
Molecular Mimicry of the Rheumatoid Arthritis-Related Immunodominant T-Cell Epitope within Type II Collagen (CII260-270) by the Bacterial L-Asparaginase






Abstract
:1. Introduction
2. Results
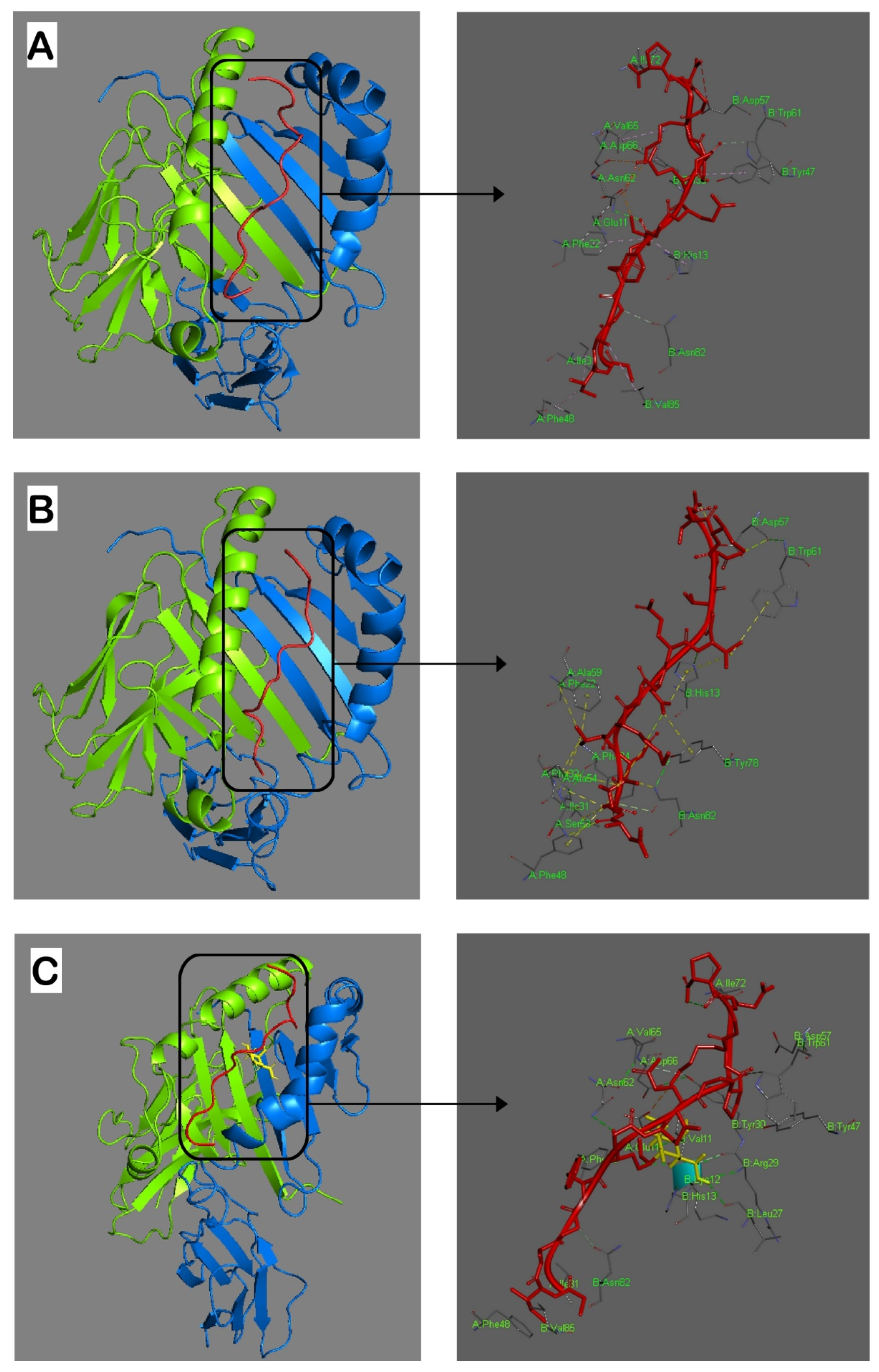
2.1. MHC Class II (MHCII) Binding Predictions and Molecular Docking
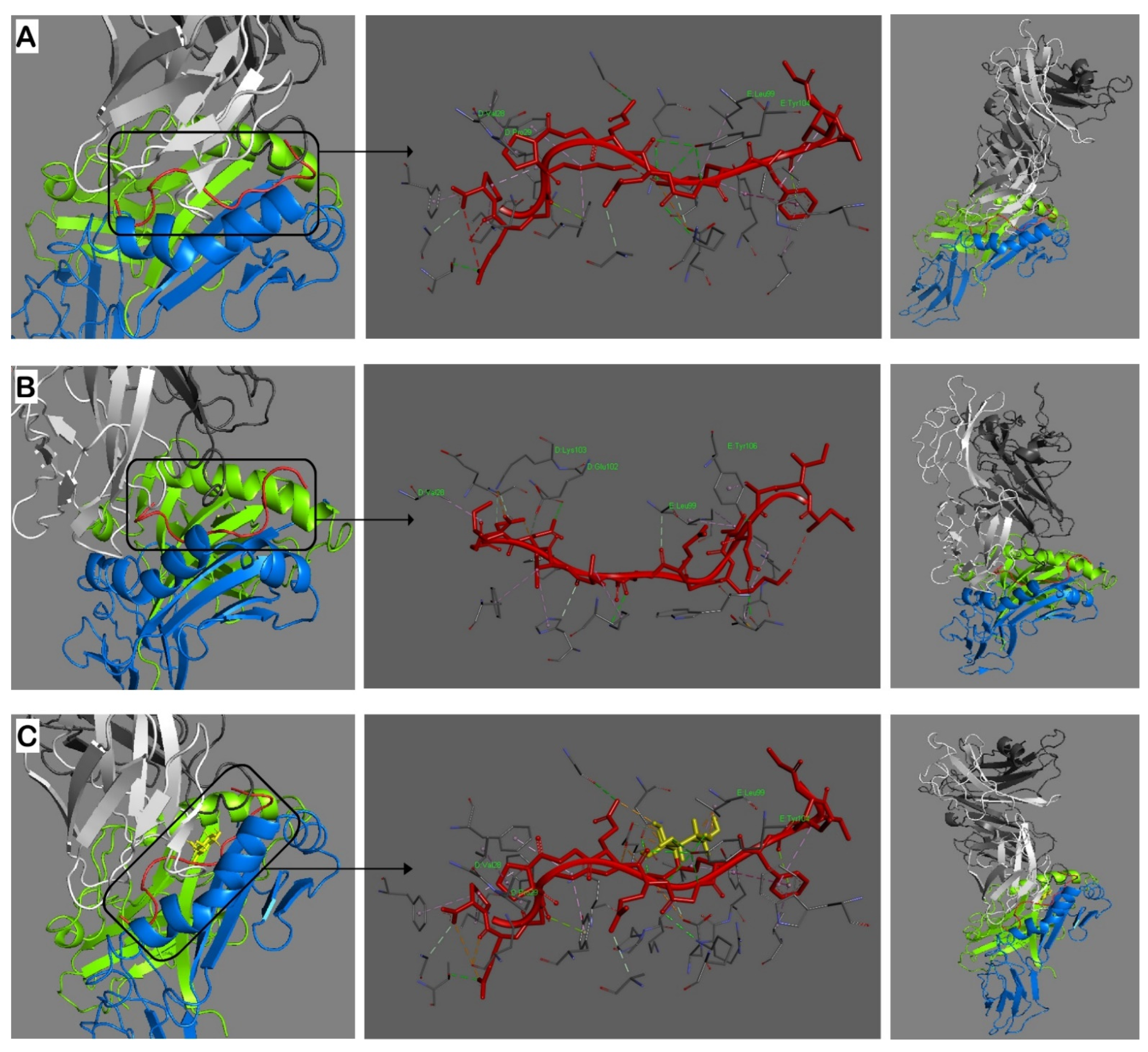
2.2. Modeling of Interactions in the Trimolecular Complex HLA-DRB1*04:01—Peptide—TCR
2.3. Baseline Demographic and Disease Characteristics of Selected Early RA (ERA) Patients and Healthy Subjects
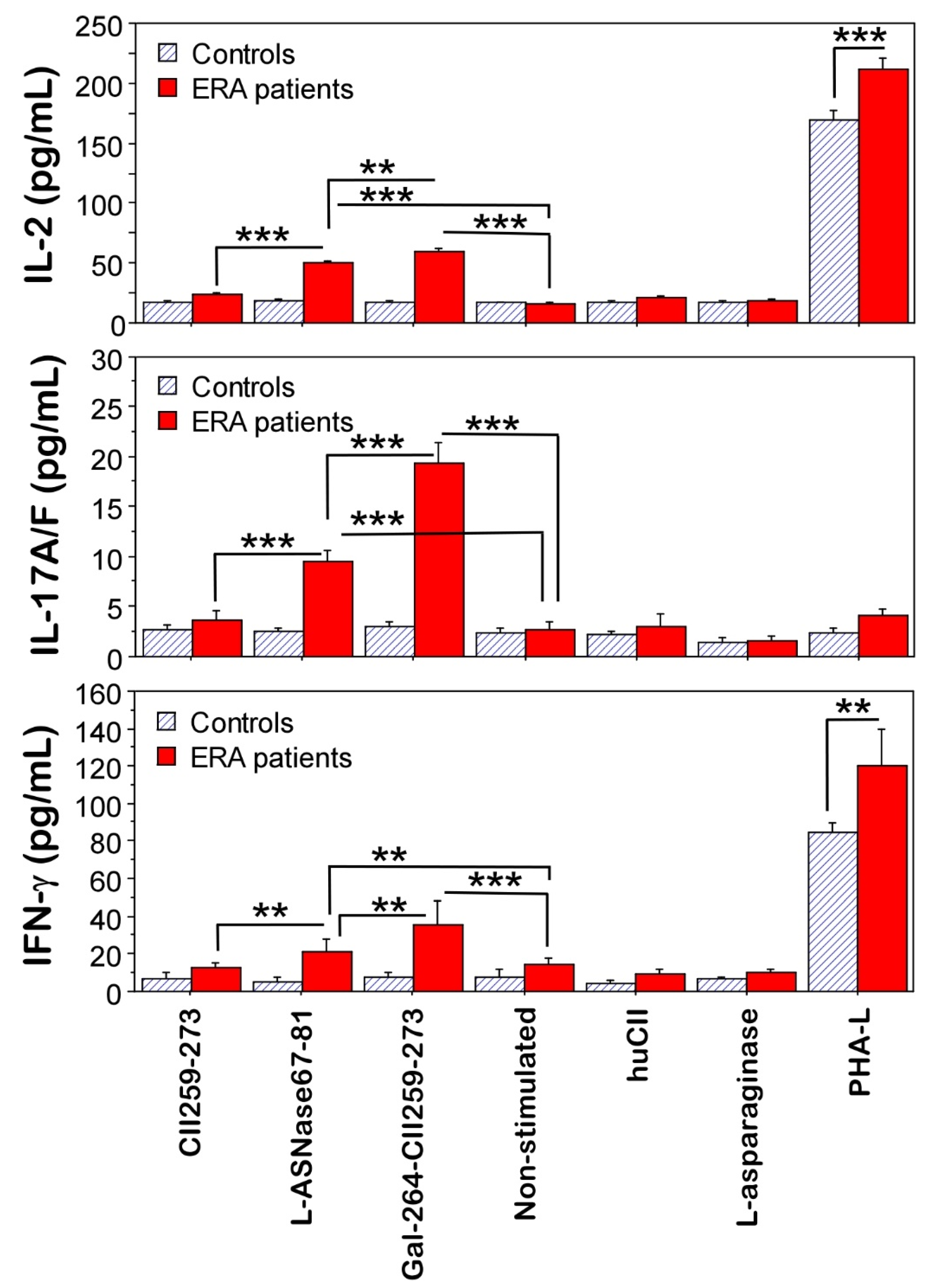
2.4. Functional Validation of T-Cell Reactivity
3. Discussion
4. Materials and Methods
4.1. Antigens
4.2. MHCII Binding Predictions and Molecular Docking
4.3. Patients, Human Leucocyte Antigen (HLA), Genotyping, and Measurement of ACPA Levels and Rheumatoid Factor (RF)
4.4. Functional Validation of T-Cell Reactivity
4.4.1. Cell Isolation and Stimulation
4.4.2. Fluorescence-Activated Cell Sorting (FACS) Analysis
4.4.3. Cytokine Measurements in Supernatants by ELISA
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chriswell, M.E.; Kuhn, K.A. Microbiota-mediated mucosal inflammation in arthritis. Best Pract. Res. Clin. Rheumatol. 2019, 33, 101492. [Google Scholar] [CrossRef]
- Cusick, M.F.; Libbey, J.E.; Fujinami, R.S. Molecular Mimicry as a Mechanism of Autoimmune Disease. Clin. Rev. Allergy Immunol. 2012, 42, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, A.M.; Miller, S.D. The role of infections in autoimmune disease. Clin. Exp. Immunol. 2009, 155, 1–15. [Google Scholar] [CrossRef]
- Rojas, M.; Restrepo-Jiménez, P.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Leung, P.S.C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.-M. Molecular mimicry and autoimmunity. J. Autoimmun. 2018, 95, 100–123. [Google Scholar] [CrossRef] [PubMed]
- Yurkovetskiy, L.A.; Pickard, J.M.; Chervonsky, A.V. Microbiota and Autoimmunity: Exploring New Avenues. Cell Host Microbe 2015, 17, 548–552. [Google Scholar] [CrossRef]
- Repac, J.; Mandić, M.; Lunić, T.; Božić, B.; Božić Nedeljković, B. Mining the capacity of human-associated microorganisms to trigger rheumatoid arthritis—A systematic immunoinformatics analysis of T cell epitopes. PLoS ONE 2021, 16, e0253918. [Google Scholar] [CrossRef]
- Chemin, K.; Gerstner, C.; Malmström, V. Effector Functions of CD4+ T Cells at the Site of Local Autoimmune Inflammation—Lessons From Rheumatoid Arthritis. Front. Immunol. 2019, 10, 353. [Google Scholar] [CrossRef]
- Yang, P.; Qian, F.Y.; Zhang, M.F.; Xu, A.L.; Wang, X.; Jiang, B.P.; Zhou, L.L. Th17 cell pathogenicity and plasticity in rheumatoid arthritis. J. Leukoc. Biol. 2019, 106, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, M.A.; Redwan, E.M. Interplay of Microbiota and Citrullination in the Immunopathogenesis of Rheumatoid Arthritis. Probiotics Antimicrob. Proteins 2021, 14, 99–113. [Google Scholar] [CrossRef]
- Sherina, N.; de Vries, C.; Kharlamova, N.; Sippl, N.; Jiang, X.; Brynedal, B.; Kindstedt, E.; Hansson, M.; Mathsson-Alm, L.; Israelsson, L.; et al. Antibodies to a Citrullinated Porphyromonas gingivalis Epitope Are Increased in Early Rheumatoid Arthritis, and Can Be Produced by Gingival Tissue B Cells: Implications for a Bacterial Origin in RA Etiology. Front. Immunol. 2022, 13, 4822. [Google Scholar] [CrossRef]
- Kampstra, A.S.B.; Toes, R.E.M. HLA class II and rheumatoid arthritis: The bumpy road of revelation. Immunogenetics 2017, 69, 597–603. [Google Scholar] [CrossRef]
- Karami, J.; Aslani, S.; Jamshidi, A.; Garshasbi, M.; Mahmoudi, M. Genetic implications in the pathogenesis of rheumatoid arthritis; an updated review. Gene 2019, 702, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Pratt, A.G.; Isaacs, J.D. Genotyping in rheumatoid arthritis: A game changer in clinical management? Expert Rev. Clin. Immunol. 2015, 11, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Gregersen, P.K.; Silver, J.; Winchester, R.J. The shared epitope hypothesis. an approach to understanding the molecular genetics of susceptibility to rheumatoid arthritis. Arthritis Rheum. 1987, 30, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Larid, G.; Pancarte, M.; Offer, G.; Clavel, C.; Martin, M.; Pradel, V.; Auger, I.; Lafforgue, P.; Roudier, J.; Serre, G.; et al. In Rheumatoid Arthritis Patients, HLA-DRB1*04:01 and Rheumatoid Nodules Are Associated With ACPA to a Particular Fibrin Epitope. Front. Immunol. 2021, 12, 692041. [Google Scholar] [CrossRef]
- Bäcklund, J.; Carlsen, S.; Höger, T.; Holm, B.; Fugger, L.; Kihlberg, J.; Burkhardt, H.; Holmdahl, R. Predominant selection of T cells specific for the glycosylated collagen type II epitope (263–270) in humanized transgenic mice and in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2002, 99, 9960–9965. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Kim, W.-U.; Cho, M.-L.; Lee, S.K.; Youn, J.; Kim, S.-I.; Yoo, W.-H.; Park, J.-H.; Min, J.-K.; Lee, S.-H.; et al. Enhanced T cell proliferative response to type II collagen and synthetic peptide CII (255-274) in patients with rheumatoid arthritis. Arthritis Rheum. 1999, 42, 2085–2093. [Google Scholar] [CrossRef]
- Londei, M.; Savill, C.M.; Verhoef, A.; Brennan, F.; Leech, Z.A.; Duance, V.; Maini, R.N.; Feldmann, M. Persistence of collagen type II-specific T-cell clones in the synovial membrane of a patient with rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 1989, 86, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Snir, O.; Bäcklund, J.; Boström, J.; Andersson, I.; Kihlberg, J.; Buckner, J.H.; Klareskog, L.; Holmdahl, R.; Malmström, V. Multifunctional T cell reactivity with native and glycosylated type II collagen in rheumatoid arthritis. Arthritis Rheum. 2012, 64, 2482–2488. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Tsutsumi, A.; Sakamaki, T.; Sumida, T. T cell epitopes of type II collagen in HLA-DRB1*0101 or DRB1*0405-positive Japanese patients with rheumatoid arthritis. Int. J. Mol. Med. 2003, 11, 331–335. [Google Scholar] [CrossRef]
- Holmdahl, R.; Andersson, M.; Goldschmidt, T.J.; Gustafsson, K.E.; Jansson, L.; Mo, J.A. Type II Collagen Autoimmunity in Animals and Provocations Leading to Arthritis. Immunol. Rev. 1990, 118, 193–232. [Google Scholar] [CrossRef]
- Di Sante, G.; Tolusso, B.; Fedele, A.L.; Gremese, E.; Alivernini, S.; Nicolò, C.; Ria, F.; Ferraccioli, G. Collagen Specific T-Cell Repertoire and HLA-DR Alleles: Biomarkers of Active Refractory Rheumatoid Arthritis. EBioMedicine 2015, 2, 2037–2045. [Google Scholar] [CrossRef] [PubMed]
- Lubkowski, J.; Wlodawer, A. Structural and biochemical properties of L-asparaginase. FEBS J. 2021, 288, 4183–4209. [Google Scholar] [CrossRef] [PubMed]
- Pokrovskaya, M.V.; Pokrovsky, V.S.; Aleksandrova, S.S.; Sokolov, N.N.; Zhdanov, D.D. Molecular Analysis of L-Asparaginases for Clarification of the Mechanism of Action and Optimization of Pharmacological Functions. Pharmaceutics 2022, 14, 599. [Google Scholar] [CrossRef]
- Batool, T.; Makky, E.A.; Jalal, M.; Yusoff, M.M. A Comprehensive Review on l-Asparaginase and Its Applications. Appl. Biochem. Biotechnol. 2015, 178, 900–923. [Google Scholar] [CrossRef] [PubMed]
- Nunes, J.C.F.; Cristóvão, R.O.; Freire, M.G.; Santos-Ebinuma, V.C.; Faria, J.L.; Silva, C.G.; Tavares, A.P.M. Recent Strategies and Applications for l-Asparaginase Confinement. Molecules 2020, 25, 5827. [Google Scholar] [CrossRef]
- Batsalova, T.; Dzhambazov, B.; Merky, P.; Bäcklund, A.; Bäcklund, J. Breaking T cell tolerance against self-type II collagen in HLA-DR4 transgenic mice and development of autoimmune arthritis. Arthritis Rheum. 2010, 62, 1911–1920. [Google Scholar] [CrossRef]
- Berner, B. Increased expression of CD40 ligand (CD154) on CD4+ T cells as a marker of disease activity in rheumatoid arthritis. Ann. Rheum. Dis. 2000, 59, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Tung, C.-H.; Lu, M.-C.; Lai, N.-S.; Wu, S.-F. Tumor necrosis factor-α blockade treatment decreased CD154 (CD40-ligand) expression in rheumatoid arthritis. PLoS ONE 2017, 12, e0183726. [Google Scholar] [CrossRef]
- Andersson, E.C.; Hansen, B.E.; Jacobsen, H.; Madsen, L.S.; Andersen, C.B.; Engberg, J.; Rothbard, J.B.; McDevitt, G.S.; Malmström, V.; Holmdahl, R.; et al. Definition of MHC and T cell receptor contacts in the HLA-DR4restricted immunodominant epitope in type II collagen and characterization of collagen-induced arthritis in HLA-DR4 and human CD4 transgenic mice. Proc. Natl. Acad. Sci. USA 1998, 95, 7574–7579. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Weisse, S.; Xu, B.; Dobritzsch, D.; Viljanen, J.; Kihlberg, J.; Do, N.-N.; Schneider, N.; Lanig, H.; Holmdahl, R.; et al. Key interactions in the trimolecular complex consisting of the rheumatoid arthritis-associated DRB1*04:01 molecule, the major glycosylated collagen II peptide and the T-cell receptor. Ann. Rheum. Dis. 2022, 81, 480–489. [Google Scholar] [CrossRef]
- Rosloniec, E.F.; Whittington, K.B.; Zaller, D.M.; Kang, A.H. HLA-DR1 (DRB1*0101) and DR4 (DRB1*0401) Use the Same Anchor Residues for Binding an Immunodominant Peptide Derived from Human Type II Collagen. J. Immunol. 2002, 168, 253–259. [Google Scholar] [CrossRef]
- Liu, M.F.; Chao, S.C.; Wang, C.R.; Lei, H.Y. Expression of CD40 and CD40 ligand among cell populations within rheumatoid synovial compartment. Autoimmunity 2001, 34, 107–113. [Google Scholar] [CrossRef]
- Snir, O.; Rieck, M.; Gebe, J.A.; Yue, B.B.; Rawlings, C.A.; Nepom, G.; Malmström, V.; Buckner, J.H. Identification and functional characterization of T cells reactive to citrullinated vimentin in HLA-DRB1*0401-positive humanized mice and rheumatoid arthritis patients. Arthritis Rheum. 2011, 63, 2873–2883. [Google Scholar] [CrossRef]
- Liu, S.; Watanabe, S.; Shudou, M.; Kuno, M.; Miura, H.; Maeyama, K. Upregulation of store-operated Ca2+ entry in the naïve CD4+ T cells with aberrant cytokine releasing in active rheumatoid arthritis. Immunol. Cell Biol. 2014, 92, 752–760. [Google Scholar] [CrossRef]
- Hassan, G.S.; Salti, S.; Mourad, W. Novel Functions of Integrins as Receptors of CD154: Their Role in Inflammation and Apoptosis. Cells 2022, 11, 1747. [Google Scholar] [CrossRef]
- Spolski, R.; Leonard, W.J. Cytokine mediators of Th17 function. Eur. J. Immunol. 2009, 39, 658–661. [Google Scholar] [CrossRef]
- Dienz, O.; Eaton, S.M.; Bond, J.P.; Neveu, W.; Moquin, D.; Noubade, R.; Briso, E.M.; Charland, C.; Leonard, W.J.; Ciliberto, G.; et al. The induction of antibody production by IL-6 is indirectly mediated by IL-21 produced by CD4+ T cells. J. Exp. Med. 2009, 206, 69–78. [Google Scholar] [CrossRef]
- Wang, P.; Sidney, J.; Dow, C.; Mothé, B.; Sette, A.; Peters, B. A Systematic Assessment of MHC Class II Peptide Binding Predictions and Evaluation of a Consensus Approach. PLoS Comput. Biol. 2008, 4, e1000048. [Google Scholar] [CrossRef]
- Wang, P.; Sidney, J.; Kim, Y.; Sette, A.; Lund, O.; Nielsen, M.; Peters, B. Peptide binding predictions for HLA DR, DP and DQ molecules. BMC Bioinform. 2010, 11, 568. [Google Scholar] [CrossRef]
- Temml, V.; Kaserer, T.; Kutil, Z.; Landa, P.; Vanek, T.; Schuster, D. Pharmacophore modeling for COX-1 and -2 inhibitors with LigandScout in comparison to Discovery Studio. Future Med. Chem. 2014, 6, 1869–1881. [Google Scholar] [CrossRef]
- Shen, Y.; Maupetit, J.; Derreumaux, P.; Tufféry, P. Improved PEP-FOLD Approach for Peptide and Miniprotein Structure Prediction. J. Chem. Theory Comput. 2014, 10, 4745–4758. [Google Scholar] [CrossRef]
- Alekseenko, A.; Ignatov, M.; Jones, G.; Sabitova, M.; Kozakov, D. Protein–Protein and Protein–Peptide Docking with ClusPro Server. In Protein Structure Prediction; Methods in Molecular Biology; Humana: New York, NY, USA, 2020; pp. 157–174. [Google Scholar]
- Hennecke, J.; Wiley, D.C. Structure of a Complex of the Human α/β T Cell Receptor (TCR) HA1.7, Influenza Hemagglutinin Peptide, and Major Histocompatibility Complex Class II Molecule, HLA-DR4 (DRA*0101 and DRB1*0401). J. Exp. Med. 2002, 195, 571–581. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HLA-DRB1 Allele | Peptide Sequence | Method # | Prediction Score (1—log50K) | Affinity IC50 in nM) | Adjusted Rank (%) |
|---|---|---|---|---|---|
| HLA-DRB1*01:01 | CII259-273 | Consensus | 0.5438 | 139.2 | 35.00 |
| L-ASNase67-81 | Consensus | 0.6301 | 54.7 | 48.00 | |
| HLA-DRB1*04:01 | CII259-273 | Consensus | 0.5352 | 152.7 | 30.00 |
| L-ASNase67-81 | Consensus | 0.3810 | 810.7 | 28.00 |
| HLA-DRB1*04:01 | CII259-273 | L-ASNase67-81 | Gal264-CII259-273 |
|---|---|---|---|
| α chain | |||
| Glu(E)11 | Lys(K)264—3.18 Å | — | Lys(K)264—3.56 Å |
| Phe(F)22 | Lys(K)264—5.27 Å | Val(V)71—3.99 Å | Lys(K)264—5.27 Å |
| Ile(I)31 | Ile(I)260—4.45 Å | Ala(A)69—4.25 Å | Ile(I)260—4.45 Å |
| Phe(F)32 | — | Ala(A)69—4.12 Å | — |
| Phe(F)48 | Ile(I)260—4.58 Å | Ala(A)69—4.53 Å | Ile(I)260—4.58 Å |
| Ala(A)54 | — | Ile(I)68—4.87 Å | — |
| Ala(A)59 | — | Val(V)71—5.04 Å | — |
| Asn(N)62 | Lys(K)264—2.68 Å | — | Lys(K)264—2.68 Å |
| Val(V)65 | — | — | Lys(K)270—5.04 Å |
| Asp(D)66 | Lys(K)270—5.10 Å | — | — |
| Ile(I)72 | Pro(P)273—5.04 Å | — | Pro(P)273—5.04 Å |
| β chain | |||
| His(H)13 | Lys(K)264—4.37 Å | Val(V)76—5.33 Å | — |
| Leu(L)27 | — | — | Gal-Hyl264—2.90 Å |
| Tyr(Y)30 | Lys(K)270—3.26 Å | — | Lys(K)270—5.12 Å |
| Tyr(Y)47 | — | — | Pro(P)269—4.99 Å |
| Asp(D)57 | — | Asn(N)78—3.00 Å | — |
| Trp(W)61 | Gly(G)268—2.92 Å | Val(V)76—5.48 Å | Gly(G)268—2.92 Å |
| Tyr(Y)78 | — | Glu(E)74—4.33 Å | — |
| Asn(N)82 | Gly(G)262—3.18 Å | Ala(A)69—3.40 Å | Gly(G)262—3.18 Å |
| Val(V)85 | Ala(A)261—4.82 Å | — | — |
| Human α/β T-Cell Receptor | CII259-273 | L-ASNase67-81 | Gal264-CII259-273 |
|---|---|---|---|
| α chain | |||
| Val(V)28 | Pro(P)269—4.37 Å | Ile(I)79—4.25 Å | Pro(P)269—4.37 Å |
| Pro(P)29 | Pro(P)269—5.42 Å | — | Pro(P)269—4.37 Å |
| Glu(E)102 | — | Asp(D)78—3.09 Å | — |
| Lys(K)103 | — | Ser(S)81—2.73 Å | — |
| β chain | |||
| Leu(L)99 | Lys(K)264—5.06 Å | Glu(E)74—3.29 Å | Lys(K)264—5.06 Å |
| Gal:C1(2.25Å); Gal:C2(2.27Å); Gal:O2(2.21Å) | |||
| Tyr(Y)104 | Gly(G)265—2.90 Å | — | Gly(G)265—2.90 Å; Glu(E)266—2.82 Å; |
| Glu(E)266—2.82 Å | Ala(A)69—5.11 Å | Gal:HO2(2.41 Å); Gal:O3(2.74 Å) | |
| Tyr(Y)106 | — | Asn(N)70—2.66 Å | — |
| Characteristics | ERA Patients | Healthy Controls |
|---|---|---|
| Number of subjects | 12 | 11 |
| Sex (female, %) | 100 | 100 |
| Age (years) | 53 ± 1 (45–60) | 52 ± 2 (45–60) |
| HLA-DRB1*04:01 (%) | 100 | 100 |
| Non-smokers (%) | 100 | 100 |
| Disease duration (months) | 9 ± 1 | — |
| Anti-CCP3 level (U/mL) | 205.2 ± 26.0 | 31.5 ± 10.0 |
| RF (U/mL) | 171.8 ± 16.6 | 21.2 ± 7.6 |
| DAS28 (CRP) | 4.35 ± 0.16 | — |
| CRP (mg/L) | 22.5 ± 2.0 | — |
| Methotrexate, MTX (%) | 100 | — |
| MTX dosage (mg/week) | 10.8 ± 0.9 | — |
| Prednisolone (%) | 33.33 | — |
| Prednisolone dosage (mg/day) | 5.0 ± 0.0 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moten, D.; Teneva, I.; Apostolova, D.; Batsalova, T.; Dzhambazov, B. Molecular Mimicry of the Rheumatoid Arthritis-Related Immunodominant T-Cell Epitope within Type II Collagen (CII260-270) by the Bacterial L-Asparaginase. Int. J. Mol. Sci. 2022, 23, 9149. https://doi.org/10.3390/ijms23169149
Moten D, Teneva I, Apostolova D, Batsalova T, Dzhambazov B. Molecular Mimicry of the Rheumatoid Arthritis-Related Immunodominant T-Cell Epitope within Type II Collagen (CII260-270) by the Bacterial L-Asparaginase. International Journal of Molecular Sciences. 2022; 23(16):9149. https://doi.org/10.3390/ijms23169149
Chicago/Turabian StyleMoten, Dzhemal, Ivanka Teneva, Desislava Apostolova, Tsvetelina Batsalova, and Balik Dzhambazov. 2022. "Molecular Mimicry of the Rheumatoid Arthritis-Related Immunodominant T-Cell Epitope within Type II Collagen (CII260-270) by the Bacterial L-Asparaginase" International Journal of Molecular Sciences 23, no. 16: 9149. https://doi.org/10.3390/ijms23169149
APA StyleMoten, D., Teneva, I., Apostolova, D., Batsalova, T., & Dzhambazov, B. (2022). Molecular Mimicry of the Rheumatoid Arthritis-Related Immunodominant T-Cell Epitope within Type II Collagen (CII260-270) by the Bacterial L-Asparaginase. International Journal of Molecular Sciences, 23(16), 9149. https://doi.org/10.3390/ijms23169149