibit diverse stemness, proliferation, and chemosensitivity. Cancer Res. 2014, 74, 6291–6305. [Google Scholar] [CrossRef] [Green Version]
- Qiu, C.; Ma, Y.; Wang, J.; Peng, S.; Huang, Y. Lin28-mediated post-transcriptional regulation of Oct4 expression in human embryonic stem cells. Nucleic Acids Res. 2010, 38, 1240–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).