
Label-Free Quantitative Proteomics Reveal the Involvement of PRT6 in Arabidopsis thaliana Seed Responsiveness to Ethylene


 ,
,
Abstract
:1. Introduction
2. Results
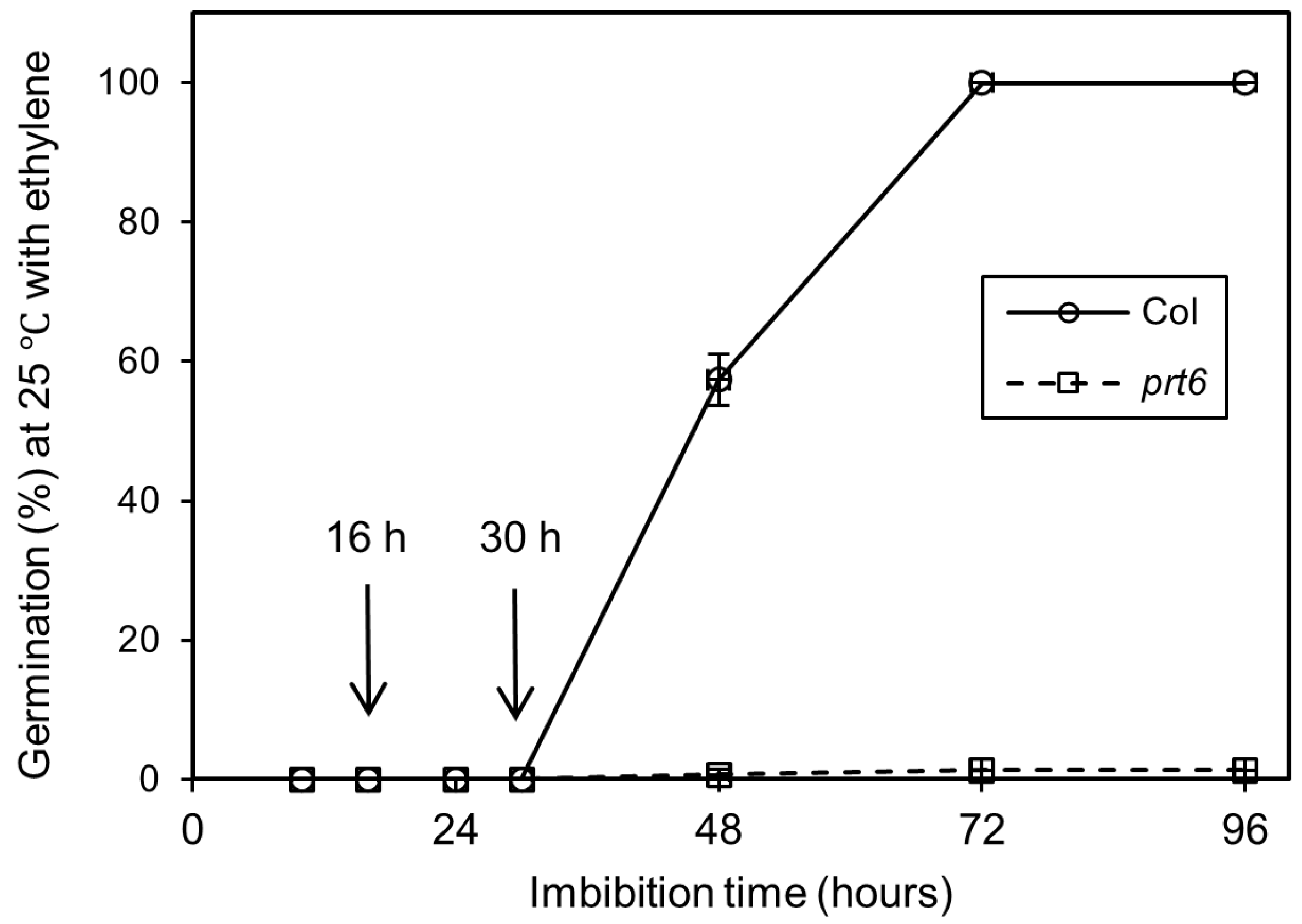
2.1. Effects of Ethylene Treatment on the Germination of Dormant Seeds
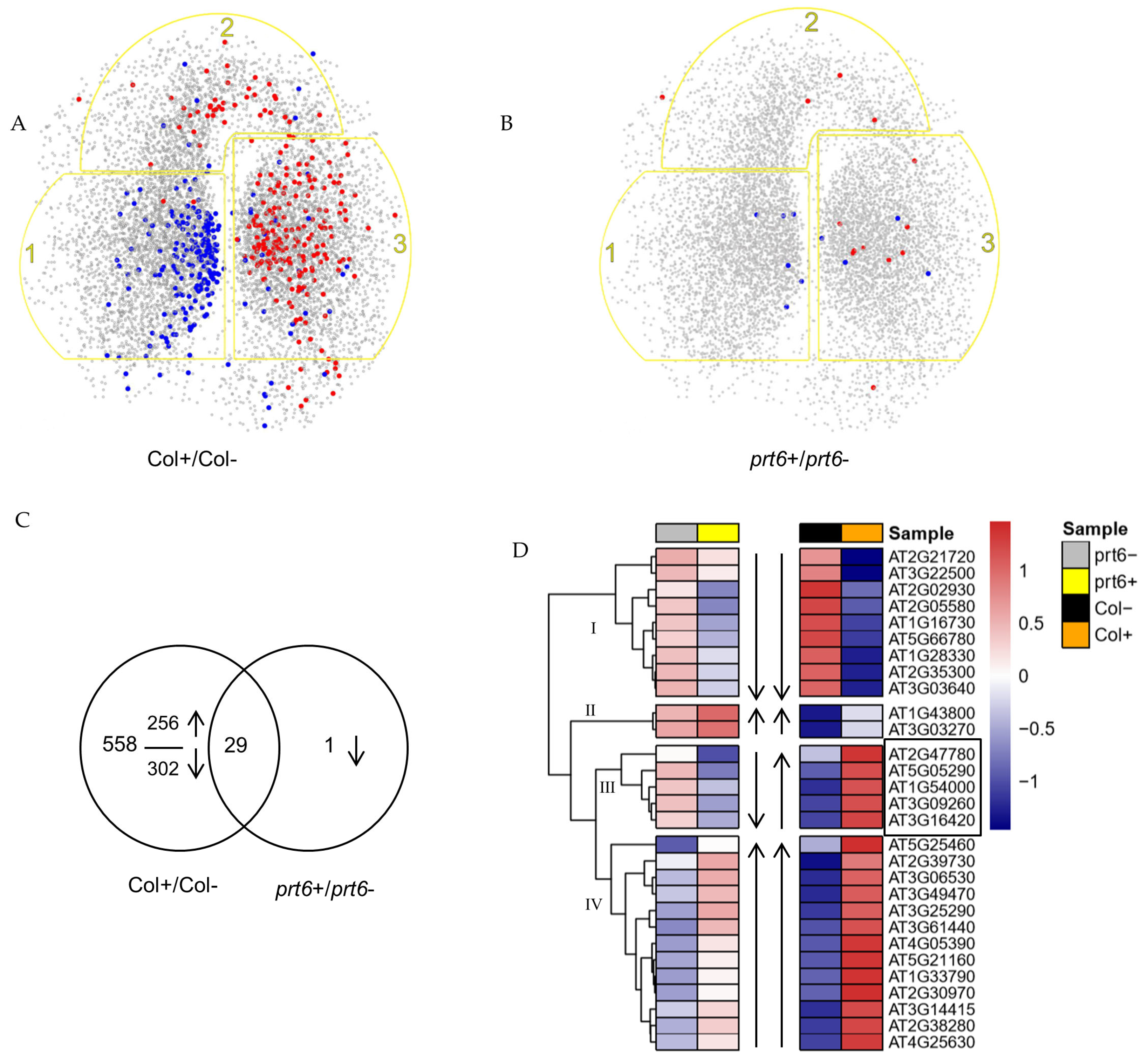
2.2. Ethylene-Induced Proteome Changes Observed at 30 h but Not 16 h after Treatments
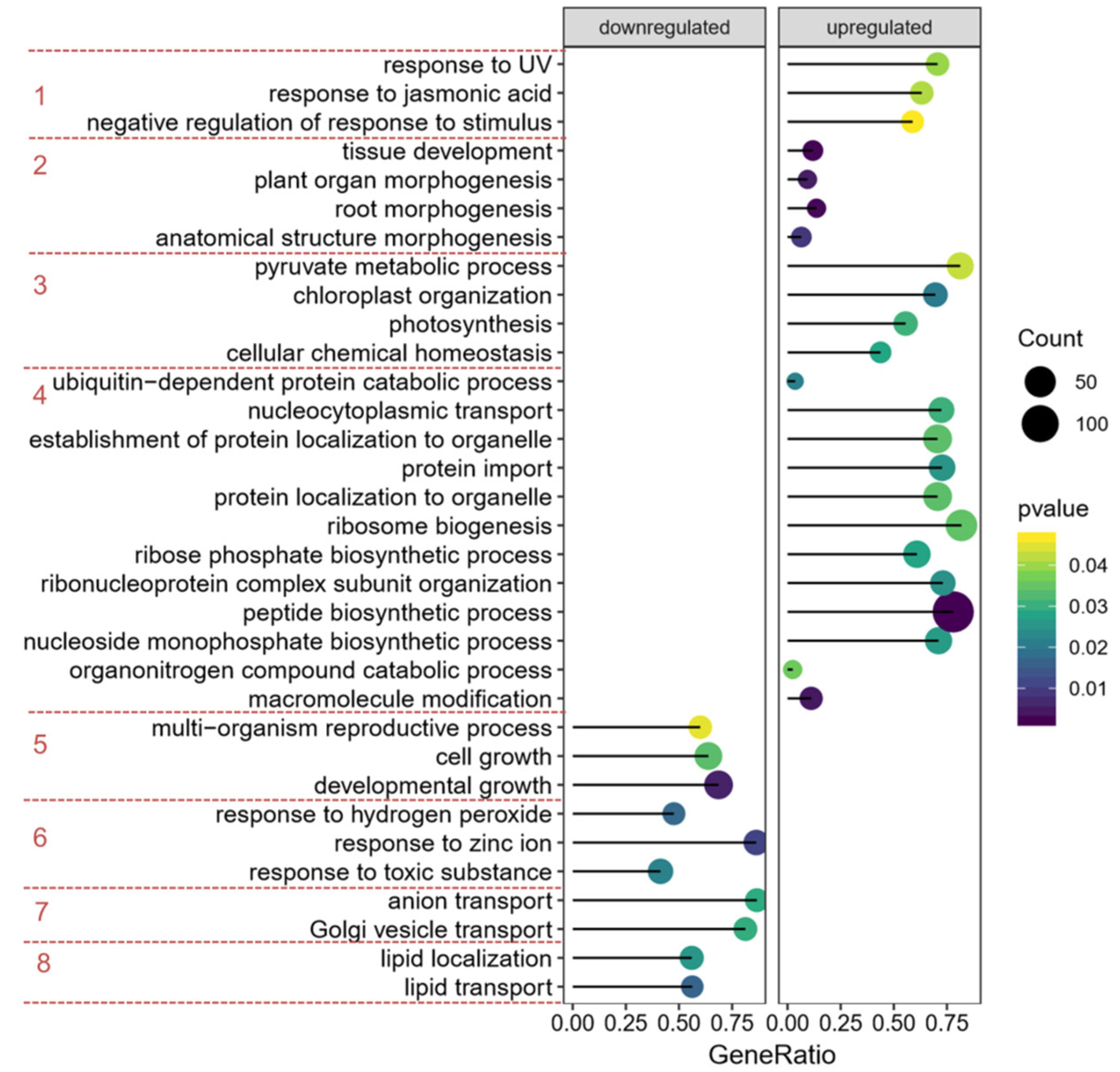
2.3. Effects of Ethylene on the Expression of DEPs
2.4. Effect of prt6 on the Expression of DEPs in the Absence of Ethylene after 30 h of Imbibition
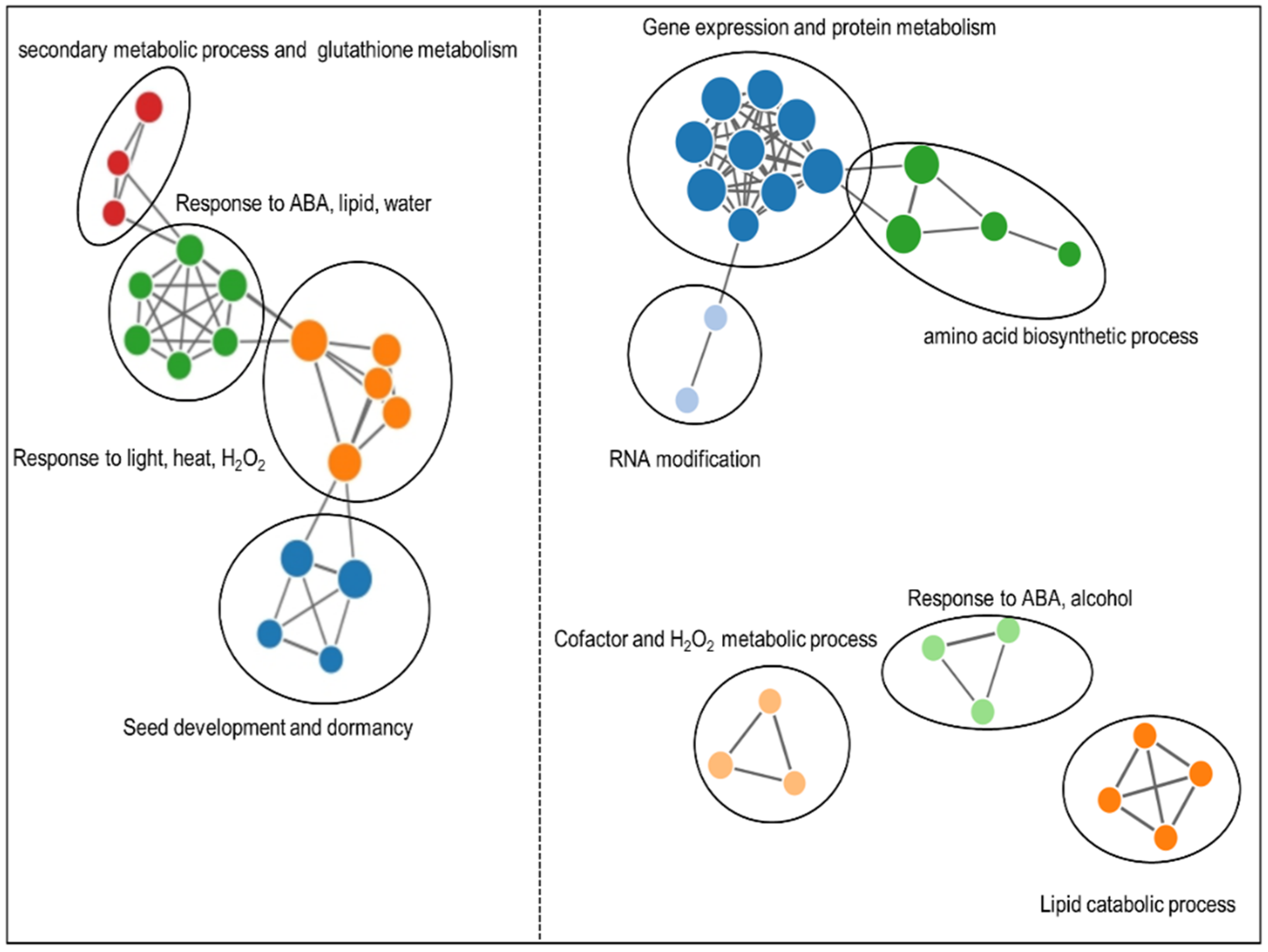
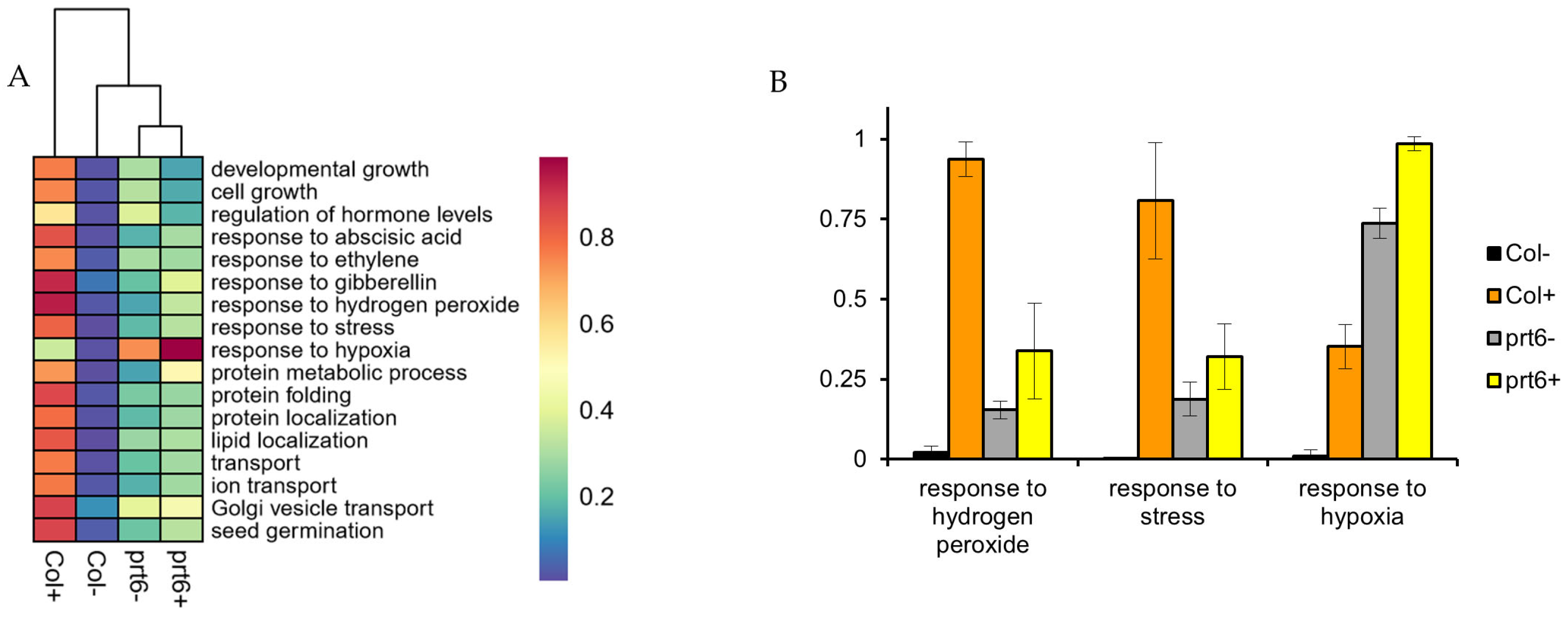
2.5. Functional Class Scoring Analysis without Filtering DEPs in 30 h-Proteome
2.6. Effect of prt6 on the Fate of Storage Proteins during Germination
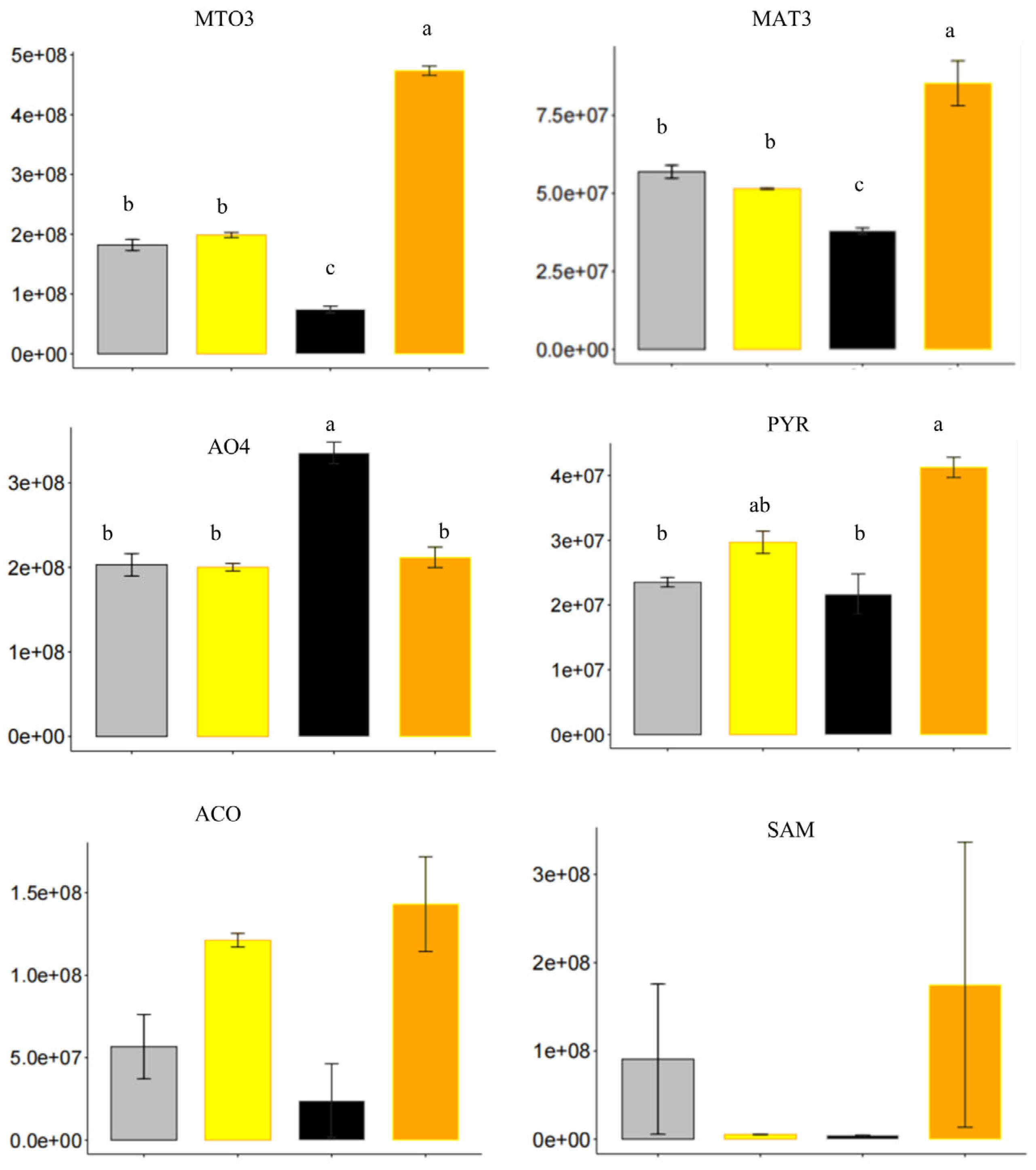
2.7. Hormone-Related Proteins in Seed
3. Discussion
4. Materials and Methods
4.1. Seed Materials
4.2. Germination Assay
4.3. Seed Treatment
4.4. Protein Extraction and LC-MS Analysis
4.5. Protein Identification and Quantification
4.6. Bioinformatic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bewley, J.D. Seed Germination and Dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Chen, Z. The Control of Seed Dormancy and Germination by Temperature, Light and Nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Leymarie, J.; Vitkauskaité, G.; Hoang, H.H.; Gendreau, E.; Chazoule, V.; Meimoun, P.; Corbineau, F.; El-Maarouf-Bouteau, H.; Bailly, C. Role of reactive oxygen species in the regulation of Arabidopsis seed dormancy. Plant Cell Physiol. 2012, 53, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yesbergenova-Cuny, Z.; Biniek, C.; Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. Revisiting the Role of Ethylene and N-End Rule Pathway on Chilling-Induced Dormancy Release in Arabidopsis Seeds. Int. J. Mol. Sci. 2018, 19, 3577. [Google Scholar] [CrossRef]
- Derkx, M.P.M.; Karssen, C.M. Variability in light, gibberellin and nitrate requirement of Arabidopsis thaliana seeds due to harvest time and conditions of dry storage. J. Plant Physiol. 1993, 141, 574–582. [Google Scholar] [CrossRef]
- Koornneef, M.; Karssen, C.M. Seed dormancy and germination. In Arabidopsis, 1st ed.; Meyerowitz, E.M., Somerville, C.R., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1994; Volume 27, pp. 313–334. [Google Scholar]
- Bethke, P.C.; Libourel, I.; Jones, R.L. Nitric oxide in seed dormancy and germination. In Annual Plant Reviews. Seed Development, Dormancy and Germination; Bradford, K., Nonogaki, H., Eds.; Wiley-Blackwell: Oxford, UK, 2007; Volume 27, pp. 153–175. [Google Scholar]
- Feurtado, J.A.; Kermode, A.R.; Bradford, K.; Nonogaki, H. A merging of paths: Abscisic acid and hormonal cross-talk in the control of seed dormancy maintenance and alleviation. In Seed Development, Dormancy and Germination; Bradford, K., Nonogaki, H., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2007; Volume 27, pp. 176–223. [Google Scholar]
- Holdsworth, M.J.; Bentsink, L.; Soppe, W.J.J. Molecular networks regulating Arabidopsis seed maturation, after-ripening, dormancy and germination. New Phytol. 2008, 179, 33–54. [Google Scholar] [CrossRef]
- Hilhorst, H.E.; Finch-Savage, W.; Buitink, J.; Bolingue, W.; Leubner-Metzger, G. Dormancy in plant seeds. In Dormancy and Resistance in Harsh Environments; Luzens, E., Cerda, J., Clark, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 43–67. [Google Scholar]
- Nambara, E.; Okamoto, M.; Tatematsu, K.; Yano, R.; Seo, M.; Kamiya, Y. Abscisic acid and the control of seed dormany and germination. Seed Sci. Res. 2010, 20, 55–67. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.D.; Xie, Q.; He, Z.H. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Linkies, A.; Müller, K.; Morris, K.; Turečková, V.; Wenk, M.; Cadman, C.S.C.; Corbineau, F.; Strnad, M.; Lynn, J.R.; Finch-Savage, W.E. Ethylene interacts with abscisic acid to regulate endosperm rupture during germination: A comparative approach using Lepidium sativum and Arabidopsis thaliana. Plant Cell 2009, 21, 3803–3822. [Google Scholar] [CrossRef] [PubMed]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 63. [Google Scholar] [CrossRef] [PubMed]
- Corbineau, F.; Xia, Q.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a key factor in the regulation of seed dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [PubMed]
- Bogatek, R.; Gniazdowska, A. Ethylene in Seed Development, Dormancy and Germination. In Annual Plant Reviews Volume 44: The Plant Hormone Ethylene; McManus, M.T., Ed.; Wiley-Blackwell: Oxford, UK, 2012; Volume 44, pp. 189–218. [Google Scholar]
- Liu, M.; Gomes, B.L.; Mila, I.; Purgatto, E.; Peres, L.E.P.; Frasse, P.; Maza, E.; Zouine, M.; Roustan, J.-P.; Bouzayen, M.; et al. Comprehensive Profiling of Ethylene Response Factor Expression Identifies Ripening-Associated ERF Genes and Their Link to Key Regulators of Fruit Ripening in Tomato. Plant Physiol. 2016, 170, 1732–1744. [Google Scholar] [CrossRef]
- Wang, X.; Gomes, M.M.; Bailly, C.; Nambara, E.; Corbineau, F. Role of ethylene and proteolytic N-degron pathway in the regulation of Arabidopsis seed dormancy. J. Integr. Plant Biol. 2021, 63, 2110–2122. [Google Scholar] [CrossRef]
- Bachmair, A.; Finley, D.; Varshavsky, A. In vivo half-life of a protein is a function of its amino-terminal residue. Science 1986, 234, 179–186. [Google Scholar] [CrossRef]
- Dissmeyer, N.; Rivas, S.; Graciet, E. Life and death of proteins after protease cleavage: Protein degradation by the N-end rule pathway. New Phytol. 2018, 218, 929–935. [Google Scholar] [CrossRef]
- Lee, K.E.; Heo, J.E.; Kim, J.M.; Hwang, C.S. N-Terminal Acetylation-Targeted N-End Rule Proteolytic System: The Ac/N-End Rule Pathway. Mol. Cells 2016, 39, 169–178. [Google Scholar]
- Dong, C.; Zhang, H.; Li, L.; Tempel, W.; Loppnau, P.; Min, J.R. Molecular basis of GID4-mediated recognition of degrons for the Pro/N-end rule pathway. Nat. Chem. Biol. 2018, 14, 466. [Google Scholar] [CrossRef]
- Dougan, D.A.; Varshavsky, A. Understanding the Pro/N-end rule pathway. Nat. Chem. Biol. 2018, 14, 415–416. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Mun, S.H.; Lee, C.S.; Hwang, C.S. Control of protein degradation by N-terminal acetylation and the N-end rule pathway. Exp. Mol. Med. 2018, 50, 91. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Lee, C.S.; Mun, S.H.; Truong, N.T.; Park, S.K.; Hwang, C.S. N-terminal acetylation and the N-end rule pathway control degradation of the lipid droplet protein PLIN2. J. Biol. Chem. 2019, 294, 379–388. [Google Scholar] [CrossRef]
- Graciet, E.; Wellmer, F. The plant N-end rule pathway: Structure and functions. Trends Plant Sci. 2010, 15, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Sriram, S.M.; Kim, B.Y.; Kwon, Y.T. The N-end rule pathway: Emerging functions and molecular principles of substrate recognition. Nat. Rev. Mol. Cell Biol. 2011, 12, 735–747. [Google Scholar] [CrossRef]
- Graciet, E.; Walter, F.; O’Maoileidigh, D.S.; Pollmann, S.; Meyerowitz, E.M.; Varshavsky, A.; Wellmer, F. The N-end rule pathway controls multiple functions during Arabidopsis shoot and leaf development. Proc. Natl. Acad. Sci. USA 2009, 106, 13618–13623. [Google Scholar] [CrossRef] [PubMed]
- Choy, M.K.; Sullivan, J.A.; Theobald, J.C.; Davies, W.J.; Gray, J.C. An Arabidopsis mutant able to green after extended dark periods shows decreased transcripts of seed protein genes and altered sensitivity to abscisic acid. J. Exp. Bot. 2008, 59, 3869–3884. [Google Scholar] [CrossRef]
- Riber, W.; Muller, J.T.; Visser, E.J.W.; Sasidharan, R.; Voesenek, L.A.C.J.; Mustroph, A. The Greening after Extended Darkness1 Is an N-End Rule Pathway Mutant with High Tolerance to Submergence and Starvation. Plant Physiol. 2015, 167, 1616–1629. [Google Scholar] [CrossRef]
- Gibbs, D.J.; Lee, S.C.; Isa, N.M.; Gramuglia, S.; Fukao, T.; Bassel, G.W.; Correia, C.S.; Corbineau, F.; Theodoulou, F.L.; Bailey-Serres, J.; et al. Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 2011, 479, 415–418. [Google Scholar] [CrossRef]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A.; Perata, P.; van Dongen, J.T. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef]
- de Marchi, R.; Sorel, M.; Mooney, B.; Fudal, I.; Goslin, K.; Kwasniewska, K.; Ryan, P.T.; Pfalz, M.; Kroymann, J.; Pollmann, S.; et al. The N-end rule pathway regulates pathogen responses in plants. Sci. Rep. 2016, 6, 26020. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Mendiondo, G.M.; Movahedi, M.; Peirats-Llobet, M.; Juan, Y.T.; Shen, Y.Y.; Dambire, C.; Smart, K.; Rodriguez, P.L.; Charng, Y.Y.; et al. The Cys-Arg/N-End Rule Pathway Is a General Sensor of Abiotic Stress in Flowering Plants. Curr. Biol. 2017, 27, 3183–3190. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Mendiondo, G.M.; Pauwels, J.; Pastor, V.; Izquierdo, Y.; Naumann, C.; Movahedi, M.; Rooney, D.; Gibbs, D.J.; Smart, K.; et al. Distinct branches of the N-end rule pathway modulate the plant immune response. New Phytol. 2019, 221, 988–1000. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Md Isa, N.; Movahedi, M.; Lozano-Juste, J.; Mendiondo, G.M.; Berckhan, S.; Marin-de la Rosa, N.; Vicente Conde, J.; Sousa Correia, C.; Pearce, S.P.; et al. Nitric oxide sensing in plants is mediated by proteolytic control of group VII ERF transcription factors. Mol. Cell 2014, 53, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Holman, T.J.; Jones, P.D.; Russell, L.; Medhurst, A.; Ubeda Tomas, S.; Talloji, P.; Marquez, J.; Schmuths, H.; Tung, S.A.; Taylor, I.; et al. The N-end rule pathway promotes seed germination and establishment through removal of ABA sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4549–4554. [Google Scholar] [CrossRef]
- Majovsky, P.; Naumann, C.; Lee, C.W.; Lassowskat, I.; Trujillo, M.; Dissmeyer, N.; Hoehenwarter, W. Targeted proteomics analysis of protein degradation in plant signaling on an LTQ-Orbitrap mass spectrometer. J. Proteome Res. 2014, 13, 4246–4258. [Google Scholar] [CrossRef]
- Zhang, H.; Deery, M.J.; Gannon, L.; Powers, S.J.; Lilley, K.S.; Theodoulou, F.L. Quantitative proteomics analysis of the Arg/N-end rule pathway of targeted degradation in Arabidopsis roots. Proteomics 2015, 15, 2447–2457. [Google Scholar] [CrossRef]
- Zhang, H.; Gannon, L.; Hassall, K.L.; Deery, M.J.; Gibbs, D.J.; Holdsworth, M.J.; van der Hoorn, R.A.L.; Lilley, K.S.; Theodoulou, F.L. N-terminomics reveals control of Arabidopsis seed storage proteins and proteases by the Arg/N-end rule pathway. New Phytol. 2018, 218, 1106–1126. [Google Scholar] [CrossRef]
- Mustroph, A.; Zanetti, M.E.; Jang, C.J.H.; Holtan, H.E.; Repetti, P.P.; Galbraith, D.W.; Girke, T.; Bailey-Serres, J. Profiling translatomes of discrete cell populations resolves altered cellular priorities during hypoxia in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18843–18848. [Google Scholar] [CrossRef]
- Bassel, G.W.; Lan, H.; Glaab, E.; Gibbs, D.J.; Gerjets, T.; Krasnogor, N.; Bonner, A.J.; Holdsworth, M.J.; Provart, N.J. Genome-wide network model capturing seed germination reveals coordinated regulation of plant cellular phase transitions. Proc. Natl. Acad. Sci. USA 2011, 108, 9709–9714. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Inter. J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Allaire, J.; Ellis, P.; Gandrud, C.; Kuo, K.; Lewis, B.; Owen, J.; Russell, K.; Rogers, J.; Sese, C.; Yetman, C. Package ‘NetworkD3′. D3 JavaScript Network Graphs from R. R Package Version 0.4. 2017. Available online: https://CRAN.R-project.org/package=networkD3 (accessed on 14 July 2022).
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Drier, Y.; Sheffer, M.; Domany, E. Pathway-based personalized analysis of cancer. Proc. Natl. Acad. Sci. USA 2013, 110, 6388–6393. [Google Scholar] [CrossRef] [PubMed]
- Melcher, K.; Xu, Y.; Ng, L.-M.; Zhou, X.E.; Soon, F.-F.; Chinnusamy, V.; Suino-Powell, K.M.; Kovach, A.; Tham, F.S.; Cutler, S.R.; et al. Identification and mechanism of ABA receptor antagonism. Nat. Struct. Mol. Biol. 2010, 17, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Colasuonno, P.; Marcotuli, I.; Lozito, M.L.; Simeone, R.; Blanco, A.; Gadaleta, A. Characterization of Aldehyde Oxidase (AO) Genes Involved in the Accumulation of Carotenoid Pigments in Wheat Grain. Front. Plant Sci. 2017, 8, 863. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.M.; Melcher, K.; Teh, B.T.; Xu, H.E. Abscisic acid perception and signaling: Structural mechanisms and applications. Acta Pharmacol. Sin. 2014, 35, 567–584. [Google Scholar] [CrossRef]
- Yang, D.; Li, Y.; Xiao, H.; Liu, Q.; Zhang, M.; Zhu, J.; Ma, W.; Yao, C.; Wang, J.; Wang, D.; et al. Gaining confidence in biological interpretation of the microarray data: The functional consistence of the significant GO categories. Bioinformatics 2008, 24, 265–271. [Google Scholar] [CrossRef]
- Hong, G.; Zhang, W.; Li, H.; Shen, X.; Guo, Z. Separate enrichment analysis of pathways for up- and downregulated genes. J. R. Soc. Interface 2014, 11, 20130950. [Google Scholar] [CrossRef]
- Galland, M.; Job, D.; Rajjou, L. The seed proteome web portal. Front. Plant Sci. 2012, 3, 98. [Google Scholar] [CrossRef]
- Licausi, F.; van Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J. 2010, 62, 302–315. [Google Scholar] [CrossRef]
- Sasidharan, R.; Mustroph, A. Plant oxygen sensing is mediated by the N-end rule pathway: A milestone in plant anaerobiosis. Plant Cell 2011, 23, 4173–4183. [Google Scholar] [CrossRef] [PubMed]
- Bui, L.T.; Giuntoli, B.; Kosmacz, M.; Parlanti, S.; Licausi, F. Constitutively expressed ERF-VII transcription factors redundantly activate the core anaerobic response in Arabidopsis thaliana. Plant Sci. 2015, 236, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Conde, J.V.; Berckhan, S.; Prasad, G.; Mendiondo, G.M.; Holdsworth, M.J. Group VII Ethylene Response Factors Coordinate Oxygen and Nitric Oxide Signal Transduction and Stress Responses in Plants. Plant Physiol. 2015, 169, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Gasch, P.; Fundinger, M.; Muller, J.T.; Lee, T.; Bailey-Serres, J.; Mustroph, A. Redundant ERF-VII Transcription Factors Bind to an Evolutionarily Conserved cis-Motif to Regulate Hypoxia-Responsive Gene Expression in Arabidopsis. Plant Cell 2016, 28, 160–180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gannon, L.; Jones, P.D.; Rundle, C.A.; Hassall, K.L.; Gibbs, D.J.; Holdsworth, M.J.; Theodoulou, F.L. Genetic interactions between ABA signalling and the Arg/N-end rule pathway during Arabidopsis seedling establishment. Sci. Rep. 2018, 8, 15192. [Google Scholar] [CrossRef]
- Abbas, M.; Berckhan, S.; Rooney, D.J.; Gibbs, D.J.; Vicente Conde, J.; Sousa Correia, C.; Bassel, G.W.; Marin-de la Rosa, N.; Leon, J.; Alabadi, D.; et al. Oxygen sensing coordinates photomorphogenesis to facilitate seedling survival. Curr. Biol. 2015, 25, 1483–1488. [Google Scholar] [CrossRef]
- Matilla, A.J.; Matilla-Vázquez, M.A. Involvement of ethylene in seed physiology. Plant Sci. 2008, 175, 87–97. [Google Scholar] [CrossRef]
- Liu, X.; Shiomi, S.; Nakatsuka, A.; Kubo, Y.; Nakamura, R.; Inaba, A. Characterization of ethylene biosynthesis associated with ripening in banana fruit. Plant Physiol. 1999, 121, 1257–1266. [Google Scholar] [CrossRef]
- Beaudoin, N.; Serizet, C.; Gosti, F.; Giraudat, J. Interactions between abscisic acid and ethylene signaling cascades. Plant Cell 2000, 12, 1103–1115. [Google Scholar] [CrossRef]
- Castillo, M.C.; Lozano-Juste, J.; Gonzalez-Guzman, M.; Rodriguez, L.; Rodriguez, P.L.; Leon, J. Inactivation of PYR/PYL/RCAR ABA receptors by tyrosine nitration may enable rapid inhibition of ABA signaling by nitric oxide in plants. Sci. Signal. 2015, 8, ra89. [Google Scholar] [CrossRef]
- Desjardin, C.; Balliau, T.; Valot, B.; Zivy, M.; Wimel, L.; Guerin, G.; Cribiu, E.; Schibler, L. A method for proteomic analysis of equine subchondral bone and epiphyseal cartilage. Proteomics 2012, 12, 1870–1874. [Google Scholar] [CrossRef]
- Craig, R.; Beavis, R.C. TANDEM: Matching proteins with tandem mass spectra. Bioinformatics 2004, 20, 1466–1467. [Google Scholar] [CrossRef]
- Langella, O.; Valot, B.; Balliau, T.; Blein-Nicolas, M.; Bonhomme, L.; Zivy, M. X!TandemPipeline: A Tool to Manage Sequence Redundancy for Protein Inference and Phosphosite Identification. J. Proteome Res. 2017, 16, 494–503. [Google Scholar] [CrossRef]
- Valot, B.; Langella, O.; Nano, E.; Zivy, M. MassChroQ: A versatile tool for mass spectrometry quantification. Proteomics 2011, 11, 3572–3577. [Google Scholar] [CrossRef]
- Belouah, I.; Blein-Nicolas, M.; Balliau, T.; Gibon, Y.; Zivy, M.; Colombié, S. Peptide filtering differently affects the performances of XIC-based quantification methods. Proteomics 2019, 193, 131–141. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Package ‘Factoextra’: Extract and Visualize the Results of Multivariate Data Analyses. R package version 1.0.7. 2017. Available online: http://www.sthda.com/english/rpkgs/factoextra (accessed on 14 July 2022).
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. 2019. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 14 July 2022).
- Blighe, K.; Rana, S.; Lewis, M. EnhancedVolcano: Publication-Ready Volcano Plots with Enhanced Colouring and Labeling. R package Version 1.8.0. 2019. Available online: https://github.com/kevinblighe/EnhancedVolcano (accessed on 14 July 2022).
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R package for comparing biological themes among gene clusters. Omics A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Duration of Seed Incubation in the Presence of Ethylene (h) | Germination (%) after 7 Days at 25 °C in Air after Ethylene Treatment | |
|---|---|---|
| 2016 Harvest | 2018 Harvest | |
| 0 | 11.9 ± 4.0 | 0 |
| 10 | 39.9 ± 3.0 | - |
| 16 | 48.3 ± 4.0 | 7.7 ± 7.5 |
| 24 | 54.5 ± 6.2 | 21.0 ± 1.0 |
| 30 | - | 54.6 ± 4.6 |
| 48 | 61.9 ± 3.7 | 81.3 ± 4.0 |
| ID. | Description | prt6−/Col− |
|---|---|---|
| Hypoxia-responsive | ||
| AT1G19530 | DNA polymerase epsilon catalytic subunit A | 1.64 |
| AT2G16060 | Hemoglobin 1 | 18.74 |
| AT1G43800 | Plant stearoyl-acyl-carrier-protein desaturase family protein | 21.64 |
| Carbohydrate metabolism | ||
| AT4G33820 | Glycosyl hydrolase superfamily protein | 0.39 |
| AT5G63800 | Glycosyl hydrolase family 35 protein | 0.48 |
| AT3G09260 | Glycosyl hydrolase superfamily protein | 5.78 |
| AT1G53580 | Glyoxalase II 3 | 1.52 |
| AT4G34260 | 1,2-alpha-L-fucosidase | 0.70 |
| AT5G26120 | Alpha-L-arabinofuranosidase 2 | 0.65 |
| AT4G15210 | Beta-amylase 5 | 1.98 |
| AT4G36360 | Beta-galactosidase 3 | 0.60 |
| AT5G49360 | Beta-xylosidase 1 | 1.51 |
| AT1G02640 | Beta-xylosidase 2 | 1.47 |
| AT5G57550 | Xyloglucan endotransglucosylase/hydrolase 25 | 0.47 |
| AT2G47510 | Fumarase 1 | 1.30 |
| AT1G12780 | UDP-D-glucose/UDP-D-galactose 4-epimerase 1 | 1.57 |
| AT2G39730 | Rubisco activase | 1.80 |
| Cell wall | ||
| AT1G19900 | Glyoxal oxidase-related protein (involved in cell wall modification) | 0.51 |
| AT5G15490 | UDP-glucose 6-dehydrogenase protein (required for the formation of cell wall ingrowths) | 1.47 |
| AT5G05290 | Expansin A2 | 4.24 |
| AT3G14310 | Pectin methylesterase 3 | 1.32 |
| Oxidation and reduction | ||
| AT3G61070 | Peroxin 11E (integral to peroxisome membrane, controls peroxisome proliferation) | 1.34 |
| AT2G46750 | D-arabinono-1,4-lactone oxidase activity | 0.51 |
| AT3G14415 | Aldolase-type TIM barrel family protein (modulates ROS-mediated signal transduction) | 1.95 |
| AT4G35000 | Ascorbate peroxidase 3 | 1.67 |
| AT4G20860 | FAD-binding Berberine family protein (involved in the generation of H2O2) | 0.53 |
| AT2G02930 | Glutathione S-transferase F3 | 0.52 |
| AT2G30870 | Glutathione S-transferase PHI 10 | 1.95 |
| AT2G47730 | Glutathione S-transferase phi 8 | 2.14 |
| AT5G66920 | SKU5 similar 17 (oxidation-reduction process) | 1.63 |
| AT3G27820 | Monodehydroascorbate reductase 4 | 1.83 |
| AT1G60680 | NAD(P)-linked oxidoreductase superfamily protein | 0.66 |
| AT3G44880 | Pheophorbide a oxygenase family protein with Rieske 2Fe-2S domain-containing protein | 1.38 |
| AT5G63030 | Thioredoxin superfamily protein | 0.67 |
| Transcription | ||
| AT5G02490 | Heat shock protein 70 (HSP 70) (mediator of RNA polymerase II transcription subunit 37c) | 1.38 |
| AT4G25630 | Ibrillarin 2 (Mediator of RNA polymerase II transcription subunit 36a) | 1.45 |
| AT3G08030 | DNA-directed RNA polymerase subunit beta | 1.77 |
| AT3G03270 | Adenine nucleotide alpha hydrolases-like superfamily protein | 7.91 |
| AT1G74260 | Purine biosynthesis 4 | 1.61 |
| AT3G57610 | Adenylosuccinate synthase (purine synthesis) | 1.50 |
| AT2G45300 | RNA 3’-terminal phosphate cyclase/enolpyruvate transferase%2C alpha/beta | 1.80 |
| AT3G52150 | RNA-binding (RRM/RBD/RNP motifs) family protein | 1.37 |
| AT3G19130 | RNA-binding protein 47B | 1.86 |
| AT5G63420 | RNA-metabolising metallo-beta-lactamase family protein | 1.34 |
| AT3G58510 | DEA(D/H)-box RNA helicase family protein | 1.31 |
| Translation | ||
| AT4G01310 | Ribosomal L5P family protein | 1.35 |
| AT5G23900 | Ribosomal protein L13e family protein | 1.38 |
| AT3G63490 | Ribosomal protein L1p/L10e family | 1.44 |
| AT1G50920 | Nucleolar GTP-binding protein ( Involved in the biogenesis of the 60S ribosomal subunit) | 1.57 |
| ATCG00830 | Ribosomal protein L2 | 1.67 |
| AT2G33800 | Ribosomal protein S5 family protein | 1.67 |
| AT1G32990 | Plastid ribosomal protein l11 | 1.91 |
| AT2G40290 | Eukaryotic translation initiation factor 2 subunit 1 | 1.39 |
| AT3G56150 | Eukaryotic translation initiation factor 3C | 1.74 |
| AT4G20360 | RAB GTPase homolog E1B (Elongation factor Tu) | 1.55 |
| Protein folding | ||
| AT3G13470 | TCP-1/cpn60 chaperonin family protein (protein folding) | 1.49 |
| AT1G56340 | Calreticulin 1a (Molecular calcium-binding chaperone promoting folding) | 2.38 |
| AT1G09210 | Calreticulin 1b (Molecular calcium-binding chaperone promoting folding) | 2.07 |
| AT3G09440 | Heat shock protein 70 (HSP70) (mediate the folding of newly translated peptides) | 1.48 |
| AT1G55490 | Chaperonin 60 beta (suppressing protein aggregation in vitro) | 1.34 |
| AT3G16420 | PYK10-binding protein 1 (chaperone that facilitates the correct polymerization of PYK10) | 4.71 |
| Protein transport | ||
| AT4G10480 | Nascent polypeptide-associated complex (NAC) 2C alpha | 1.50 |
| AT3G49470 | Nascent polypeptide-associated complex subunit alpha-like protein 2 | 1.40 |
| AT3G15980 | Coatomer subunit beta -3 | 1.44 |
| AT3G46830 | RAB GTPase homolog A2C | 1.44 |
| Protein modification | ||
| AT4G17040 | CLP protease R subunit 4 | 1.38 |
| AT2G47390 | Prolyl oligopeptidase family protein | 1.46 |
| AT2G38280 | AMP deaminase%2C putative / myoadenylate deaminase | 1.34 |
| AT3G45010 | Serine carboxypeptidase-like 48 | 3.57 |
| AT5G10240 | Asparagine synthetase 3 | 1.45 |
| AT5G11880 | Pyridoxal-dependent decarboxylase family protein | 1.33 |
| AT1G61790 | Oligosaccharyltransferase complex/protein N-linked glycosylation | 1.41 |
| AT1G48630 | Receptor for activated C kinase 1B (shuttle activated protein kinase C ) | 2.04 |
| AT3G18130 | Receptor for activated C kinase 1C (shuttle activated protein kinase C ) | 1.66 |
| AT1G25490 | ARM repeat superfamily protein (encoding phosphoprotein phosphatase) | 1.69 |
| LEAs | ||
| AT3G22500 | Seed maturation protein (LEA protein) | 0.63 |
| AT3G51810 | Stress induced protein (LEA protein) | 0.69 |
| AT5G66780 | Late embryogenesis abundant protein | 0.55 |
| Hormone | ||
| AT1G04580 | Aldehyde oxidase 4 (ABA synthesis) | 0.61 |
| AT3G17390 | S-adenosylmethionine synthetase family protein (ethylene synthesis) | 2.45 |
| AT2G36880 | Methionine adenosyltransferase 3 (ethylene synthesis) | 1.50 |
| Locus | Description | 0 | 16 | 30 |
|---|---|---|---|---|
| AT5G44120.3 | Cruciferin A | 1.44 | 2.73 | 0.56 |
| AT5G44120.1 | Cruciferin A | 1.13 | 1.74 | 0.88 |
| AT1G03880 | Cruciferin B | 1.12 | 1.43 | 0.81 |
| AT4G28520 | Cruciferin C | 1.00 | 1.04 | 0.73 |
| AT4G30880 | 2S albumin | 1.59 | 1.42 | 0.90 |
| AT3G22640 | Cupin | 0.97 | 1.14 | 0.73 |
| AT1G07750 | RmlC-like cupins | 0.73 | 1.65 | 1.05 |
| AT4G36700 | RmlC-like cupins | 0.77 | 1.19 | 0.81 |
| AT1G03890 | RmlC-like cupins | 0.85 | 1.01 | 0.75 |
| AT2G28490 | RmlC-like cupins | 0.94 | 1.09 | 0.83 |
| AT2G18540 | RmlC-like cupins | 1.07 | 1.17 | 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Davanture, M.; Zivy, M.; Bailly, C.; Nambara, E.; Corbineau, F. Label-Free Quantitative Proteomics Reveal the Involvement of PRT6 in Arabidopsis thaliana Seed Responsiveness to Ethylene. Int. J. Mol. Sci. 2022, 23, 9352. https://doi.org/10.3390/ijms23169352
Wang X, Davanture M, Zivy M, Bailly C, Nambara E, Corbineau F. Label-Free Quantitative Proteomics Reveal the Involvement of PRT6 in Arabidopsis thaliana Seed Responsiveness to Ethylene. International Journal of Molecular Sciences. 2022; 23(16):9352. https://doi.org/10.3390/ijms23169352
Chicago/Turabian StyleWang, Xu, Marlène Davanture, Michel Zivy, Christophe Bailly, Eiji Nambara, and Françoise Corbineau. 2022. "Label-Free Quantitative Proteomics Reveal the Involvement of PRT6 in Arabidopsis thaliana Seed Responsiveness to Ethylene" International Journal of Molecular Sciences 23, no. 16: 9352. https://doi.org/10.3390/ijms23169352