Abstract
Soy isoflavones are phytochemicals that possess various beneficial physiological properties such as anti-aging, anti-tumor, and antioxidant properties. Since soy isoflavones exist in glycoside forms, their bioavailability requires initial hydrolysis of the sugar moieties bound to them to be efficiently absorbed through the gut epithelium. Instead of conventional chemical hydrolysis using acids or organic solvents, alternative strategies for enhancing the bioavailability of soy isoflavones using biological methods are gaining attention. Here, we engineered Leuconostoc citreum isolated from Korean kimchi for efficient bioconversion of soy isoflavone glycosides into their aglycone forms to enhance their bioavailability. We first constructed an expression module based on the isoflavone hydrolase (IH)-encoding gene of Bifidobacterium lactis, which mediates conversion of isoflavone glycosides to aglycone forms. Using a high copy number plasmid and bicistronic expression design, the IH was successfully synthesized in L. citreum. Additionally, we determined enzymatic activity of the IH using an in vivo β-glucosidase assay and confirmed its highly efficient bioconversion efficiency for various types of isoflavone glycosides. Finally, we successfully demonstrated that the engineered L. citreum could convert isoflavone glycosides present in fermented soymilk into aglycones.
1. Introduction
Edible plants are dietary sources of numerous phytochemicals, which are typically secondary metabolites that occur in plant-derived foods. Among the various classes of phytochemicals, polyphenols, which include phenolic acids, flavonoids, and stilbenes are the major classes of bioactive phytochemicals [1]. One of the important physiological properties of these molecules is their superior ability to promote human health and reduce oxidative stress in mammals [1]. Soy isoflavones present in various plants, particularly in soybeans, are classified as flavonoids with a structure similar to estrogen and possess both estrogen-agonist and estrogen-antagonist properties [1,2]. In recent years, the potential health benefits of soy isoflavones have been revealed to include numerous physiological properties such as anti-aging and anti-tumor properties [2,3]. Furthermore, soy isoflavones have the ability to improve learning and memory and also aid in preventing heart diseases, diabetes, and Kawasaki disease [2]. Isoflavones also possess protective antioxidant properties that help prevent oxidative stress by reducing the generation of free radicals and reactive oxygen species (ROS) via decomposition of hydrogen peroxide and quenching of singlet oxygen [1,4]. In their natural environment, soy isoflavones exist in glycoside forms (genistin, daidzin, and glycitin) which bind to sugar molecules. However, to be absorbed in mammals, the bound sugar molecules need to be released, thus leading to the formation of aglycones such as genistein, daidzein, and glycitein [5]. In soybeans, since most isoflavones exist in glycoside forms, their bioavailability requires initial hydrolysis of the sugar moieties by isoflavone hydrolases produced by commensal bacteria present in the gut, because mammals do not possess intrinsic isoflavone hydrolases [6,7]. In this regard, to enhance the bioactivity and bioavailability of soy isoflavones, various strategies based on chemical methods, including hydrolysis of isoflavone glycosides into aglycone forms using acid or organic solvents, have been developed for the extraction of isoflavones [8,9,10]. However, these methods are energy-intensive, and byproducts produced during the extraction can result in environmental pollution and safety issues for downstream applications. Consequently, alternative strategies that use biological methods relying on microorganisms or immobilized enzyme systems to enhance the bioavailability of soy isoflavones are gaining attention [11,12].
Lactic acid bacteria (LAB) are a well-known group of non-sporulating gram-positive bacteria composed of several different genera, including Lactococcus, Lactobacillus, Leuconostoc, Pediococcus, Streptococcus, and Oenococcus [13]. Historically, they have long been used in the manufacture of fermented foods such as milk and vegetables to enhance taste and natural preservation [14]. In recent years, LAB are drawing increasing attention for novel purposes, owing to their safe use for human and animal consumption, metabolic versatility, and a wide ecological niche [15]. Furthermore, beneficial properties such as simple carbon metabolism, effective energy consumption, and a small genome make LAB crucial candidates for metabolic engineering strategies that are focused on regulating synthetic pathways to produce important fermentation end-products including sweeteners, flavors, and vitamins [16]. Considering these beneficial characteristics, LAB have been used along with Bifidobacterium as an excellent probiotic for bioconversion of soy isoflavone glycosides [17,18]. Nevertheless, the constraints of anaerobic conditions required for their growth and low conversion efficiency limit their utilization for enhancing the bioavailability of soy isoflavones.
Leuconostoc citreum is a hetero-fermentative LAB that plays a significant role in the fermentation of dairy products such as kimchi, milk, vegetables, and meat [19]. Recently, L. citreum has been identified as a crucial probiotic candidate to produce vitamins and bioactive antimicrobial peptides, as well as to regulate immune responses [20,21,22]. Currently, L. citreum has been attracting attention for its use in food biotechnology owing to its favorable advantages such as non-production of endotoxins and a good growth rate under both aerobic and anaerobic conditions [23]. In addition, recent advances in genetic engineering tools related to L. citreum have enabled its application as a potential probiotic strain [24,25,26].
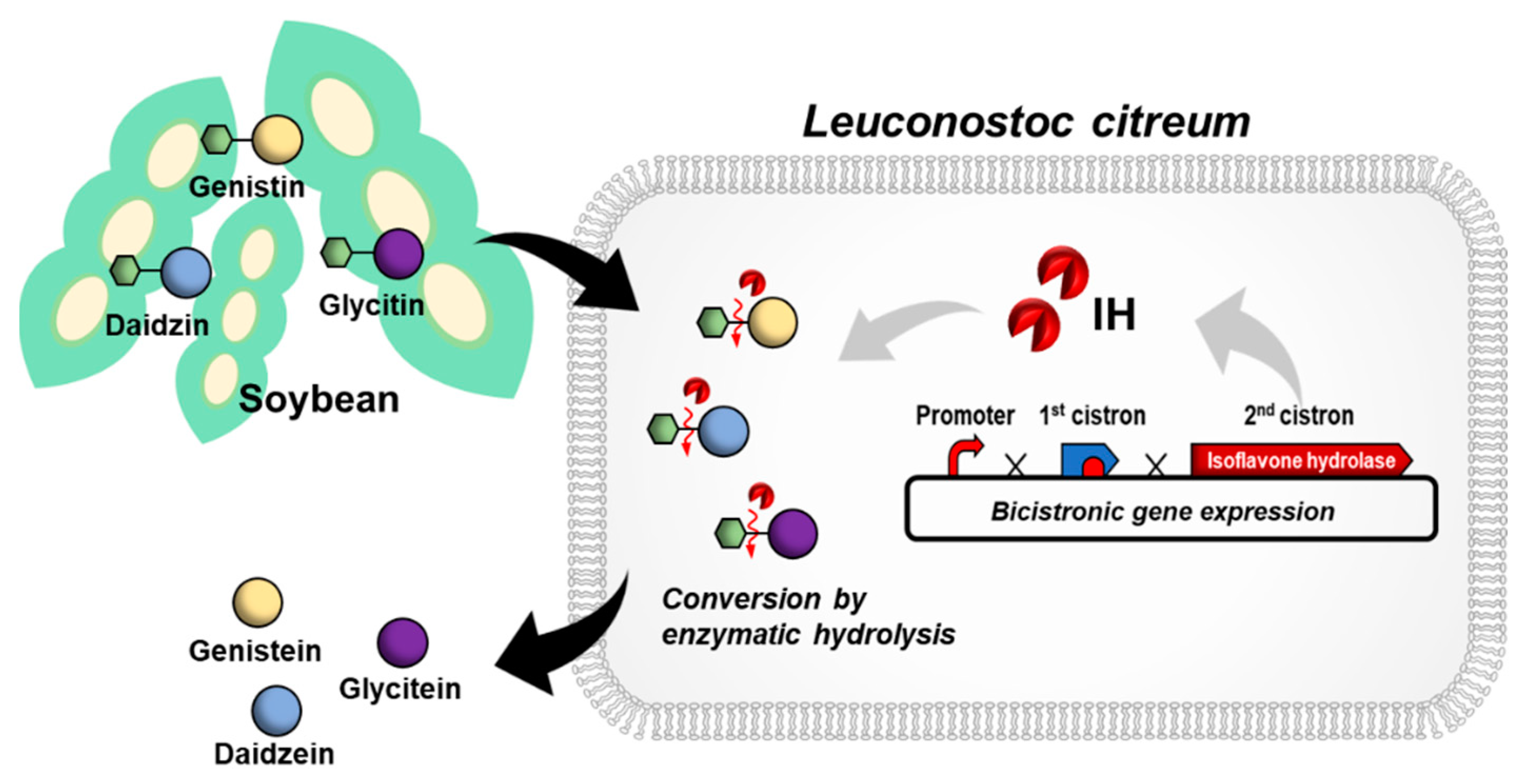
In this study, we report the engineering of L. citreum for efficient bioconversion of soybean isoflavone glycosides (Figure 1). β-glucosidase is known as the enzyme that is responsible for the hydrolytic breakdown of carbohydrates such as cellobiose [27], and it can be classified as isoflavone hydrolase with high specificity towards isoflavone glycosidic conjugates. Therefore, an expression module based on the isoflavone hydrolase encoding gene of Bifidobacterium lactis, which mediates the conversion of isoflavone glycosides to aglycone forms, was constructed in L. citreum. Using a high copy number plasmid (pCB4270) and bicistronic expression design, the isoflavone hydrolase from B. lactis (IH) was successfully synthesized in L. citreum, and enzymatic activity of the IH was determined using an in vivo β-glucosidase assay. Naturally, there are several types of soy isoflavone glycosides including malonyl, acetyl, and underivatized glucosides, and their content differs depending on product type and processing conditions [2,28]. Among them, the major isoflavones in soybeans are underivatized glucosides such as daidzin, genistin, and glycitin [28,29,30], so we focused on the bioconversion of underivatized glucosides (genistin, daidzin, and glycitin) and confirmed its highly efficient bioconversion efficiency for all those glucosides to their aglycone forms. Finally, using engineered L. citreum, we demonstrated that isoflavone glycosides produced during fermentation of soymilk could be converted into aglycones with a high conversion yield.
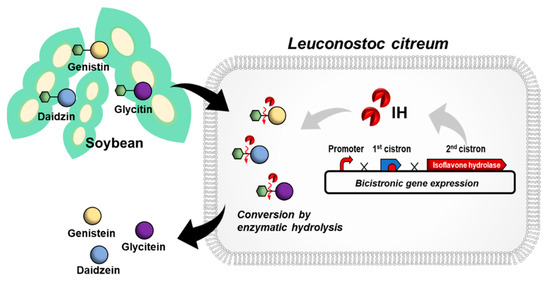
Figure 1.
Schematic of L. citreum engineering for the efficient bioconversion of soy isoflavone glycoside forms to aglycone forms.
2. Results and Discussion
2.1. Expression of the IH Gene in L. citreum for Hydrolysis of Isoflavone Glycosides
To engineer L. citreum that is capable of hydrolyzing isoflavone glycosides, it is necessary to develop a glycoside hydrolysis system using a highly specific and efficacious enzyme. Previously, it was reported that LAB, including Lactobacillus, Enterococcus, Lactococcus, and Bifidobacterium, have a great potential in hydrolyzing glycosides into aglycones [7]. Youn et al. (2012) isolated a novel isoflavone hydrolase (IH) from Bifidobacterium lactis SH5 that showed superior hydrolytic activity against isoflavone glycoside forms [31]. Furthermore, kinetic analysis of this IH revealed that the optimal pH and temperature (30 °C) required for its stability were also suitable for the growth of L. citreum [31]. Therefore, we decided to employ IH from B. lactis SH5 for efficient production of isoflavone aglycone forms in L. citreum.
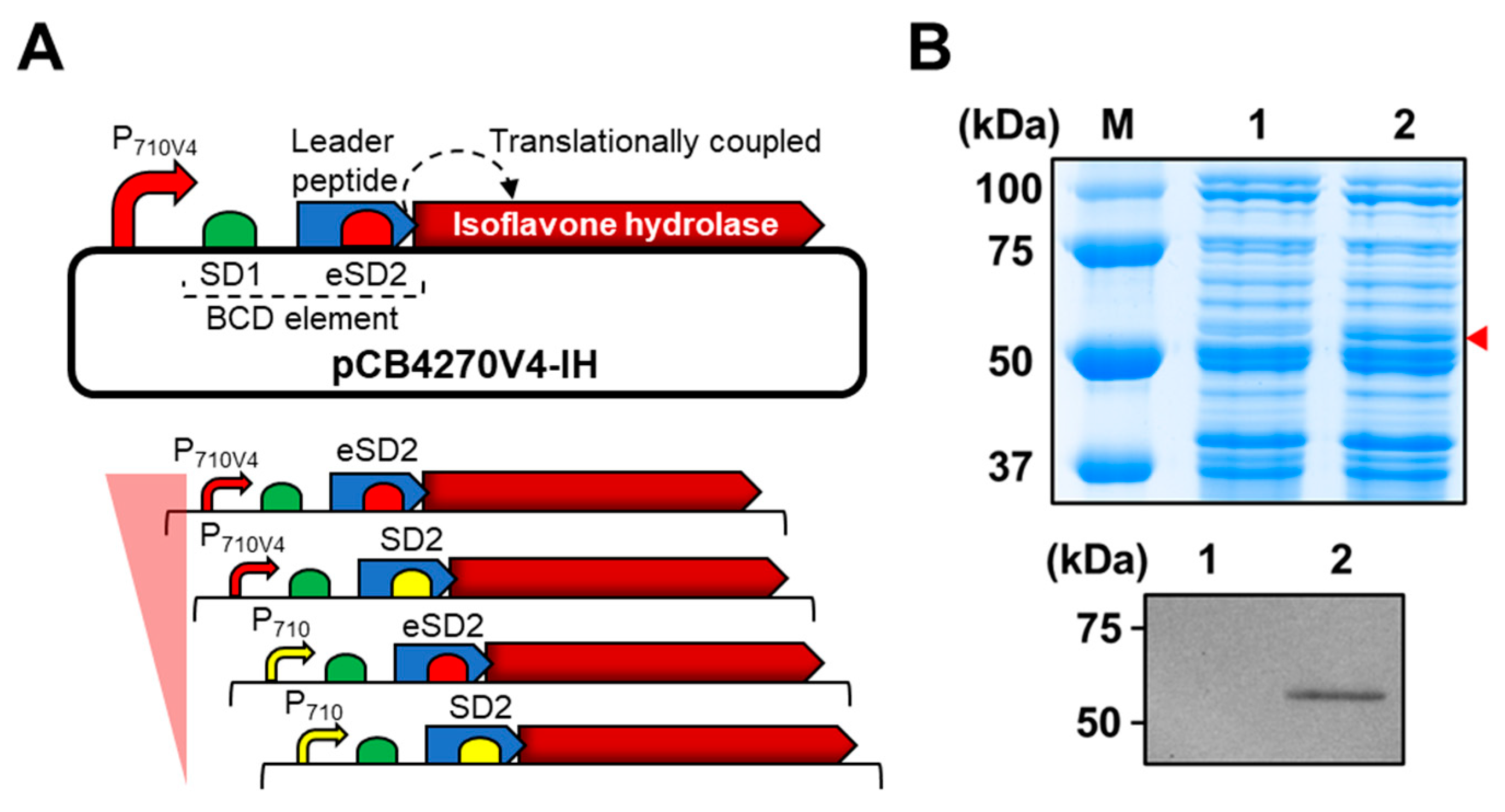
It is well known that establishing a highly efficient whole-cell biocatalyst is correlated with the content of a particular key enzyme in a cell; therefore, we used the high copy number plasmid pCB4270 [26] to clone the IH-coding gene under the control of bicistronic expression design, which was developed for stable expression of heterologous proteins in L. citreum [25] (Figure 2A). In the bicistronic design, the first cistron consisting of the Shine–Dalgarno (SD) sequence 1 (SD1) and a leader peptide (17 amino acids) was positioned between the promoter and IH gene (Figure 2A) to reliably express the target gene in L. citreum. As a second SD sequence, SD2 was cloned at the 3′-end of a leader peptide sequence; it serves as a ribosome binding site (RBS) for translation of the IH gene (second cistron) by means of translational coupling (Figure 2A). Four sets of variants with a bicistronic design that enabled the expression of the gene encoding for IH at various intensities based on two different strength promoters (strong P710V4 and weak P710) and SD sequences (strong eSD2 and weak SD2) were constructed (Figure 2A), and expression level of the IH protein in each variant was analyzed by SDS-PAGE. Among the four sets of variants, we found that the construct, consisting of a strong constitutive promoter (P710V4) and strong SD sequence (eSD2), resulted in higher expression levels of heterologous protein and induced successful expression of the IH protein in L. citreum (Figure 2B). Therefore, we decided to use this strong expression system for subsequent conversion reactions in L. citreum.
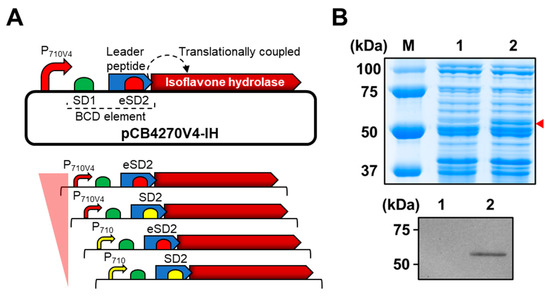
Figure 2.
Expression of the gene encoding for IH in L. citreum for bioconversion of soy isoflavone glycosides. (A) Schematic diagram of bicistronic design (BCD) for expression of the gene encoding for IH. Up, plasmid pCB4270V4-IH constructed for the expression of the gene encoding for IH in L. citreum; down, four different expression systems with different strengths of promoters (P710 and P710V4) and SD (SD2 and eSD2) parts. (B) SDS-PAGE and western blot analysis of IH gene expression in L. citreum harboring pCB4270V4-IH (P710V4 and eSD2). Up, SDS-PAGE analysis. Lane M, molecular weight markers (kDa); Lane 1, soluble fractions of L. citreum wild type; Lane 2, soluble fractions of L. citreum harboring pCB4270V4-IH. Arrowhead indicates the band of IH; down, confirmation of IH by western blot assay. Lane 1, L. citreum wild type; Lane 2, L. citreum harboring pCB4270V4-IH.
2.2. Evaluation of Hydrolysis of Isoflavone Glycosides with Engineered L. citreum
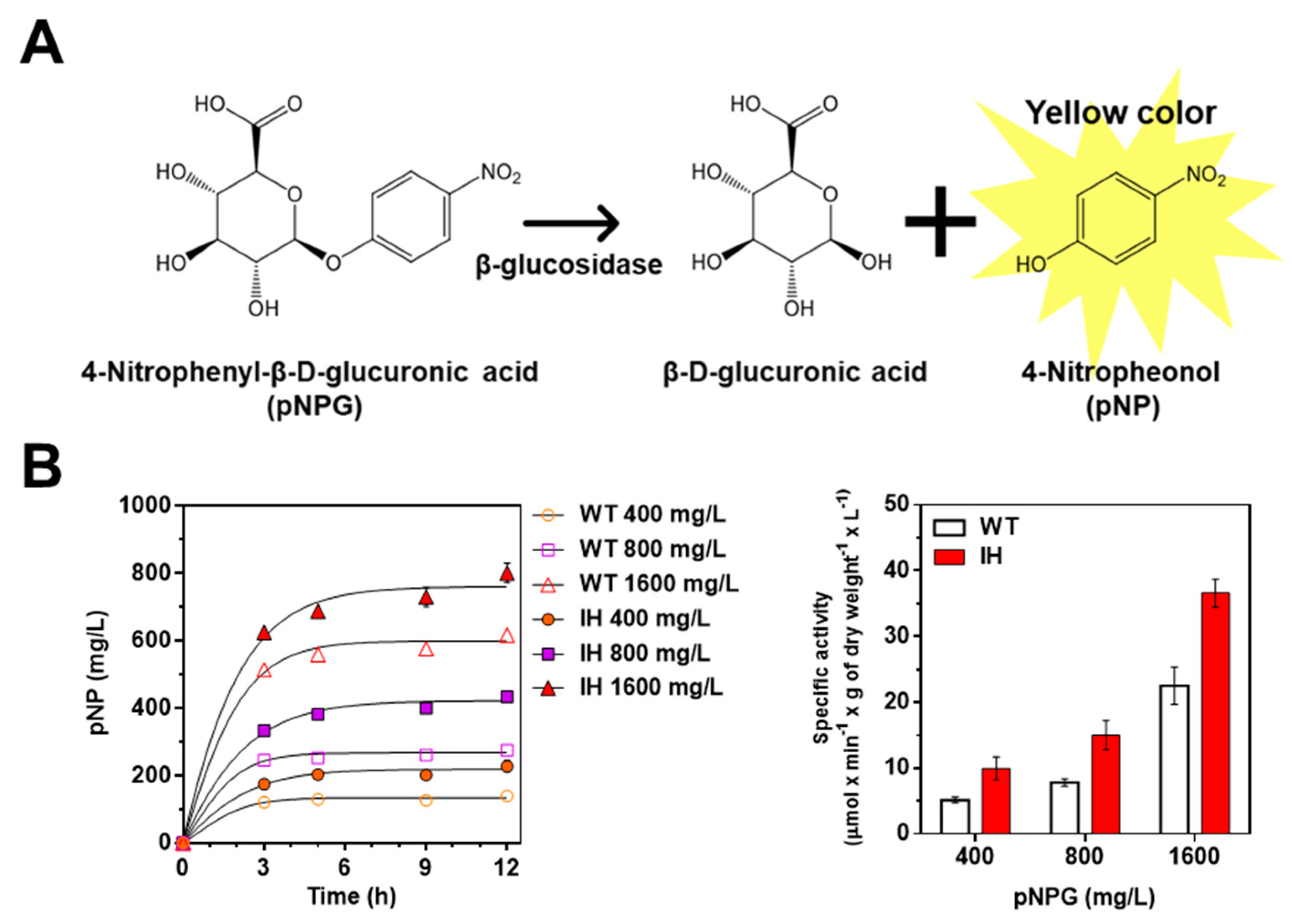
After we confirmed that the IH gene was properly expressed in L. citreum harboring pCB4270V4-IH, an in vivo β-glucosidase activity assay was performed to assess whether the synthesized IH was capable of being enzymatically active in L. citreum. The β-glucosidase activity was determined by bioconversion of 4-nitrophenyl-β-d-glucopyranoside (pNPG) into 4-nitrophenol (pNP) during bacterial cultivation (Figure 3A). The enzymatic product pNP was measured directly from the supernatant of the culture medium at an absorbance of 405 nm. Since MRS medium also displays absorbance at the same wavelength (405 nm), 0.1 M sodium carbonate was added to the supernatant to increase the molar extinction coefficient of pNP [32].
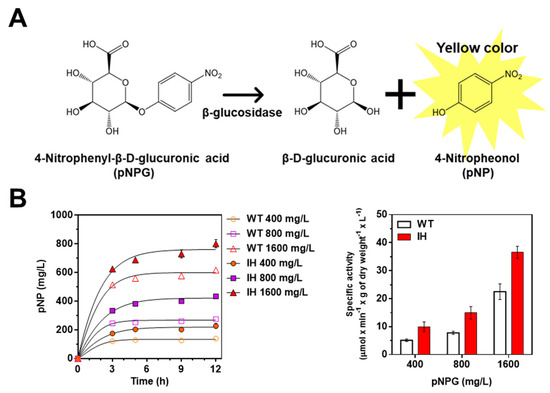
Figure 3.
Identification of enzymatic activity of IH via in vivo β-glucosidase activity assay. (A) Schematic of reaction mechanism of β-glucosidase assay. (B) β-glucosidase activity of L. citreum wild type and L. citreum harboring pCB4270V4-IH in MRS medium supplemented with different concentrations of pNPG. The pNP production was measured at different culture times. Left, time profiles of pNP production; right, normalized specific activity of β-glucosidase activity of L. citreum wild type and L. citreum harboring pCB4270V4-IH at 12 h. WT represents L. citreum wild type and IH represents L. citreum harboring pCB4270V4-IH. Results are the mean of duplicate experiments, and error bars indicate standard deviations.
As expected, enzymatic activity of β-glucosidase was observed in both L. citreum wild type and L. citreum harboring pCB4270V4-IH (Figure 3B and Table 1) with various concentrations of pNPG (400, 800, and 1600 mg/L). As shown in Figure 3B, we found that L. citreum wild type also showed enzymatic activity for β-glucosidase, and it was assumed that the intrinsic β-glucosidases, due to their requirement in the metabolism of diverse carbon sources such as cellulose and lactose [33] in L. citreum, showed hydrolytic activity for pNPG regardless of their substrate specificity for isoflavone glycoside forms. Meanwhile, taking into consideration normalized specific activity of β-glucosidase, L. citreum harboring pCB4270V4-IH showed approximately two-fold higher activity than L. citreum wild type (Figure 3B and Table 1). This result indicated that the expressed IH in L. citreum harboring pCB4270V4-IH was biologically active in L. citreum.

Table 1.
Summary of in vivo β-glucosidase activity in L. citreum harboring pCB4270V4-IH.
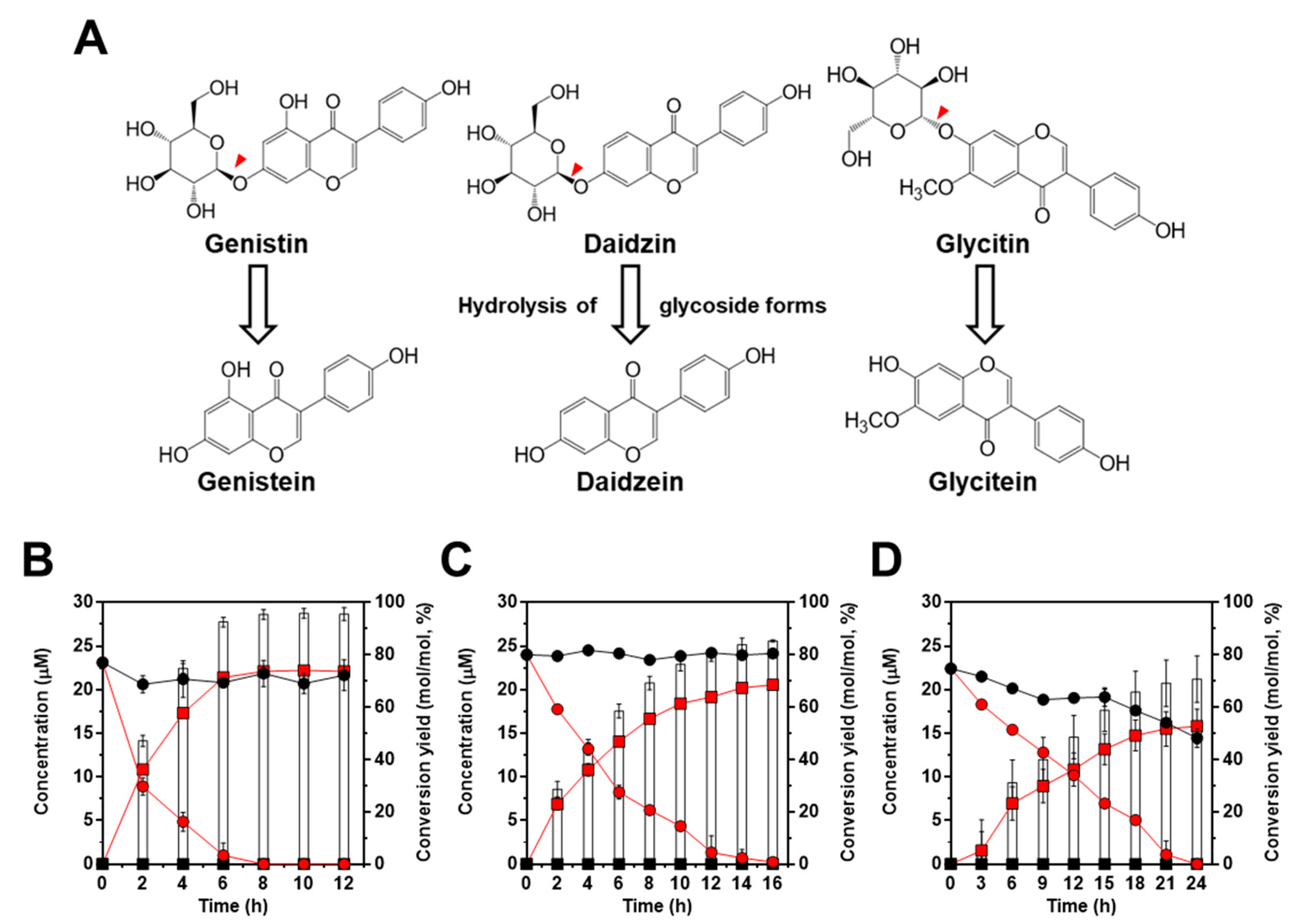
After we confirmed the enzymatic activity of IH using the β-glucosidase activity assay, bioconversion of isoflavone glycoside into aglycone forms was evaluated using three isoflavone glycosides (genistin, daidzin, and glycitin) which are major isoflavones in soybean (Figure 4A). As shown in Figure 4B–D, for genistin, daidzin, and glycitin, we found a high conversion yield of glycoside into aglycone forms in L. citreum harboring pCB4270V4-IH. First, contrary to the result of β-glucosidase activity observed in L. citreum wild type, bioconversion of isoflavone was not observed in wild type strain (Figure 4B–D). Therefore, it was confirmed that intrinsic β-glucosidases in L. citreum CB2567 did not have substrate specificity for isoflavone glycoside forms. Meanwhile, L. citreum harboring pCB4270V4-IH showed the highest activity for genistin among three examined substrates: 22.1 µM of genistein (aglycone form) was produced from 23.1 µM of genistin (100 mg/L) (glycoside form), with a conversion rate of 2.7 µM/h and a yield of 95% (0.95 mol/mol) at 8 h in L. citreum harboring pCB4270V4-IH (Figure 4B). Next, for 24 µM of daidzin (100 mg/L) (glycoside form), 20.5 µM of daidzein (aglycone form) was produced, with a conversion rate of 1.2 µM/h and a yield of 85% (0.85 mol/mol) at 16 h (Figure 4C). Finally, in the case of glycitin substrate, it was found that 15.8 µM of glycitein (aglycone form) was produced from 22.4 µM of glycitin (100 mg/L), with a conversion rate of 0.6 µM/h and a yield of 70% (0.70 mol/mol) at 24 h (Figure 4D). These results clearly confirmed that the engineered L. citreum had a highly efficient bioconversion property to convert all the isoflavone glycoside forms into aglycone forms.
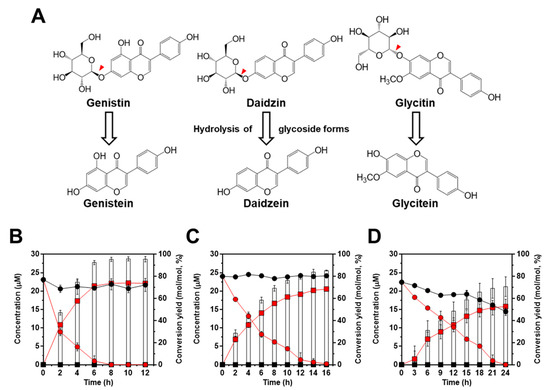
Figure 4.
Bioconversion of isoflavones with standard chemicals by L. citreum wild type and L. citreum harboring pCB4270V4-IH. (A) Schematic diagram of bioconversion of isoflavone glycosides to aglycone forms. Red-color arrowheads indicate the hydrolysis site for IH. Bioconversion profiles of (B) 23.1 µM of genistin (100 mg/L) (C) 24 µM of daidzin (100 mg/L) and (D) 22.4 µM of glycitin (100 mg/L). Symbols: black circle, isoflavone glycoside forms in L. citreum wild type; black square, isoflavone aglycone forms in L. citreum wild type; red circle, isoflavone glycoside forms in L. citreum harboring pCB4270V4-IH; red square, isoflavone aglycone forms in L. citreum harboring pCB4270V4-IH, white bar, conversion yields of isoflavones in L. citreum harboring pCB4270V4-IH. Results are the mean of duplicate experiments, and error bars indicate standard deviations.
2.3. Hydrolysis of Isoflavone Glycosides in Fermented Soymilk by Engineered L. citreum
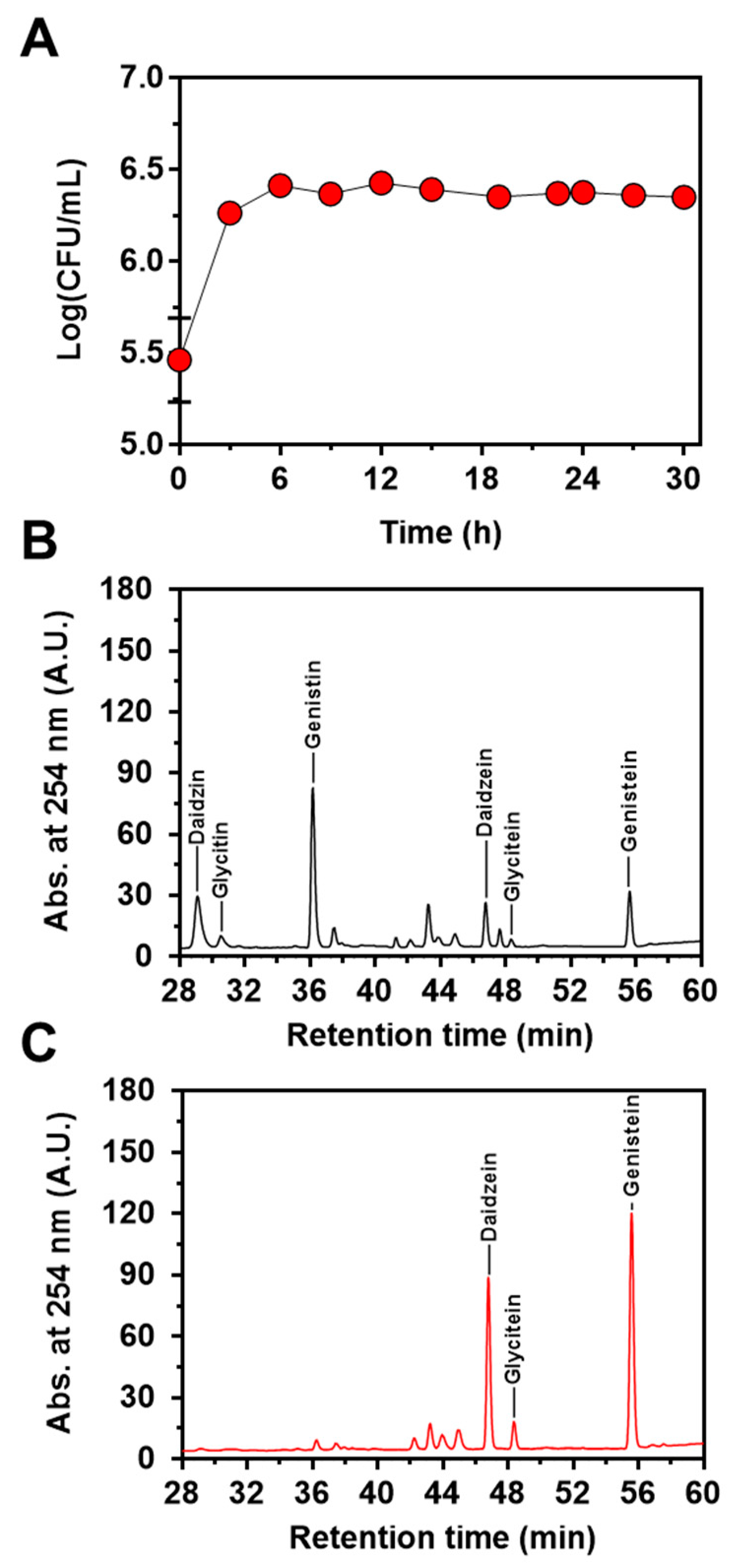
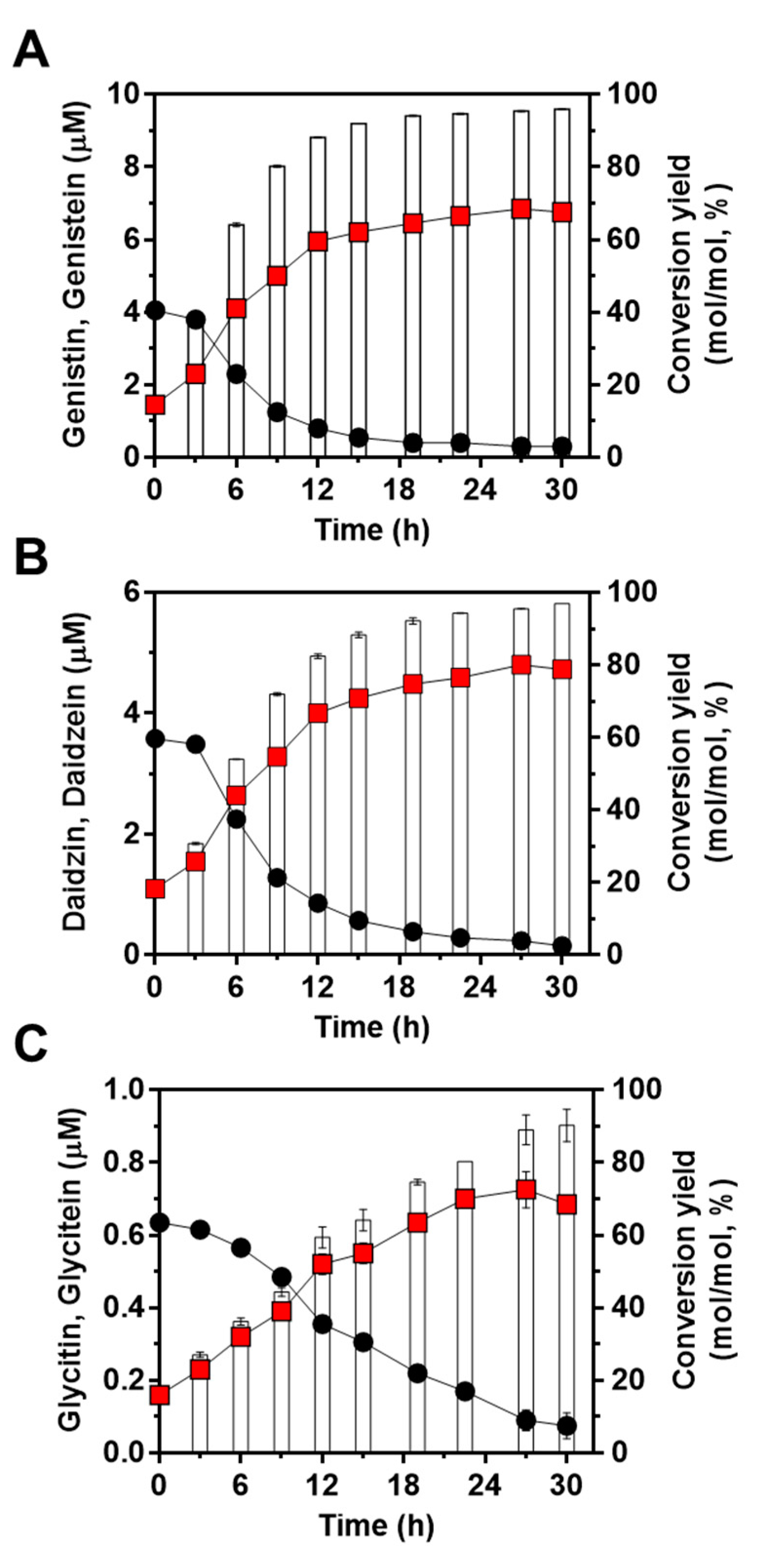
Next, to confirm the feasibility of the engineered strain of L. citreum to convert soy isoflavones during soymilk fermentation, cells were cultivated in 4% soymilk-containing media, and the concentrations of the three isoflavones (genistin, daidzin, and glycitin) and their aglycons (genistein, daidzein, and glycitein) were determined during 30 h of fermentation. As shown in Figure 5A, L. citreum harboring pCB4270V4-IH showed a constant growth rate until 6 h of fermentation and reached a stationary phase during the fermentation process. All the soy isoflavones, including genistin, daidzin, and glycitin, were successfully converted to aglycone forms by L. citreum harboring pCB4270V4-IH during soymilk fermentation (Figure 5B,C). L. citreum harboring pCB4270V4-IH gradually converted 4 µM genistin present in soymilk to 6.8 µM of genistein, with a conversion rate of 0.23 µM/h and a yield of 95% (0.95 mol/mol) (Figure 6A). Next, about 3.5 µM of daidzin present in soymilk was converted to 4.7 µM of daidzein, with a conversion rate of 0.16 µM/h and a yield of 96% (0.96 mol/mol) (Figure 6B). Finally, about 0.63 µM glycitin was converted to 0.68 µM of glycitein, with a conversion rate of 0.02 µM/h and a yield of 90% (0.90 mol/mol) (Figure 6C). All conversion data are summarized in Table 2. These results clearly confirmed that the engineered L. citreum had a highly efficient bioconversion property to convert all the isoflavone glycosides’ forms into aglycone forms, even during fermentation of soymilk.
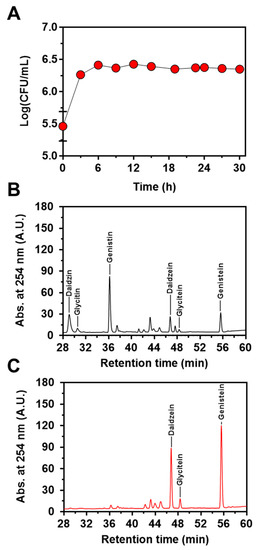
Figure 5.
Soymilk fermentation of L. citreum harboring pCB4270V4-IH. (A) Growth profiles of L. citreum harboring pCB4270V4-IH. Results are the mean of duplicate experiments, and error bars indicate standard deviations. Chromatogram of HPLC analysis of isoflavones in soymilk during soymilk fermentation at (B) 0 h and (C) 30 h.
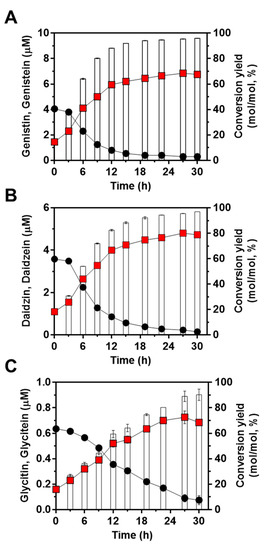
Figure 6.
Bioconversion of isoflavones during soymilk fermentation by L. citreum harboring pCB4270V4-IH. (A) Production profiles of genistein. (B) Production profiles of daidzein. (C) Production profiles of glycitein. Symbols: black circle, isoflavone glycoside forms in soymilk; red square, isoflavone aglycone forms in soymilk; white bar, conversion yields of isoflavones in L. citreum harboring pCB4270V4-IH. Results are the mean of triplicate experiments, and error bars indicate standard deviations.
Table 2.
Summary of isoflavone aglycone forms production with soymilk fermentation.
With advances in synthetic biology and metabolic engineering during the last two decades, various synthetic moieties for gene expression have been developed, which offer innovative approaches for redesigning the existing biological systems in many bacterial hosts, including LABs, rendering them to be increasingly used as cell factories [34,35]. In this study, using a high copy number plasmid (pCB4270) and bicistronic expression design with synthetic parts [24,25,26], an optimized expression module for IH that mediates the conversion of isoflavone glycoside to aglycone forms was successfully constructed in L. citreum. To the best of our knowledge, this is the first report on the engineering of L. citreum to be used as a whole-cell biocatalyst for production of isoflavone aglycones, and our L. citreum-based novel platform has advantages over other bacterial hosts. To date, most of the studies on the hydrolyzation of isoflavone glycoconjugates in fermented soy products have focused on Bifidobacterium strains [36,37,38,39,40,41]. However, constraints regarding the cultivation in anaerobic conditions, lack of useful expression systems, and genome editing tools for Bifidobacterium strains limit their utilization in expanding the bioavailability of soy isoflavones. In addition, most intestinal bacteria including Bifidobacterium strains have metabolic pathways that degrade isoflavone aglycone forms into a variety of metabolites, such as dihydroisoflavones including dihydrogenistein, dihydrodaidzein, and dihydroglycitein [42,43,44], which cause a loss of active aglycone forms. In contrast, in L. citreum, no inherent metabolic pathway for the degradation of isoflavone aglycone forms has been reported. As shown here (Figure 5 and Figure 6), we could not observe the degradation of isoflavone aglycone during hydrolysis with our L. citreum-based whole cell system, so high conversion yields could be achieved without loss of aglycone. In addition, as shown here, various genetic tools are available in L. citreum, which will make this host more potential. It is proven that the engineered L. citreum can be a novel starter culture for soymilk fermentation while not only increasing the content of aglycones but also enhancing both acceptability and nutritional value of fermented soymilk, owing to its intrinsic properties for end products of carbohydrates. Recent scientific advancements clarified the relationship between the diet of soy isoflavones and human health. Particularly, their property for phytoestrogen with a similar structure to 17-β-estradiol allows them to decrease the risk of hormone-related diseases including breast and prostate tumors and even osteoporosis [2,3]. Furthermore, their antioxidant properties contribute to protecting neurodegenerative disease and photoaging effects by insulating ultraviolet (UV) radiation and chemicals [1,4]. In this regard, expectations for the utilization of soy isoflavones are drawing attention more and more, and we believe that our L. citreum-based whole-cell biocatalyst system will contribute to the development of an industrial platform for enhancing the bioavailability of soy isoflavones.
3. Materials and Methods
3.1. Bacterial Strains, Plasmids, and Growth Conditions
The bacterial strains and plasmids used in this study are listed in Table 3. Escherichia coli XL1-Blue was used for gene cloning and plasmid maintenance, and L. citreum CB2567 [45] was used as a major host for bioconversion of isoflavone glycosides. E. coli XL1-Blue was cultivated in Luria-Bertani medium (tryptone 10 g/L, yeast extract 5 g/L, and sodium chloride 10 g/L; BD, Franklin Lakes, NJ, USA) at 37 °C and 200 rpm. L. citreum was cultivated in MRS medium (proteose peptone no. 3 10 g/L, beef extract 10 g/L, yeast extract 5 g/L, dextrose 20 g/L, polysorbate 80 1 g/L, ammonium citrate 2 g/L, sodium acetate 5 g/L, magnesium sulfate 0.1 g/L, manganese sulfate 0.05 g/L, and dipotassium phosphate 2 g/L, purchased as premixed media from BD) at 30 °C with shaking at 200 rpm. Ampicillin (100 µg/mL) and chloramphenicol (Cm, 10 µg/mL) were used for the selection and cultivation of E. coli and L. citreum, respectively.
Table 3.
List of strains and plasmids used in this study.
3.2. Plasmid Construction
For gene expression in L. citreum, pCB4270 [24], a high copy number plasmid derivative of pCB42 [46] from L. citreum CB2567, was used as the backbone plasmid. Polymerase chain reaction (PCR) was performed using a C1000TM Thermal Cycler (Bio-Rad, Richmond, CA, USA) and PrimeSTAR HS Polymerase (Takara, Shiga, Japan). Oligonucleotides used for the PCR are listed in Table 4. To express IH, the native IH gene was synthesized by Macrogen (Daejeon, Korea) and amplified by PCR using primers F-IH and R-IH (Table 4). Next, the PCR product was cloned into pCB4270V4 [24] using the Gibson Assembly method employing the Gibson Assembly Master Mix (NEB, Ipswich, MA, USA), which yielded pCB4270V4-IH, in which the IH gene was constitutively expressed under the bicistronic expression design [25]. Subsequently, pCB4270V4-IH was transformed into L. citreum CB2567 by electroporation with a capacitance of 25 µF and voltage of 1.0 kV and 400 Ω using 0.1 cm cuvettes and a MicroPulser (Bio-Rad). Transformed cells were recovered using MRS agar plates containing 10 µg/mL chloramphenicol at 30 °C for 16 h.
Table 4.
Primers used in this study.
3.3. Protein Preparation and Analysis
Following overnight cultivation of the recombinant L. citreum, the cells were transferred to 5 mL of fresh MRS medium in a 50 mL tube at 1:50 dilution and grown at 30 °C with gentle shaking at 200 rpm. To identify the protein expression level of the IH gene, after cultivating for 12 h, the cells were harvested by centrifugation at 13,000 rpm for 5 min at 4 °C and washed twice with phosphate-buffered saline (PBS, containing 135 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4, pH 7.2). Later, a total protein fraction of the cells was prepared by sonication (amplitude 20%, time 7 min, using “5 s on and 3 s off” mode). The samples were then heated at 100 °C for 5 min, and proteins were separated using 12% SDS-PAGE at 250 V for 30 min. The proteins were subsequently transferred to a polyvinylidene fluoride (PVDF) membrane using a Trans-Blot® SD Semi-Dry Transfer Cell (Bio-Rad) for 1.5 h at 70 mA. Next, the membrane was incubated with 5% (w/v) skim milk to block non-specific binding of antibodies and then incubated with Monoclonal ANTI-HIS®-Peroxidase (HRP) antibody (Sigma-Aldrich, St. Louis, MO, USA) diluted 1:5000 in Tris-buffered saline containing 0.1% Tween (TBST; 20 mM Tris, 150 mM NaCl, and 0.1% Tween® 20 detergent). After washing, his-tagged IH (52 kDa) was detected using Amersham ECL Prime Western Blotting Detection Reagent (GE Healthcare, Chicago, IL, USA). Images were captured using the ChemiDoc imaging system (Bio-Rad).
3.4. In Vivo β-Glucosidase Activity Assay
To analyze enzymatic activity of IH in L. citreum, an in vivo β-glucosidase activity assay was performed. β-glucosidase activity was determined using various concentrations of pNPG (Sigma-Aldrich) and pNP (Sigma-Aldrich). L. citreum wild-type cells and L. citreum cells harboring pCB4270V4-IH were inoculated into 5 mL of MRS (BD) medium and cultivated for 16 h at 30 °C with shaking at 200 rpm. Next, they were transferred to 50 mL of MRS medium in 250 mL baffled flasks at 1:100 dilution and grown at 30 °C with shaking at 200 rpm. Cell growth was analyzed by measuring optical density at 600 nm (OD600) using a spectrophotometer (Optizen POP; Mecasys, Daejeon, Korea). After the stationary phase was reached, 10 mL of the cultured cells was harvested by centrifugation at 13,000 rpm for 5 min and washed with PBS. The washed cells were resuspended in 10 mL fresh MRS medium containing various concentrations of pNPG. At each time point, 200 µL of the sample was loaded into 1.5 mL tubes and harvested by centrifugation at 13,000 rpm for 5 min. The supernatant (150 µL) containing pNP as a product was transferred to a 96-well plate (SPL Life Sciences, Pocheon, Korea), and 50 µL of 0.1 M sodium carbonate was added to each well. The pNP concentration was determined at 405 nm using a microplate reader (Tecan, Männedorf, Switzerland).
3.5. Whole-Cell Bioconversion in a Shake Flask
L. citreum cells harboring pCB4270V4-IH were inoculated into MRS (BD) medium. After overnight cultivation, the cells were transferred into a 250 mL baffled flask containing 50 mL of fresh MRS medium at 1:100 dilution and grown at 30 °C with shaking at 200 rpm for 24 h. Next, 10 mL of the cultured cells were harvested by centrifugation at 13,000 rpm for 5 min and washed twice with PBS buffer. The washed cells were resuspended in 10 mL of fresh MRS medium containing isoflavone glycoside forms (genistin, daidzin, and glycitin; Sigma-Aldrich). All the conversion reactions in the flasks were performed at 200 rpm in a shaking incubator at 30 °C. To measure the amount of isoflavone glycoside and aglycone forms, the cells were pelleted by centrifugation at 13,000 rpm for 5 min, and the supernatants thus obtained were filtered using a 0.22 µm syringe filter (Futecs, Daejeon, Korea). The filtered supernatants were diluted in 80% methanol and analyzed by HPLC.
3.6. Soymilk Fermentation
Soymilk (4%, w/v) was prepared by dissolving soybean flour (Sigma-Aldrich) in deionized water and autoclaving at 121 °C for 15 min. L. citreum cells harboring pCB4270V4-IH were inoculated into MRS (BD) medium. After the cells were cultivated for 16 h at 30 °C with shaking at 200 rpm at 30 °C, they were transferred to 250 mL baffled flasks containing 50 mL of MRS medium at 1:100 dilution and grown at 30 °C with shaking (200 rpm) for 24 h. Later, the cells were transferred to 50 mL of 4% soymilk at 5% (v/v) dilution and incubated at 30 °C with shaking (200 rpm) for 30 h. To analyze growth profiles of L. citreum harboring pCB4270V4-IH, 10 µL of the soymilk sample was diluted with PBS buffer and spread onto MRS agar plates with 10 µg/mL chloramphenicol at 30 °C for 24 h. The number of bacteria (CFU/mL) was calculated using the recovered colonies. Extraction of isoflavones from the fermented soymilk was performed in triplicate. Briefly, 1 mL aliquots of the sample were freeze-dried using a lyophilizer (ilShinBioBase, Daejeon, Korea). The lyophilized samples were dissolved in 80% methanol and heated to 60 °C for 2 h to extract the isoflavones. Next, the samples were centrifuged at 13,000 rpm for 5 min, and the supernatants thus obtained were loaded onto HPLC vial to measure the amount of isoflavone using HPLC.
3.7. HPLC Analysis
The HPLC system (Shimadzu, Kyoto, Japan) consisted of a pump (LC-20AD), autosampler (SIL-30AC), column oven (CTO-20 A), and refractive index detector (RID-10 A), and was equipped with a Zorbox Eclipse AAA column (150 × 4.6 mm, 3.5 microns; Agilent Technologies, PA, CA, USA). Samples were fractionated using a binary nonlinear gradient with mobile phase A (0.1% acetic acid in deionized water) and mobile phase B (0.1% acetic acid in acetonitrile). The column temperature was 35 °C, flow rate was 0.5 mL/min, and UV detection wavelength was 254 nm. The percentages of the mobile phases and the time periods were as follows: 95% mobile phase A for 4 min, 65% mobile phase A for 48 min, 15% mobile phase A for 54 min, 100% mobile phase B for 56 min, 100% mobile phase B for 66 min, 95% mobile phase A for 75 min, and 95% mobile phase A for 80 min.
Author Contributions
Conceptualization, K.J.J. and J.S.; methodology, J.S.; data analysis, J.S.; writing—original draft preparation, J.S. and K.J.J.; writing—review and editing, K.J.J.; supervision, K.J.J.; funding acquisition, K.J.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through the Crop Viruses and Pests Response Industry Technology Development Program, funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA) (Grant no. 321109-04-1-HD020) and by the National Research Foundation of Korea (NRF) grant funded by the Ministry of Science and ICT (MSIT) (No. NRF-2020R1A2C2012537).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| LAB | Lactic acid bacteria |
| IH | Isoflavone hydrolase from Bifidobacterium animalis subsp. lactis SH5 |
References
- Voicescu, M.; Hellwig, P.; Meghea, A. Antioxidant activity of phytoestrogen type isoflavones in biomimetic environments. N. J. Chem. 2016, 40, 606–612. [Google Scholar] [CrossRef]
- Wang, Q.; Ge, X.; Tian, X.; Zhang, Y.; Zhang, J.; Zhang, P. Soy isoflavone: The multipurpose phytochemical. Biomed. Rep. 2013, 1, 697–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogbuewu, I.; Uchegbu, M.; Emenalom, O.; Okoli, I.; Iloeje, M. Overview of the chemistry of soy isoflavones, potential threats and potential therapeutic benefits. Electron. J. Environ. Agric. Food Chem. 2010, 9, 682–695. [Google Scholar]
- Yoon, G.-A.; Park, S. Antioxidant action of soy isoflavones on oxidative stress and antioxidant enzyme activities in exercised rats. Nutr. Res. Pract. 2014, 8, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkor, O.; Shah, N.P. Production of β-Glucosidase and Hydrolysis of Isoflavone Phytoestrogens by Lactobacillus acidophilus, Bifidobacterium lactis, and Lactobacillus casei in Soymilk. J. Food Sci. 2008, 73, M15–M20. [Google Scholar] [CrossRef]
- Hur, H.-G.; Lay, J.O., Jr.; Beger, R.D.; Freeman, J.P.; Rafii, F. Isolation of human intestinal bacteria metabolizing the natural isoflavone glycosides daidzin and genistin. Arch. Microbiol. 2000, 174, 422–428. [Google Scholar] [CrossRef]
- Gaya, P.; Peirotén, Á.; Medina, M.; Landete, J.M. Isoflavone metabolism by a collection of lactic acid bacteria and bifidobacteria with biotechnological interest. Int. J. Food Sci. Nutr. 2016, 67, 117–124. [Google Scholar] [CrossRef]
- Lee, J.H.; Choung, M.-G. Determination of optimal acid hydrolysis time of soybean isoflavones using drying oven and microwave assisted methods. Food Chem. 2011, 129, 577–582. [Google Scholar] [CrossRef]
- Matthäus, B.; Brühl, L. Comparison of different methods for the determination of the oil content in oilseeds. J. Am. Oil Chem. Soc. 2001, 78, 95–102. [Google Scholar] [CrossRef]
- Da Silva, L.H.; Celeghini, R.M.; Chang, Y.K. Effect of the fermentation of whole soybean flour on the conversion of isoflavones from glycosides to aglycones. Food Chem. 2011, 128, 640–644. [Google Scholar] [CrossRef]
- Liu, C.T.; Erh, M.H.; Lin, S.P.; Lo, K.Y.; Chen, K.I.; Cheng, K.C. Enrichment of two isoflavone aglycones in black soymilk by Rhizopus oligosporus NTU 5 in a plastic composite support bioreactor. J. Sci. Food Agric. 2016, 96, 3779–3786. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-I.; Yao, Y.; Chen, H.-J.; Lo, Y.-C.; Yu, R.-C.; Cheng, K.-C. Hydrolysis of isoflavone in black soy milk using cellulose bead as enzyme immobilizer. J. Food. Drug. Anal. 2016, 24, 788–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Castro, C.P.; Drumond, M.M.; Batista, V.L.; Nunes, A.; Mancha-Agresti, P.; Azevedo, V. Vector development timeline for mucosal vaccination and treatment of disease using Lactococcus lactis and design approaches of next generation food grade plasmids. Front. Microbiol. 2018, 9, 1805. [Google Scholar] [CrossRef] [PubMed]
- Michon, C.; Langella, P.; Eijsink, V.; Mathiesen, G.; Chatel, J. Display of recombinant proteins at the surface of lactic acid bacteria: Strategies and applications. Microb. Cell. Fact. 2016, 15, 70. [Google Scholar] [CrossRef]
- Börner, R.A.; Kandasamy, V.; Axelsen, A.M.; Nielsen, A.T.; Bosma, E.F. Genome editing of lactic acid bacteria: Opportunities for food, feed, pharma and biotech. FEMS Microbiol. Lett. 2018, 366, fny291. [Google Scholar] [CrossRef]
- Sybesma, W.; Burgess, C.; Starrenburg, M.; van Sinderen, D.; Hugenholtz, J. Multivitamin production in Lactococcus lactis using metabolic engineering. Metab. Eng. 2004, 6, 109–115. [Google Scholar] [CrossRef]
- Zhang, S.; Shi, Y.; Zhang, S.; Shang, W.; Gao, X.; Wang, H. Whole soybean as probiotic lactic acid bacteria carrier food in solid-state fermentation. Food Control 2014, 41, 1–6. [Google Scholar] [CrossRef]
- Havas, P.; Kun, S.; Perger-Mészáros, I.; Rezessy-Szabó, J.M.; Nguyen, Q.D. Performances of new isolates of Bifidobacterium on fermentation of soymilk. Acta Microbiol. Immunol. Hung. 2015, 62, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [Google Scholar] [CrossRef]
- Li, L.; Shin, S.Y.; Lee, K.; Han, N. Production of natural antimicrobial compound d-phenyllactic acid using Leuconostoc mesenteroides ATCC 8293 whole cells involving highly active d-lactate dehydrogenase. Lett. Appl. Microbiol. 2014, 59, 404–411. [Google Scholar] [CrossRef]
- Sybesma, W.; Starrenburg, M.; Tijsseling, L.; Hoefnagel, M.H.; Hugenholtz, J. Effects of cultivation conditions on folate production by lactic acid bacteria. Appl. Environ. Microbiol. 2003, 69, 4542–4548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.-K.; Ahn, S.-B.; Lee, S.-M.; Shon, M.-Y.; Kim, S.-Y.; Shin, M.-S. Immune-enhancing effects of Leuconostoc strains isolated from Kimchi. J. Biomed. Res. 2012, 13, 353–356. [Google Scholar]
- Eom, H.-J.; Moon, J.-S.; Seo, E.-Y.; Han, N.S. Heterologous expression and secretion of Lactobacillus amylovorus α-amylase in Leuconostoc citreum. Biotechnol. Lett. 2009, 31, 1783–1788. [Google Scholar] [CrossRef] [PubMed]
- Son, J.; Jang, S.H.; Cha, J.W.; Jeong, K.J. Development of CRISPR interference (CRISPRi) platform for metabolic engineering of Leuconostoc citreum and its application for engineering riboflavin biosynthesis. Int. J. Mol. Sci. 2020, 21, 5614. [Google Scholar] [CrossRef]
- Jang, S.H.; Cha, J.W.; Han, N.S.; Jeong, K.J. Development of bicistronic expression system for the enhanced and reliable production of recombinant proteins in Leuconostoc citreum. Sci. Rep. 2018, 8, 8852. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.J.; Ryu, A.J.; Li, L.; Han, N.S.; Jeong, K.J. Development of a high-copy plasmid for enhanced production of recombinant proteins in Leuconostoc citreum. Microb. Cell. Fact. 2016, 15, 12. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Verma, A.; Kumar, V. Catalytic properties, functional attributes and industrial applications of β-glucosidases. 3 Biotech 2016, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Ismail, B.; Hayes, K. β-Glycosidase activity toward different glycosidic forms of isoflavones. J. Agric. Food Chem. 2005, 53, 4918–4924. [Google Scholar] [CrossRef]
- Zubik, L.; Meydani, M. Bioavailability of soybean isoflavones from aglycone and glucoside forms in American women. Am. J. Clin. Nutr. 2003, 77, 1459–1465. [Google Scholar] [CrossRef]
- Peñalvo, J.L.; Nurmi, T.; Adlercreutz, H. A simplified HPLC method for total isoflavones in soy products. Food Chem. 2004, 87, 297–305. [Google Scholar] [CrossRef]
- Youn, S.Y.; Park, M.S.; Ji, G.E. Identification of the β-Glucosidase Gene from Bifidobacterium animalis subsp. lactis and Its Expression in B. bifidum BGN4. J. Microbiol. Biotechnol. 2012, 22, 1714–1723. [Google Scholar]
- Strahsburger, E.; de Lacey, A.M.L.; Marotti, I.; DiGioia, D.; Biavati, B.; Dinelli, G. In vivo assay to identify bacteria with β-glucosidase activity. Electron. J. Biotechnol. 2017, 30, 83–87. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 2013, 127, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Katz, L.; Chen, Y.Y.; Gonzalez, R.; Peterson, T.C.; Zhao, H.; Baltz, R.H. Synthetic biology advances and applications in the biotechnology industry: A perspective. J. Ind. Microbiol. Biotechnol. 2018, 45, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Son, J.; Jeong, K.J. Recent advances in synthetic biology for the engineering of lactic acid bacteria. Biotechnol. Bioprocess Eng. 2020, 25, 962–973. [Google Scholar] [CrossRef]
- Bae, E.-A.; Park, S.-Y.; KIM, D.-H. Constitutive β-glucosidases hydrolyzing ginsenoside Rb1 and Rb2 from human intestinal bacteria. Biol. Pharm. Bull. 2000, 23, 1481–1485. [Google Scholar] [CrossRef]
- Bae, E.-A.; Han, M.J.; Choo, M.-K.; Park, S.-Y.; Kim, D.-H. Metabolism of 20 (S)-and 20 (R)-ginsenoside Rg3 by human intestinal bacteria and its relation to in vitro biological activities. Biol. Pharm. Bull. 2002, 25, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Bae, E.-A.; Han, M.J.; Kim, E.-J.; Kim, D.-H. Transformation of ginseng saponins to ginsenoside Rh2 by acids and human intestinal bacteria and biological activities of their transformants. Arch. Pharm. Res. 2004, 27, 61–67. [Google Scholar] [CrossRef]
- Cheng, D.-L.; Hashimoto, K.; Uda, Y. In vitro digestion of sinigrin and glucotropaeolin by single strains of Bifidobacterium and identification of the digestive products. Food Chem. Toxicol. 2004, 42, 351–357. [Google Scholar] [CrossRef]
- Dabek, M.; McCrae, S.I.; Stevens, V.J.; Duncan, S.H.; Louis, P. Distribution of β-glucosidase and β-glucuronidase activity and of β-glucuronidase gene gus in human colonic bacteria. FEMS Microbiol. Ecol. 2008, 66, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Marotti, I.; Bonetti, A.; Biavati, B.; Catizone, P.; Dinelli, G. Biotransformation of common bean (Phaseolus vulgaris L.) flavonoid glycosides by Bifidobacterium species from human intestinal origin. J. Agric. Food Chem. 2007, 55, 3913–3919. [Google Scholar] [CrossRef] [PubMed]
- Hur, H.-G.; Rafii, F. Biotransformation of the isoflavonoids biochanin A, formononetin, and glycitein by Eubacterium limosum. FEMS Microbiol. Lett. 2000, 192, 21–25. [Google Scholar] [CrossRef]
- Wang, X.-L.; Hur, H.-G.; Lee, J.H.; Kim, K.T.; Kim, S.-I. Enantioselective synthesis of S-equol from dihydrodaidzein by a newly isolated anaerobic human intestinal bacterium. Appl. Environ. Microbiol. 2005, 71, 214–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-L.; Shin, K.-H.; Hur, H.-G.; Kim, S.-I. Enhanced biosynthesis of dihydrodaidzein and dihydrogenistein by a newly isolated bovine rumen anaerobic bacterium. J. Biotechnol. 2005, 115, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Otgonbayar, G.-E.; Eom, H.-J.; Kim, B.-S.; Ko, J.-H.; Han, N.-S. Mannitol production by Leuconostoc citreum KACC 91348P isolated from kimchi. J. Microbiol. Biotechnol. 2011, 21, 968–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Lee, M.; Jung, J.; Kim, J. pIH01, a small cryptic plasmid from Leuconostoc citreum IH3. Plasmid 2005, 54, 184–189. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).