
Rosavin Ameliorates Hepatic Inflammation and Fibrosis in the NASH Rat Model via Targeting Hepatic Cell Death

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
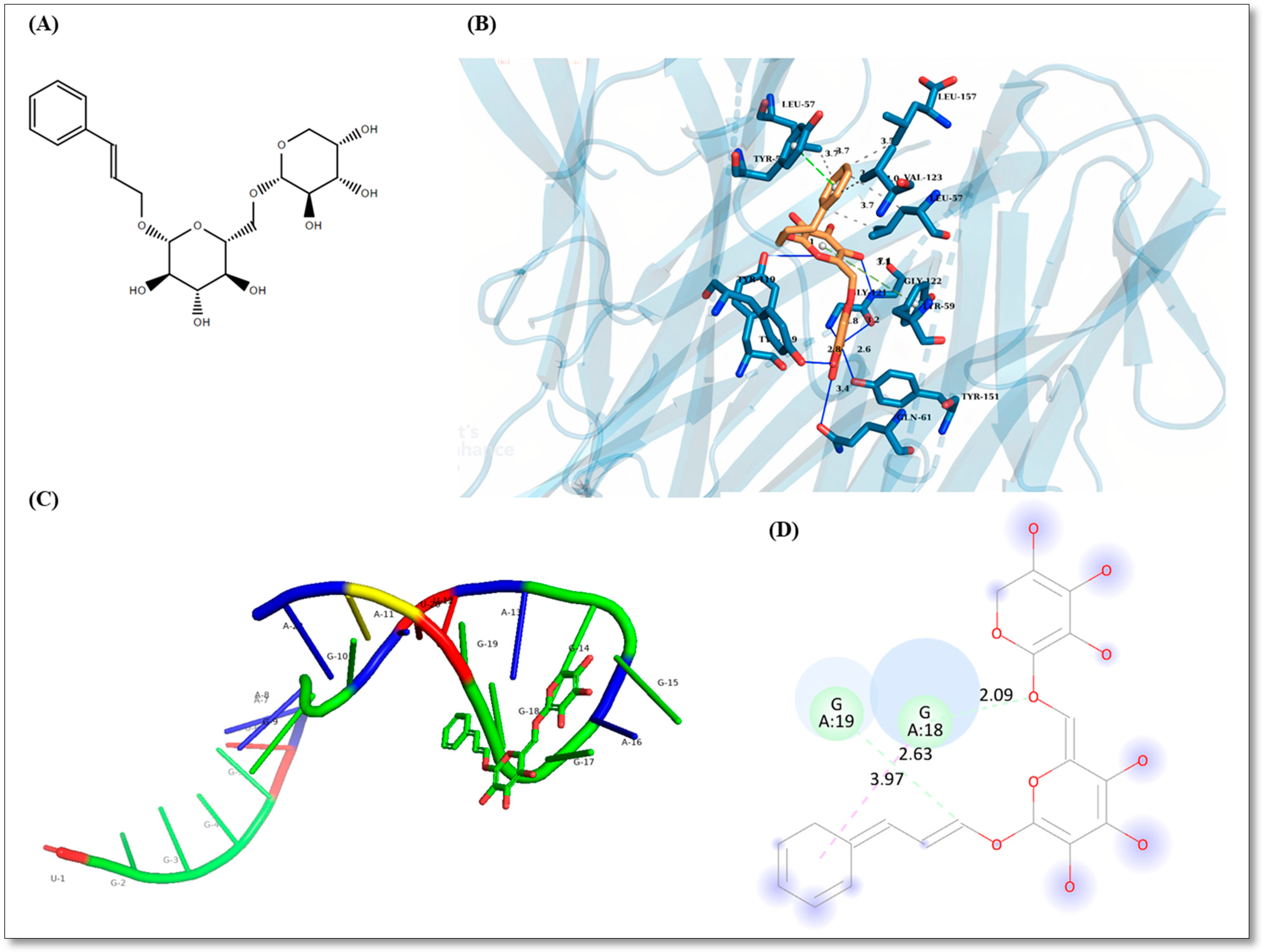
2.1. Rosavin-Protein Interaction Analysis by Blind Docking
2.2. Rosavin-miRNA-6881-5P Analysis
2.3. The Effect of Rosavin Treatment on Liver Function and Lipid Profile
2.4. Histological Findings
2.5. The Effect of Rosavin Treatment on the Expression Profile of Hepatic HSPD1/TNF/MMP14/ITGB1/miRNA-6881-5P/lnc-SPARCL1-1:2 Panel
2.6. The Effect of Rosavin Treatment on the Protein Expression of Hepatic Caspase-3, TNF-α and IL-6
2.7. Correlation among Hepatic IL-6, TNF-α, Caspase-3 and Cell Death-Related RNA Panel
3. Discussion
4. Material and Methods
4.1. Chemicals and Drugs
4.2. Experimental Animals and Design
4.3. Blood Sampling and Liver Tissue Collection
4.4. Serum Biochemical Assays (Liver Function and Lipid Profile Markers)
4.5. Hepatic Histological Evaluation
4.6. Bioinformatics Set Up
4.6.1. Construction of the RNAs-Based Panel
4.6.2. Rosavin-Protein Interaction Analysis by Blind Docking
4.6.3. Rosavin-miRNA-6881-5P Docking Interaction
4.7. Total RNA Extraction (mRNA, miRNA and lncRNA)
4.8. Quantitative Polymerase Chain Reaction (qPCR)
4.9. Immunostaining of Hepatic IL-6, TNF-α and Caspase-3
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NASH | Non-alcoholic steatohepatitis |
| HSPD1 | Heat shock protein Family D Member 1 |
| TNF | tumor necrosis factor |
| MMP14 | Matrix metalloproteinase 14 |
| ITGB1 | integrin β1 |
References
- Pouwels, S.; Sakran, N.; Graham, Y.; Leal, A.; Pintar, T.; Yang, W.; Kassir, R.; Singhal, R.; Mahawar, K.; Ramnarain, D. Non-alcoholic fatty liver disease (NAFLD): A review of pathophysiology, clinical management and effects of weight loss. BMC Endocr. Disord. 2022, 22, 63. [Google Scholar] [CrossRef] [PubMed]
- Gadallah, S.H.; Eissa, S.; Ghanem, H.M.; Ahmed, E.K.; Hasanin, A.H.; El Mahdy, M.M.; Matboli, M. Probiotic-prebiotic-synbiotic modulation of (YAP1, LATS1 and NF2 mRNAs/miR-1205/lncRNA SRD5A3-AS1) panel in NASH animal model. Biomed. Pharmacother. 2021, 140, 111781. [Google Scholar] [CrossRef] [PubMed]
- Tzanaki, I.; Agouridis, A.P.; Kostapanos, M.S. Is there a role of lipid-lowering therapies in the management of fatty liver disease? World J. Hepatol. 2022, 27, 119. [Google Scholar] [CrossRef] [PubMed]
- Albadawy, R.; Agwa, S.H.A.; Khairy, E.; Saad, M.; El Touchy, N.; Othman, M.; Matboli, M. Clinical significance of hspd1/mmp14/itgb1/mir-6881-5p/lnc-sparcl1-1:2 rna panel in nafld/nash diagnosis: Egyptian pilot study. Biomedicines 2021, 9, 1248. [Google Scholar] [CrossRef]
- Byrne, C.D.; Targher, G. NAFLD: A multisystem disease. J. Hepatol. 2015, 62, S47–S64. [Google Scholar] [CrossRef]
- Ibrahim, S.H. Sinusoidal endotheliopathy in nonalcoholic steatohepatitis: Therapeutic implications. Am. J. Physiol.-Gastrointest. Liver Physiol. 2021, 321, G67–G74. [Google Scholar] [CrossRef]
- Nasiri-Ansari, N.; Androutsakos, T.; Flessa, C.; Kyrou, I.; Siasos, G.; Randeva, H.S.; Kassi, E.; Papavassiliou, A.G. Endothelial Cell Dysfunction and Nonalcoholic Fatty Liver Disease (NAFLD): A Concise Review. Cell 2022, 11, 2511. [Google Scholar] [CrossRef]
- Lafoz, E.; Ruart, M.; Anton, A.; Oncins, A.; Hernández-Gea, V. The Endothelium as a Driver of Liver Fibrosis and Regeneration. Cells 2020, 9, 929. [Google Scholar] [CrossRef]
- Duarte, S.; Baber, J.; Fujii, T.; Coito, A.J. Matrix metalloproteinases in liver injury, repair and fibrosis. Matrix Biol. 2015, 44, 147–156. [Google Scholar] [CrossRef]
- Sulaiman, S.A.; Muhsin, N.I.A.; Jamal, R. Regulatory non-coding RNAs network in non-alcoholic fatty liver disease. Front. Physiol. 2019, 10, 279. [Google Scholar] [CrossRef]
- Matboli, M.; Gadallah, S.H.; Rashed, W.M.; Hasanin, A.H.; Essawy, N.; Ghanem, H.M.; Eissa, S. Mrna-mirna-lncrna regulatory network in nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2021, 22, 6770. [Google Scholar] [CrossRef]
- Soronen, J.; Yki-Järvinen, H.; Zhou, Y.; Sädevirta, S.; Sarin, A.P.; Leivonen, M.; Sevastianova, K.; Perttilä, J.; Laurila, P.P.; Sigruener, A.; et al. Novel hepatic microRNAs upregulated in human nonalcoholic fatty liver disease. Physiol. Rep. 2016, 4, e12661. [Google Scholar] [CrossRef]
- Sun, C.; Liu, X.; Yi, Z.; Xiao, X.; Yang, M.; Hu, G.; Liu, H.; Liao, L.; Huang, F. Genome-wide analysis of long noncoding RNA expression profiles in patients with non-alcoholic fatty liver disease. IUBMB Life 2015, 67, 847–852. [Google Scholar] [CrossRef]
- Wang, M.; Luo, L.; Yao, L.; Wang, C.; Jiang, K.; Liu, X.; Xu, M.; Shen, N.; Guo, S.; Sun, C.; et al. Salidroside improves glucose homeostasis in obese mice by repressing inflammation in white adipose tissues and improving leptin sensitivity in hypothalamus. Sci. Rep. 2016, 6, 25399. [Google Scholar] [CrossRef]
- Zhang, K.; Si, X.P.; Huang, J.; Han, J.; Liang, X.; Xu, X.B.; Wang, Y.T.; Li, G.Y.; Wang, H.Y.; Wang, J.H. Preventive effects of Rhodiola rosea L. On bleomycin-induced pulmonary fibrosis in rats. Int. J. Mol. Sci. 2016, 17, 879. [Google Scholar] [CrossRef]
- Tang, H.; Gao, L.; Mao, J.; He, H.; Liu, J.; Cai, X.; Lin, H.; Wu, T. Salidroside protects against bleomycin-induced pulmonary fibrosis: Activation of Nrf2-antioxidant signaling, and inhibition of NF-κB and TGF-β1/Smad-2/-3 pathways. Cell Stress Chaperones 2016, 21, 239–249. [Google Scholar] [CrossRef]
- Marchev, A.S.; Dimitrova, P.; Koycheva, I.K.; Georgiev, M.I. Altered expression of TRAIL on mouse T cells via ERK phosphorylation by Rhodiola rosea L. and its marker compounds. Food Chem. Toxicol. 2017, 108, 419–428. [Google Scholar] [CrossRef]
- Pu, W.L.; Zhang, M.Y.; Bai, R.Y.; Sun, L.K.; Li, W.H.; Yu, Y.L.; Zhang, Y.; Song, L.; Wang, Z.X.; Peng, Y.F.; et al. Anti-inflammatory effects of Rhodiola rosea L.: A review. Biomed. Pharmacother. 2020, 121, 109552. [Google Scholar] [CrossRef]
- Ladewig, E.; Okamura, K.; Flynt, A.S.; Westholm, J.O.; Lai, E.C. Discovery of hundreds of mirtrons in mouse and human small RNA data. Genome Res. 2012, 22, 1634–1645. [Google Scholar] [CrossRef]
- Puengel, T.; Liu, H.; Guillot, A.; Heymann, F.; Tacke, F.; Peiseler, M. Nuclear Receptors Linking Metabolism, Inflammation, and Fibrosis in Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2022, 23, 2668. [Google Scholar] [CrossRef]
- Hirsova, P.; Bohm, F.; Dohnalkova, E.; Nozickova, B.; Heikenwalder, M.; Gores, G.J.; Weber, A. Hepatocyte apoptosis is tumor promoting in murine nonalcoholic steatohepatitis. Cell Death Dis. 2020, 11, 80. [Google Scholar] [CrossRef]
- Hirsova, P.; Gores, G.J. Death Receptor-Mediated Cell Death and Proinflammatory Signaling in Nonalcoholic Steatohepatitis. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 17–27. [Google Scholar] [CrossRef]
- Huang, Y.H.; Wang, F.S.; Wang, P.W.; Lin, H.Y.; Luo, S.D.; Yang, Y.L. Heat Shock Protein 60 Restricts Release of Mitochondrial dsRNA to Suppress Hepatic Inflammation and Ameliorate Non-Alcoholic Fatty Liver Disease in Mice. Int. J. Mol. Sci. 2022, 23, 577. [Google Scholar] [CrossRef] [PubMed]
- Hoter, A.; Rizk, S.; Naim, H.Y. Heat Shock Protein 60 in Hepatocellular Carcinoma: Insights and Perspectives. Front. Mol. Biosci. 2020, 7, 60. [Google Scholar] [CrossRef]
- Enomoto, H.; Mittal, N.; Inomata, T.; Arimura, T.; Izumi, T.; Kimura, A.; Fukuda, K.; Makino, S. Dilated cardiomyopathy-linked heat shock protein family D member 1 mutations cause up-regulation of reactive oxygen species and autophagy through mitochondrial dysfunction. Cardiovasc. Res. 2021, 117, 1118–1131. [Google Scholar] [CrossRef]
- Li, R.; Toan, S.; Zhou, H. Role of mitochondrial quality control in the pathogenesis of nonalcoholic fatty liver disease. Aging 2020, 12, 6467. [Google Scholar] [CrossRef]
- Singh, P. Evaluation of Pro-Inflammatory Markers IL-6 and TNF-a and their Correlation with Non-Alcoholic Fatty Liver Disease. J. Adv. Res. Med. 2019, 6. [Google Scholar] [CrossRef]
- Eman, E.; Hemat, E.; Hanaa, H.; Safwat, Q. Effects of berberine on high-fat/high-sucrose-induced nonalcoholic steatohepatitis in experimental rats. Tanta Med. J. 2019, 47, 80. [Google Scholar] [CrossRef]
- Roeb, E. Matrix metalloproteinases and liver fibrosis (translational aspects). Matrix Biol. 2018, 68, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Niioka, M.; Ishikawa, A.; Hozawa, S.; Arai, M.; Maruyama, K.; Okada, A.; Okazaki, I. Dynamic change of cells expressing MMP-2 mRNA and MT1-MMP mRNA in the recovery from liver fibrosis in the rat. J. Hepatol. 2001, 35, 465–473. [Google Scholar] [CrossRef]
- Guo, Q.; Furuta, K.; Lucien, F.; Gutierrez Sanchez, L.H.; Hirsova, P.; Krishnan, A.; Kabashima, A.; Pavelko, K.D.; Madden, B.; Alhuwaish, H.; et al. Integrin β1-enriched extracellular vesicles mediate monocyte adhesion and promote liver inflammation in murine NASH. J. Hepatol. 2019, 71, 1193–1205. [Google Scholar] [CrossRef]
- Kanda, T.; Matsuoka, S.; Yamazaki, M.; Shibata, T.; Nirei, K.; Takahashi, H.; Kaneko, T.; Fujisawa, M.; Higuchi, T.; Nakamura, H.; et al. Apoptosis and non-alcoholic fatty liver diseases. World J. Gastroenterol. 2018, 24, 2661. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Li, C.; Liu, Q.; Wang, A.; Lei, M. Inhibiting ceramide synthesis attenuates hepatic steatosis and fibrosis in rats with non-alcoholic fatty liver disease. Front. Endocrinol. 2019, 10, 665. [Google Scholar] [CrossRef]
- Wree, A.; Mehal, W.Z.; Feldstein, A.E. Targeting Cell Death and Sterile Inflammation Loop for the Treatment of Nonalcoholic Steatohepatitis. Semin. Liver Dis. 2016, 36, 27–36. [Google Scholar] [CrossRef]
- Feng, Y.Y.; Xu, X.Q.; Ji, C.B.; Shi, C.M.; Guo, X.R.; Fu, J.F. Aberrant hepatic MicroRNA expression in nonalcoholic fatty liver disease. Cell. Physiol. Biochem. 2014, 34, 1983–1997. [Google Scholar] [CrossRef]
- Vulf, M.; Shunkina, D.; Komar, A.; Bograya, M.; Zatolokin, P.; Kirienkova, E.; Gazatova, N.; Kozlov, I.; Litvinova, L. Analysis of miRNAs Profiles in Serum of Patients with Steatosis and Steatohepatitis. Front. Cell Dev. Biol. 2021, 9, 2398. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Peng, J.; Luan, Z.; Zheng, F.; Su, W. MicroRNAs as a novel tool in the diagnosis of liver lipid dysregulation and fatty liver disease. Molecules 2019, 24, 230. [Google Scholar] [CrossRef] [PubMed]
- Shabgah, A.G.; Norouzi, F.; Hedayati-Moghadam, M.; Soleimani, D.; Pahlavani, N.; Navashenaq, J.G. A comprehensive review of long non-coding RNAs in the pathogenesis and development of non-alcoholic fatty liver disease. Nutr. Metab. 2021, 18, 22. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Dou, G.; Wang, L. Micrornas in the pathogenesis of nonalcoholic fatty liver disease. Int. J. Biol. Sci. 2021, 17, 1851. [Google Scholar] [CrossRef]
- Roy, S.; Benz, F.; Luedde, T.; Roderburg, C. The role of miRNAs in the regulation of inflammatory processes during hepatofibrogenesis. Hepatobiliary Surg. Nutr. 2015, 4, 24. [Google Scholar] [CrossRef]
- Kim, Y.A.; Park, K.K.; Lee, S.J. LncRNAs act as a link between chronic liver disease and hepatocellular carcinoma. Int. J. Mol. Sci. 2020, 21, 2883. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and functions of long non-coding RNAs at multiple regulatory levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Park, S.; Lee, K.; Jang, D.S.; Kim, S.K. 5-HT1A receptors mediate the analgesic effect of rosavin in a mouse model of oxaliplatin-induced peripheral neuropathic pain. Korean J. Physiol. Pharmacol. 2021, 25, 489–494. [Google Scholar] [CrossRef]
- Bedossa, P.; Burt, A.A.; Gouw, A.H.A.; Lackner, C.; Schirmacher, P.; Terracciano, L.; Tiniakos, D.; Brain, J.; Bury, Y.; Cabibi, D.; et al. Utility and appropriateness of the fatty liver inhibition of progression (FLIP) algorithm and steatosis, activity, and fibrosis (SAF) score in the evaluation of biopsies of nonalcoholic fatty liver disease. Hepatology 2014, 60, 565–575. [Google Scholar] [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Chen, B.; Ma, Y.; Xue, X.; Wei, J.; Hu, G.; Lin, Y. Tetramethylpyrazine reduces inflammation in the livers of mice fed a high fat diet. Mol. Med. Rep. 2019, 19, 2561–2568. [Google Scholar] [CrossRef]
- Matboli, M.; Hasanin, A.H.; Hamady, S.; Khairy, E.; Mohamed, R.H.; Aboul-Ela, Y.M.; Raafat, M.H.; Elsebay, S.A.G.; Emam, H.Y.; Shamekh, R.S.; et al. Anti-inflammatory effect of trans-anethol in a rat model of myocardial ischemia-reperfusion injury. Biomed. Pharmacother. 2022, 150, 113070. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Klebl, D.P.; Feasey, M.C.; Hesketh, E.L.; Ranson, N.A.; Wurdak, H.; Sobott, F.; Bon, R.S.; Muench, S.P. Cryo-EM structure of human mitochondrial HSPD1. iScience 2021, 24, 102022. [Google Scholar] [CrossRef]
- Nagae, M.; Re, S.; Mihara, E.; Nogi, T.; Sugita, Y.; Takagi, J. Crystal structure of α5β1 integrin ectodomain: Atomic details of the fibronectin receptor. J. Cell Biol. 2012, 197, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, J.D.; Longenecker, K.L.; Wilson, N.S.; Goess, C.; Panchal, S.C.; Swann, S.L.; Petros, A.M.; Hobson, A.D.; Ihle, D.; Song, D.; et al. Development of Orally Efficacious Allosteric Inhibitors of TNFα via Fragment-Based Drug Design. J. Med. Chem. 2021, 65, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Tochowicz, A.; Goettig, P.; Evans, R.; Visse, R.; Shitomi, Y.; Palmisano, R.; Ito, N.; Richter, K.; Maskos, K.; Franke, D.; et al. The dimer interface of the membrane type 1 matrix metalloproteinase hemopexin domain: Crystal structure and biological functions. J. Biol. Chem. 2011, 286, 7587–7600. [Google Scholar] [CrossRef]
- Patov, S.A.; Punegov, V.V.; Kuchin, A.V. Synthesis of the Rhodiola rosea glycoside rosavin. Chem. Nat. Compd. 2006, 42, 397–399. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Linares, I.; Pérez-Sánchez, H.; Cecilia, J.M.; García, J.M. High-Throughput parallel blind Virtual Screening using BINDSURF. BMC Bioinform. 2012, 13, S13. [Google Scholar] [CrossRef]
- Trott, O.; Arthur, J. Olson AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the scope of the protein-ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- Popenda, M.; Szachniuk, M.; Antczak, M.; Purzycka, K.J.; Lukasiak, P.; Bartol, N.; Blazewicz, J.; Adamiak, R.W. Automated 3D structure composition for large RNAs. Nucleic Acids Res. 2012, 40, e112. [Google Scholar] [CrossRef]
- Antczak, M.; Popenda, M.; Zok, T.; Sarzynska, J.; Ratajczak, T.; Tomczyk, K.; Adamiak, R.W.; Szachniuk, M. New functionality of RNAComposer: An application to shape the axis of miR160 precursor structure. Acta Biochim. Pol. 2016, 63, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.Z.; Jiang, Y.; Zhou, Y.; Chen, S.J. RLDOCK: A New Method for Predicting RNA-Ligand Interactions. J. Chem. Theory Comput. 2020, 16, 7173–7183. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhao, J.; Hu, Y.; Peng, J. Targeting programmed cell death in metabolic dysfunction-associated fatty liver disease (MAFLD): A promising new therapy. Cell. Mol. Biol. Lett. 2021, 26, 17. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NC | NASH | RSV-10 | RSV-20 | RSV-30 | |
|---|---|---|---|---|---|
| AST (IU/L) | 21.17 ± 6.43 | 149.7 ± 36.49 * | 88.17 ± 10.68 # | 57.67 ± 5.78 #a | 36.17 ± 5.04 #a |
| ALT (IU/L) | 10.1 ± 3.62 | 179.2 ± 38.86 * | 83.05 ± 7.85 # | 45 ± 12.31 #a | 20.83 ± 6.11 #a |
| T. bilirubin (mg/dL) | 0.3967 ± 0.16 | 3.767 ± 0.47 * | 2.65 ± 0.27 # | 1.635 ± 0.14 #a | 0.715 ± 0.19 #ab |
| D. bilirubin (mg/dL) | 0.2133 ± 0.08 | 2.567 ± 0.42 * | 1.7 ± 0.33 # | 1.042 ± 0.15 #a | 0.495 ± 0.16 #ab |
| Albumin (g/dL) | 3.67 ± 0.37 | 1.68 ± 0.28 * | 2.03 ± 0.25 | 2.60 ± 0.17 #a | 3.13 ± 0.49 #a |
| AFP (ng/mL) | 19.65 ± 3.43 | 540.2 ± 108.2 * | 221.7 ± 32.16 # | 92.75 ± 20.5 #a | 29 ± 7.13 #a |
| TG (mg/dL) | 45.67 ± 4.89 | 173.7 ± 19.93 * | 104 ± 22.09 # | 73.83 ± 11.44 #a | 55.17 ± 5.19 #a |
| TC (mg/dL) | 106.6 ± 13.92 | 228.1 ± 8.81 * | 195.9 ± 7.90 # | 171.7 ± 6.93 #a | 118.2 ± 18.75 #ab |
| HDL-C (mg/dL) | 53.33 ± 3.72 | 21.67 ± 3.44 * | 33.33 ± 1.97 # | 38.17 ± 1.17 # | 45.5 ±3.51 #ab |
| LDL-C (mg/dL) | 40.17 ± 10.5 | 171.7 ± 7.61 * | 144.3 ± 8.48 # | 114.3 ± 12.14 #a | 72.17 ± 9.54 #ab |
| NC | NASH | RSV-10 | RSV-20 | RSV-30 | |
|---|---|---|---|---|---|
| Caspase-3 | 1.7 ± 0.3 | 45.3 ± 6.6 * | 12.2 ± 3.8 *# | 9.5 ± 2.4 *# | 2.4 ± 0.8 # ab |
| TNF-α | 0.9 ± 0.2 | 39.7 ±7.1 * | 11.4 ± 3.8 *# | 9.3 ± 2.9 *# | 1.4 ± 0.9 # ab |
| IL-6 | 0.6 ± 0.1 | 44 ± 4.9 * | 10.1 ± 4.6 *# | 8.2 ± 3.4 *# | 0.9 ± 0.3 # ab |
| Caspase-3 | TNF-α | IL-6 | HSPD1 | TNF | MMP14 | ITGB1 | miR-6881-5P | Lnc-SPARCL1-1:2 | |
|---|---|---|---|---|---|---|---|---|---|
| Caspase-3 | 1 | 0.95 * | 0.97 * | 0.74 * | 0.8 * | 0.77 * | 0.89 * | 0.79 * | 0.93 * |
| TNF-α | 0.95 * | 1 | 0.97 * | 0.75 * | 0.77 * | 0.77 * | 0.9 * | 0.77 * | 0.9 * |
| IL-6 | 0.97 * | 0.97 * | 1 | 0.71 * | 0.78 * | 0.77 * | 0.88 * | 0.73 * | 0.89 * |
| HSPD1 | 0.74 * | 0.75 * | 0.71 * | 1 | 0.68 * | 0.73 * | 0.88 * | 0.8 * | 0.75 * |
| MMP14 | 0.77 * | 0.77 * | 0.77 * | 0.73 * | 0.78 * | 1 | 0.77 * | 0.85 * | 0.85 * |
| ITGB1 | 0.89 * | 0.9 * | 0.88 * | 0.88 * | 0.79 * | 0.77 * | 1 | 0.81 * | 0.89 * |
| TNF | 0.8 * | 0.77 * | 0.78 * | 0.68 * | 1 | 0.78 * | 0.79 * | 0.65 * | 0.8 * |
| miR-6881–5P | 0.79 * | 0.77 * | 0.73 * | 0.8 * | 0.65 * | 0.85 * | 0.81 * | 1 | 0.87 * |
| Lnc-SPARCL1-1:2 | 0.93 * | 0.9 * | 0.89 * | 0.75 * | 0.8 * | 0.85 * | 0.89 * | 0.87 * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albadawy, R.; Hasanin, A.H.; Agwa, S.H.A.; Hamady, S.; Aboul-Ela, Y.M.; Raafat, M.H.; Kamar, S.S.; Othman, M.; Yahia, Y.A.; Matboli, M. Rosavin Ameliorates Hepatic Inflammation and Fibrosis in the NASH Rat Model via Targeting Hepatic Cell Death. Int. J. Mol. Sci. 2022, 23, 10148. https://doi.org/10.3390/ijms231710148
Albadawy R, Hasanin AH, Agwa SHA, Hamady S, Aboul-Ela YM, Raafat MH, Kamar SS, Othman M, Yahia YA, Matboli M. Rosavin Ameliorates Hepatic Inflammation and Fibrosis in the NASH Rat Model via Targeting Hepatic Cell Death. International Journal of Molecular Sciences. 2022; 23(17):10148. https://doi.org/10.3390/ijms231710148
Chicago/Turabian StyleAlbadawy, Reda, Amany Helmy Hasanin, Sara H. A. Agwa, Shaimaa Hamady, Yasmin M. Aboul-Ela, Mona Hussien Raafat, Samaa Samir Kamar, Mohamed Othman, Yahia A. Yahia, and Marwa Matboli. 2022. "Rosavin Ameliorates Hepatic Inflammation and Fibrosis in the NASH Rat Model via Targeting Hepatic Cell Death" International Journal of Molecular Sciences 23, no. 17: 10148. https://doi.org/10.3390/ijms231710148
APA StyleAlbadawy, R., Hasanin, A. H., Agwa, S. H. A., Hamady, S., Aboul-Ela, Y. M., Raafat, M. H., Kamar, S. S., Othman, M., Yahia, Y. A., & Matboli, M. (2022). Rosavin Ameliorates Hepatic Inflammation and Fibrosis in the NASH Rat Model via Targeting Hepatic Cell Death. International Journal of Molecular Sciences, 23(17), 10148. https://doi.org/10.3390/ijms231710148