
Mechanical Stretch Induced Skin Regeneration: Molecular and Cellular Mechanism in Skin Soft Tissue Expansion

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Clinical Problem
3. Molecular Mechanism
3.1. Intracellular Signaling under Mechanical Stretch
3.1.1. YAP/TAZ Signaling
3.1.2. MAPK-ERK Signaling
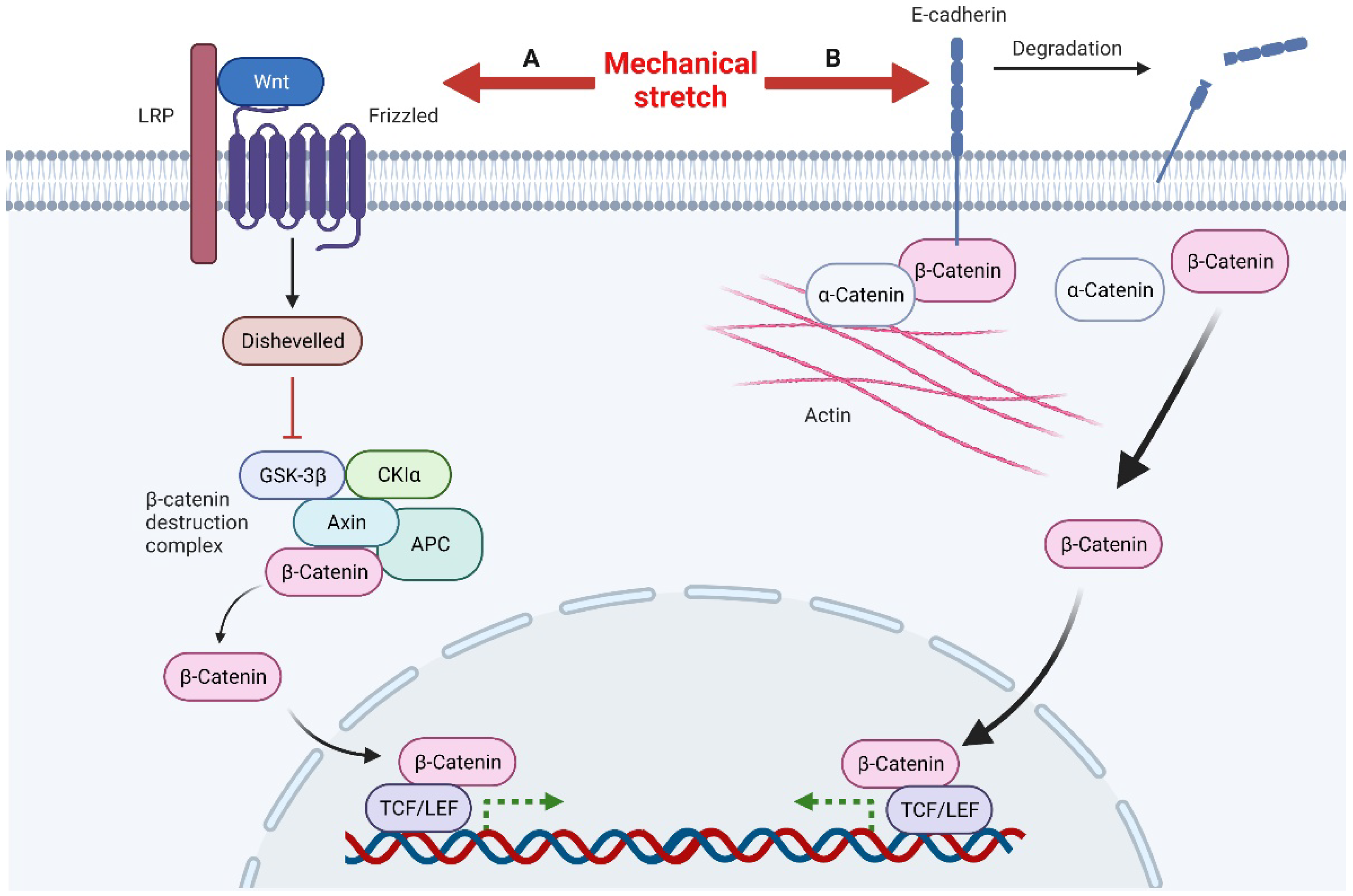
3.1.3. Wnt/β-Catenin Signaling
3.1.4. AP-1
3.2. Cellular Adhesion Molecules That Sense and Transmit Mechanical Signals
3.3. Ion Channels
3.4. Transcriptome Changes Induced by Mechanical Stretch
3.5. Molecular Strategies to Improve Skin Soft Tissue Expansion
4. Cellular Mechanism
4.1. Keratinocyte in Epidermis under Mechanical Stretch
4.2. Fibroblast in Dermis under Mechanical Stretch
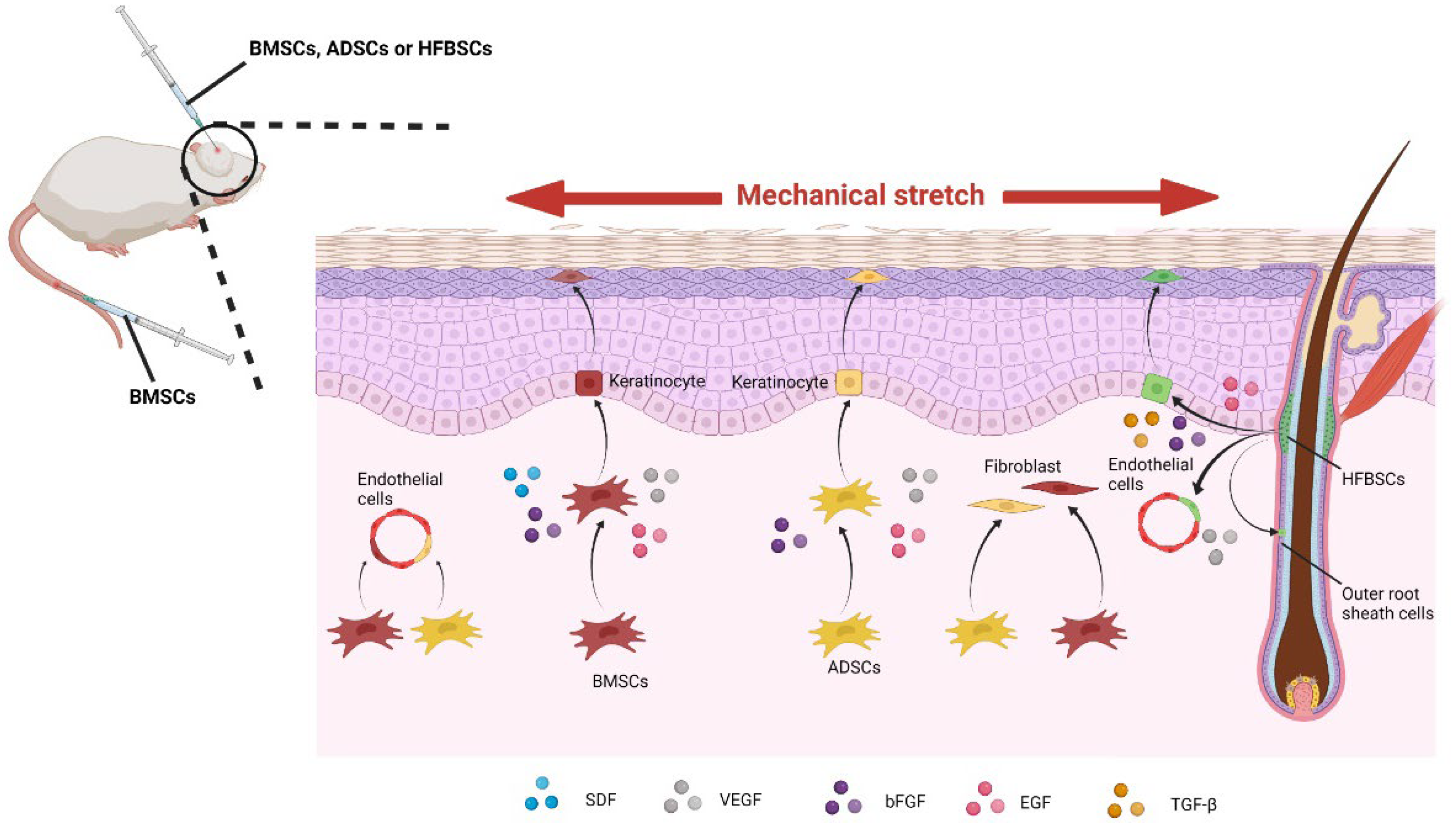
4.3. Mesenchymal Stem Cells under Mechanical Stretch
4.4. Chemical and Mechanical Regulation of Mesenchymal Stem Cells
4.5. Clinical Application of Mesenchymal Stem Cells
4.6. Cellular Strategies to Improve Skin Soft Tissue Expansion
5. Methods to Promote Skin Regeneration under Mechanical Stretch during Skin Soft Tissue Expansion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Radovan, C. Tissue expansion in soft-tissue reconstruction. Plast. Reconstr. Surg. 1984, 74, 482–490. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.L.; Hamilton, K.L.; Monson, L.A.; Buchanan, E.P.; Hollier, L.H., Jr. Tissue Expansion in Children. Semin Plast. Surg. 2016, 30, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhao, J.; Tao, R.; Guo, L.; Yang, H.; Zeng, W.; Song, B.; Xia, W. Repair of Craniomaxillofacial Traumatic Soft Tissue Defects With Tissue Expansion in the Early Stage. J. Craniofac. Surg. 2017, 28, 1477–1480. [Google Scholar] [CrossRef]
- Versaci, A.D.; Balkovich, M.E.; Goldstein, S.A. Breast reconstruction by tissue expansion for congenital and burn deformities. Ann. Plast. Surg. 1986, 16, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.-H.; Kim, S.-Y.; Lee, H.; Lim, H.-K.; Kim, J.-W.; Lee, U.-L.; Lee, J.-B.; Park, S.-H.; Kim, S.-J.; Song, J.-D.; et al. Soft tissue expander for vertically atrophied alveolar ridges: Prospective, multicenter, randomized controlled trial. Clin. Oral Implants Res. 2020, 31, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, Y.G.; Gutiérrez, J.C.L. Multiple Tissue Expansion for Giant Congenital Melanocytic Nevus. Ann. Plast. Surg. 2017, 79, e37–e40. [Google Scholar] [CrossRef]
- Guimarães, C.F.; Gasperini, L.; Marques, A.P.; Reis, R.L. The stiffness of living tissues and its implications for tissue engineering. Nature Reviews Materials 2020, 5, 351–370. [Google Scholar] [CrossRef]
- Yu, Z.; Liu, S.; Cui, J.; Song, Y.; Wang, T.; Song, B.; Peng, P.; Ma, X. Early histological and ultrastructural changes in expanded murine scalp. Ultrastruct. Pathol. 2020, 44, 141–152. [Google Scholar] [CrossRef]
- Liu, S.; Ding, J.; Zhang, Y.; Cheng, X.; Dong, C.; Song, Y.; Yu, Z.; Ma, X. Establishment of a Novel Mouse Model for Soft Tissue Expansion. J. Surg. Res. 2020, 253, 238–244. [Google Scholar] [CrossRef]
- Cherry, G.W.; Austad, E.; Pasyk, K.; McClatchey, K.; Rohrich, R.J. Increased survival and vascularity of random-pattern skin flaps elevated in controlled, expanded skin. Plast. Reconstr. Surg. 1983, 72, 680–685. [Google Scholar] [CrossRef]
- Simon, P.J.; Anderson, L.S.; Manstein, M.E. Increased hair growth and density following controlled expansion of guinea pig skin and soft tissue. Ann. Plast. Surg. 1987, 19, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Misra, J.R.; Irvine, K.D. The Hippo Signaling Network and Its Biological Functions. Annu. Rev. Genet. 2018, 52, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Gao, Y.; Shan, S.; Li, Q. EZH2 Regulates the Correlation between Skin Regeneration and the Duration of Mechanical Stretch. J. Investig. Dermatol. 2021, 141, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Koontz, L.M.; Liu-Chittenden, Y.; Yin, F.; Zheng, Y.; Yu, J.; Huang, B.; Chen, Q.; Wu, S.; Pan, D. The Hippo effector Yorkie controls normal tissue growth by antagonizing scalloped-mediated default repression. Dev. Cell 2013, 25, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Moroishi, T.; Guan, K.L. Mechanisms of Hippo pathway regulation. Genes Dev. 2016, 30, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M.; Panciera, T.; Manfrin, A.; Giulitti, S.; Michielin, F.; Elvassore, N.; Dupont, S.; Piccolo, S. A mechanical checkpoint controls multicellular growth through YAP/TAZ regulation by actin-processing factors. Cell 2013, 154, 1047–1059. [Google Scholar] [CrossRef]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Das, A.; Fischer, R.S.; Pan, D.; Waterman, C.M. YAP Nuclear Localization in the Absence of Cell-Cell Contact Is Mediated by a Filamentous Actin-dependent, Myosin II- and Phospho-YAP-independent Pathway during Extracellular Matrix Mechanosensing. J. Biol. Chem. 2016, 291, 6096–6110. [Google Scholar] [CrossRef]
- Atala, A. Re: Cell Adhesion. Mechanical Strain Induces E-Cadherin-Dependent Yap1 and β-Catenin Activation to Drive Cell Cycle Entry. J. Urol. 2016, 195, 220–221. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Zhang, N.; Wang, C.; Herrler, T.; Li, Q. An updated review of mechanotransduction in skin disorders: Transcriptional regulators, ion channels, and microRNAs. Cell. Mol. Life Sci. 2015, 72, 2091–2106. [Google Scholar] [CrossRef]
- Dasgupta, I.; McCollum, D. Control of cellular responses to mechanical cues through YAP/TAZ regulation. J. Biol. Chem. 2019, 294, 17693–17706. [Google Scholar] [CrossRef]
- Xue, Y.; Lyu, C.; Taylor, A.; Van Ee, A.; Kiemen, A.; Choi, Y.; Khavanian, N.; Henn, D.; Lee, C.; Hwang, L.; et al. Mechanical tension mobilizes Lgr6+ epidermal stem cells to drive skin growth. Sci Adv. 2022, 8, eabl8698. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, H.; Chen, W.; Huang, P.; Bi, J. Human keratinocyte-derived microvesicle miRNA-21 promotes skin wound healing in diabetic rats through facilitating fibroblast function and angiogenesis. Int. J. Biochem. Cell Biol. 2019, 114, 105570. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Zhang, Q.; Wang, W.; Zhang, D.; Jia, J.; Lv, Y.; Yuan, H.; Song, H.; Xiang, F.; et al. Microtubule-associated protein 4 phosphorylation regulates epidermal keratinocyte migration and proliferation. Int. J. Biol. Sci. 2019, 15, 1962–1976. [Google Scholar] [CrossRef]
- Sheikh, A.Q.; Taghian, T.; Hemingway, B.; Cho, H.; Kogan, A.B.; Narmoneva, D.A. Regulation of endothelial MAPK/ERK signalling and capillary morphogenesis by low-amplitude electric field. J. R. Soc. Interface 2013, 10, 20120548. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Blume, P.; Ohgi, S.; Sumpio, B.E. Effect of different frequencies of tensile strain on human dermal fibroblast proliferation and survival. Wound Repair Regen. 2007, 15, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Yang, S.; Cui, Y.-H.; He, Y.-Y. Keratinocyte autophagy enables the activation of keratinocytes and fibroblastsand facilitates wound healing. Autophagy 2021, 17, 2128–2143. [Google Scholar] [CrossRef]
- Jiang, M.; Fang, H.; Shao, S.; Dang, E.; Zhang, J.; Qiao, P.; Yang, A.; Wang, G. Keratinocyte exosomes activate neutrophils and enhance skin inflammation in psoriasis. FASEB J. 2019, 33, 13241–13253. [Google Scholar] [CrossRef]
- Chamcheu, J.C.; Esnault, S.; Adhami, V.M.; Noll, A.L.; Banang-Mbeumi, S.; Roy, T.; Singh, S.S.; Huang, S.; Kousoulas, K.G.; Mukhtar, H. Fisetin, a 3,7,3′,4′-Tetrahydroxyflavone Inhibits the PI3K/Akt/mTOR and MAPK Pathways and Ameliorates Psoriasis Pathology in 2D and 3D Organotypic Human Inflammatory Skin Models. Cells 2019, 8, 1089. [Google Scholar] [CrossRef]
- Kanemaru, K.; Nakamura, Y.; Totoki, K.; Fukuyama, T.; Shoji, M.; Kaneko, H.; Shiratori, K.; Yoneda, A.; Inoue, T.; Iwakura, Y.; et al. Phospholipase Cδ1 regulates p38 MAPK activity and skin barrier integrity. Cell Death Differ. 2017, 24, 1079–1090. [Google Scholar] [CrossRef] [Green Version]
- Claesson-Welsh, L.; Welsh, M. VEGFA and tumour angiogenesis. J. Intern. Med. 2013, 273, 114–127. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, R.; Lôbo, M.; Trindade, K.; Silva, D.F.; Pereira, N. Fibroblast Growth Factors: A Controlling Mechanism of Skin Aging. Skin. Pharmacol. Physiol. 2019, 32, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Wee, P.; Wang, Z. Epidermal Growth Factor Receptor Cell Proliferation Signaling Pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.G.; Miyazu, M.; Xiang, P.; Li, S.N.; Sokabe, M.; Naruse, K. Stretch-induced cell proliferation is mediated by FAK-MAPK pathway. Life Sci. 2005, 76, 2817–2825. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Ro, S.W. MAPK/ERK Signaling Pathway in Hepatocellular Carcinoma. Cancers 2021, 13, 3026. [Google Scholar] [CrossRef]
- Karin, M.; Marshall, C.J. The regulation of AP-1 activity by mitogen-activated protein kinases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 127–134. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- Kiwanuka, E.; Andersson, L.; Caterson, E.J.; Junker, J.P.; Gerdin, B.; Eriksson, E. CCN2 promotes keratinocyte adhesion and migration via integrin α5β1. Exp. Cell Res. 2013, 319, 2938–2946. [Google Scholar] [CrossRef]
- Stern, C.D. Neural induction: Old problem, new findings, yet more questions. Development 2005, 132, 2007–2021. [Google Scholar] [CrossRef]
- Zhu, X.J.; Liu, Y.; Dai, Z.M.; Zhang, X.; Yang, X.; Li, Y.; Qiu, M.; Fu, J.; Hsu, W.; Chen, Y.; et al. BMP-FGF signaling axis mediates Wnt-induced epidermal stratification in developing mammalian skin. PLoS Genet. 2014, 10, e1004687. [Google Scholar] [CrossRef] [Green Version]
- Houschyar, K.S.; Borrelli, M.R.; Tapking, C.; Popp, D.; Puladi, B.; Ooms, M.; Chelliah, M.P.; Rein, S.; Pförringer, D.; Thor, D.; et al. Molecular Mechanisms of Hair Growth and Regeneration: Current Understanding and Novel Paradigms. Dermatology 2020, 236, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Zhang, Y.; Xu, M.; Yang, Y.; Ito, M.; Peng, T.; Cui, Z.; Nagy, A.; Hadjantonakis, A.-K.; Lang, R.A.; et al. Distinct functions for Wnt/β-catenin in hair follicle stem cell proliferation and survival and interfollicular epidermal homeostasis. Cell Stem Cell 2013, 13, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Lim, X.; Tan, S.H.; Koh, W.L.C.; Chau, R.M.W.; Yan, K.S.; Kuo, C.J.; van Amerongen, R.; Klein, A.M.; Nusse, R. Interfollicular epidermal stem cells self-renew via autocrine Wnt signaling. Science 2013, 342, 1226–1230. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, W.; Zhao, Y.; Gao, L.; Chang, Y.; Tong, Z.; Li, H.; Jing, J. Cyclic Mechanical Strain Regulates Osteoblastic Differentiation of Mesenchymal Stem Cells on TiO2 Nanotubes Through GCN5 and Wnt/β-Catenin. Front. Bioeng. Biotechnol. 2021, 9, 735949. [Google Scholar] [CrossRef]
- You, Y.; Zheng, Q.; Dong, Y.; Wang, Y.; Zhang, L.; Xue, T.; Xie, X.; Hu, C.; Wang, Z.; Chen, R.; et al. Higher Matrix Stiffness Upregulates Osteopontin Expression in Hepatocellular Carcinoma Cells Mediated by Integrin β1/GSK3β/β-Catenin Signaling Pathway. PLoS ONE 2015, 10, e0134243. [Google Scholar] [CrossRef]
- Samuel, M.S.; Lopez, J.I.; McGhee, E.J.; Croft, D.R.; Strachan, D.; Timpson, P.; Munro, J.; Schröder, E.; Zhou, J.; Brunton, V.G.; et al. Actomyosin-mediated cellular tension drives increased tissue stiffness and β-catenin activation to induce epidermal hyperplasia and tumor growth. Cancer Cell 2011, 19, 776–791. [Google Scholar] [CrossRef]
- Cheng, X.; Yu, Z.; Song, Y.; Zhang, Y.; Du, J.; Su, Y.; Ma, X. Hair follicle bulge-derived stem cells promote tissue regeneration during skin expansion. Biomed. Pharmacother. 2020, 132, 110805. [Google Scholar] [CrossRef]
- Liu, W.; Xiong, S.; Zhang, Y.; Du, J.; Dong, C.; Yu, Z.; Ma, X. Transcriptome Profiling Reveals Important Transcription Factors and Biological Processes in Skin Regeneration Mediated by Mechanical Stretch. Front. Genet. 2021, 12, 757350. [Google Scholar] [CrossRef]
- Chu, S.-Y.; Chou, C.-H.; Huang, H.-D.; Yen, M.-H.; Hong, H.-C.; Chao, P.-H.; Wang, Y.-H.; Chen, P.-Y.; Nian, S.-X.; Chen, Y.-R.; et al. Mechanical stretch induces hair regeneration through the alternative activation of macrophages. Nat. Commun. 2019, 10, 1524. [Google Scholar] [CrossRef]
- Ledwon, J.K.; Vaca, E.E.; Huang, C.C.; Kelsey, L.J.; McGrath, J.L.; Topczewski, J.; Gosain, A.K.; Topczewska, J.M. Langerhans cells and SFRP2/Wnt/beta-catenin signalling control adaptation of skin epidermis to mechanical stretching. J. Cell. Mol. Med. 2022, 26, 764–775. [Google Scholar] [CrossRef]
- Heuberger, J.; Birchmeier, W. Interplay of cadherin-mediated cell adhesion and canonical Wnt signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a002915. [Google Scholar] [CrossRef]
- Eastman, Q.; Grosschedl, R. Regulation of LEF-1/TCF transcription factors by Wnt and other signals. Curr. Opin. Cell Biol. 1999, 11, 233–240. [Google Scholar] [CrossRef]
- Huang, X.; Liang, X.; Zhou, Y.; Li, H.; Du, H.; Suo, Y.; Liu, W.; Jin, R.; Chai, B.; Duan, R.; et al. CDH1 is Identified as A Therapeutic Target for Skin Regeneration after Mechanical Loading. Int. J. Biol. Sci. 2021, 17, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Zenz, R.; Eferl, R.; Scheinecker, C.; Redlich, K.; Smolen, J.; Schonthaler, H.B.; Kenner, L.; Tschachler, E.; Wagner, E.F. Activator protein 1 (Fos/Jun) functions in inflammatory bone and skin disease. Arthritis Res. Ther. 2008, 10, 201. [Google Scholar] [CrossRef]
- Angel, P.; Szabowski, A.; Schorpp-Kistner, M. Function and regulation of AP-1 subunits in skin physiology and pathology. Oncogene 2001, 20, 2413–2423. [Google Scholar] [CrossRef] [PubMed]
- Yates, S.; Rayner, T.E. Transcription factor activation in response to cutaneous injury: Role of AP-1 in reepithelialization. Wound Repair Regen. 2002, 10, 5–15. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Iliadi, A.; Eliades, T.; Kletsas, D. Early responses of human periodontal ligament fibroblasts to cyclic and static mechanical stretching. Eur. J. Orthod. 2017, 39, 258–263. [Google Scholar] [CrossRef]
- Aragona, M.; Sifrim, A.; Malfait, M.; Song, Y.; Van Herck, J.; Dekoninck, S.; Gargouri, S.; Lapouge, G.; Swedlund, B.; Dubois, C.; et al. Mechanisms of stretch-mediated skin expansion at single-cell resolution. Nature 2020, 584, 268–273. [Google Scholar] [CrossRef]
- Zanconato, F.; Forcato, M.; Battilana, G.; Azzolin, L.; Quaranta, E.; Bodega, B.; Rosato, A.; Bicciato, S.; Cordenonsi, M.; Piccolo, S. Genome-wide association between YAP/TAZ/TEAD and AP-1 at enhancers drives oncogenic growth. Nat. Cell Biol. 2015, 17, 1218–1227. [Google Scholar] [CrossRef]
- Sun, Z.; Costell, M.; Fässler, R. Integrin activation by talin, kindlin and mechanical forces. Nat. Cell Biol. 2019, 21, 25–31. [Google Scholar] [CrossRef]
- Chen, T.; Saw, T.B.; Mège, R.M.; Ladoux, B. Mechanical forces in cell monolayers. J. Cell Sci. 2018, 131, jcs218156. [Google Scholar] [CrossRef] [PubMed]
- Kanchanawong, P.; Shtengel, G.; Pasapera, A.M.; Ramko, E.B.; Davidson, M.W.; Hess, H.F.; Waterman, C.M. Nanoscale architecture of integrin-based cell adhesions. Nature 2010, 468, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Troyanovsky, R.B.; Troyanovsky, S.M. Binding to F-actin guides cadherin cluster assembly, stability, and movement. J. Cell Biol. 2013, 201, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Kenny, F.N.; Connelly, J.T. Integrin-mediated adhesion and mechano-sensing in cutaneous wound healing. Cell Tissue Res. 2015, 360, 571–582. [Google Scholar] [CrossRef]
- Petridou, N.I.; Skourides, P.A. A ligand-independent integrin β1 mechanosensory complex guides spindle orientation. Nat. Commun. 2016, 7, 10899. [Google Scholar] [CrossRef]
- Ferraris, G.M.S.; Schulte, C.; Buttiglione, V.; De Lorenzi, V.; Piontini, A.; Galluzzi, M.; Podestà, A.; Madsen, C.D.; Sidenius, N. The interaction between uPAR and vitronectin triggers ligand-independent adhesion signalling by integrins. EMBO J. 2014, 33, 2458–2472. [Google Scholar] [CrossRef]
- Mitra, S.K.; Hanson, D.A.; Schlaepfer, D.D. Focal adhesion kinase: In command and control of cell motility. Nat. Rev. Mol. Cell Biol. 2005, 6, 56–68. [Google Scholar] [CrossRef]
- Elbediwy, A.; Thompson, B.J. Evolution of mechanotransduction via YAP/TAZ in animal epithelia. Curr. Opin. Cell Biol. 2018, 51, 117–123. [Google Scholar] [CrossRef]
- Topczewska, J.M.; Ledwon, J.K.; Vaca, E.E.; Gosain, A.K. Mechanical stretching stimulates growth of the basal layer and rete ridges in the epidermis. J. Tissue Eng. Regen. Med. 2019, 13, 2121–2125. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, H.; Liang, X.; Du, H.; Suo, Y.; Chen, H.; Liu, W.; Duan, R.; Huang, X.; Li, Q. The CCN1 (CYR61) protein promotes skin growth by enhancing epithelial-mesenchymal transition during skin expansion. J. Cell. Mol. Med. 2020, 24, 1460–1473. [Google Scholar] [CrossRef]
- Lau, L.F. CCN1/CYR61: The very model of a modern matricellular protein. Cell. Mol. Life Sci. 2011, 68, 3149–3163. [Google Scholar] [CrossRef] [PubMed]
- Hodorogea, A.; Calinescu, A.; Antohe, M.; Balaban, M.; Nedelcu, R.I.; Turcu, G.; Ion, D.A.; Badarau, I.A.; Popescu, C.M.; Popescu, R.; et al. Epithelial-Mesenchymal Transition in Skin Cancers: A Review. Anal. Cell. Pathol. 2019, 2019, 3851576. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.S.; Chouhan, G.; Belapurkar, V.; Arora, P.; Satyanarayan; Ainavarapu, S.R.K.; Sonawane, M. A new tension induction paradigm unravels tissue response and the importance of E-cadherin in the developing epidermis. Int. J. Dev. Biol. 2020, 64, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Uttagomol, J.; Ahmad, U.S.; Rehman, A.; Huang, Y.; Laly, A.C.; Kang, A.; Soetaert, J.; Chance, R.; Teh, M.-T.; Connelly, J.T.; et al. Evidence for the Desmosomal Cadherin Desmoglein-3 in Regulating YAP and Phospho-YAP in Keratinocyte Responses to Mechanical Forces. Int. J. Mol. Sci. 2019, 20, 6221. [Google Scholar] [CrossRef]
- Kefauver, J.M.; Ward, A.B.; Patapoutian, A. Discoveries in structure and physiology of mechanically activated ion channels. Nature 2020, 587, 567–576. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Fang, B.; Shan, S.; Xie, Y.; Wang, C.; Zhang, Y.; Zhang, X.; Li, Q. Mechanical stretch promotes hypertrophic scar formation through mechanically activated cation channel Piezo1. Cell Death Dis. 2021, 12, 226. [Google Scholar] [CrossRef]
- Baylie, R.L.; Brayden, J.E. TRPV channels and vascular function. Acta Physiol. 2011, 203, 99–116. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Xue, C.; Chen, H.; Xue, Y.; Zhao, F.; Zhu, M.X.; Cao, Z. TRPV3 enhances skin keratinocyte proliferation through EGFR-dependent signaling pathways. Cell Biol. Toxicol. 2021, 37, 313–330. [Google Scholar] [CrossRef]
- Szöllősi, A.G.; Vasas, N.; Angyal, Á.; Kistamás, K.; Nánási, P.P.; Mihály, J.; Béke, G.; Herczeg-Lisztes, E.; Szegedi, A.; Kawada, N.; et al. Activation of TRPV3 Regulates Inflammatory Actions of Human Epidermal Keratinocytes. J. Investig. Dermatol. 2018, 138, 365–374. [Google Scholar] [CrossRef]
- Wei, J.J.; Kim, H.S.; Spencer, C.A.; Brennan-Crispi, D.; Zheng, Y.; Johnson, N.M.; Rosenbach, M.; Miller, C.; Leung, D.H.; Cotsarelis, G.; et al. Activation of TRPA1 nociceptor promotes systemic adult mammalian skin regeneration. Sci. Immunol. 2020, 5, eaba5683. [Google Scholar] [CrossRef]
- Ledwon, J.K.; Kelsey, L.J.; Vaca, E.E.; Gosain, A.K. Transcriptomic analysis reveals dynamic molecular changes in skin induced by mechanical forces secondary to tissue expansion. Sci. Rep. 2020, 10, 15991. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Liu, W.; Zhang, Y.; Song, Y.; Du, J.; Huang, Z.; Wang, T.; Yu, Z.; Ma, X. Identification of Common Hub Genes in Human Dermal Fibroblasts Stimulated by Mechanical Stretch at Both the Early and Late Stages. Front. Surg. 2022, 9, 846161. [Google Scholar] [CrossRef]
- Choi, J.E.; Di Nardo, A. Skin neurogenic inflammation. Semin. Immunopathol. 2018, 40, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Cowin, A.J.; Bayat, A.; Murray, R.Z.; Kopecki, Z. Editorial: Inflammation in Healing and Regeneration of Cutaneous Wounds. Front. Immunol. 2021, 12, 806687. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Lei, L.; Liu, S.; Zhang, Y.; Yu, Z.; Su, Y.; Ma, X. Macrophages are necessary for skin regeneration during tissue expansion. J. Transl. Med. 2019, 17, 36. [Google Scholar] [CrossRef]
- Kimura, S.; Tsuji, T. Mechanical and Immunological Regulation in Wound Healing and Skin Reconstruction. Int. J. Mol. Sci. 2021, 22, 5474. [Google Scholar] [CrossRef]
- Furue, M.; Furue, K.; Tsuji, G.; Nakahara, T. Interleukin-17A and Keratinocytes in Psoriasis. Int. J. Mol. Sci. 2020, 21, 1275. [Google Scholar] [CrossRef]
- Rath-Deschner, B.; Memmert, S.; Damanaki, A.; de Molon, R.S.; Nokhbehsaim, M.; Eick, S.; Kirschneck, C.; Cirelli, J.A.; Deschner, J.; Jäger, A.; et al. CXCL5, CXCL8, and CXCL10 regulation by bacteria and mechanical forces in periodontium. Ann. Anat.-Anat. Anz. 2021, 234, 151648. [Google Scholar] [CrossRef]
- Nazet, U.; Feulner, L.; Muschter, D.; Neubert, P.; Schatz, V.; Grässel, S.; Jantsch, J.; Proff, P.; Schröder, A.; Kirschneck, C. Mechanical Stress Induce PG-E2 in Murine Synovial Fibroblasts Originating from the Temporomandibular Joint. Cells 2021, 10, 298. [Google Scholar] [CrossRef]
- Nakamura, Y.; Matsuzaka, T.; Tahara-Hanaoka, S.; Shibuya, K.; Shimano, H.; Nakahashi-Oda, C.; Shibuya, A. Elovl6 regulates mechanical damage-induced keratinocyte death and skin inflammation. Cell Death Dis. 2018, 9, 1181. [Google Scholar] [CrossRef] [Green Version]
- Devalaraja, R.M.; Nanney, L.B.; Qian, Q.; Du, J.; Yu, Y.; Devalaraja, M.N.; Richmond, A. Delayed wound healing in CXCR2 knockout mice. J. Investig. Dermatol. 2000, 115, 234–244. [Google Scholar] [CrossRef]
- Korbecki, J.; Kupnicka, P.; Chlubek, M.; Gorący, J.; Gutowska, I.; Baranowska-Bosiacka, I. CXCR2 Receptor: Regulation of Expression, Signal Transduction, and Involvement in Cancer. Int. J. Mol. Sci. 2022, 23, 2168. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, P.J.; Gallagher, R.; Seaton, A.; Wilson, C.; Scullin, P.; Pettigrew, J.; Stratford, I.J.; Williams, K.J.; Johnston, P.G.; Waugh, D.J.J. HIF-1 and NF-κB-mediated upregulation of CXCR1 and CXCR2 expression promotes cell survival in hypoxic prostate cancer cells. Oncogene 2007, 26, 7333–7345. [Google Scholar] [CrossRef] [PubMed]
- Meitei, H.T.; Jadhav, N.; Lal, G. CCR6-CCL20 axis as a therapeutic target for autoimmune diseases. Autoimmun. Rev. 2021, 20, 102846. [Google Scholar] [CrossRef] [PubMed]
- Giang, J.; Seelen, M.A.J.; van Doorn, M.B.A.; Rissmann, R.; Prens, E.P.; Damman, J. Complement Activation in Inflammatory Skin Diseases. Front. Immunol. 2018, 9, 639. [Google Scholar] [CrossRef]
- Warwick, C.A.; Shutov, L.P.; Shepherd, A.J.; Mohapatra, D.P.; Usachev, Y.M. Mechanisms underlying mechanical sensitization induced by complement C5a: The roles of macrophages, TRPV1, and calcitonin gene-related peptide receptors. Pain 2019, 160, 702–711. [Google Scholar] [CrossRef]
- Manicone, A.M.; McGuire, J.K. Matrix metalloproteinases as modulators of inflammation. Semin. Cell Dev. Biol. 2008, 19, 34–41. [Google Scholar] [CrossRef]
- Rohani, M.G.; Parks, W.C. Matrix remodeling by MMPs during wound repair. Matrix Biol. 2015, 44–46, 113–121. [Google Scholar] [CrossRef]
- Li, W.-D.; Zhou, D.-M.; Sun, L.-L.; Xiao, L.; Liu, Z.; Zhou, M.; Wang, W.-B.; Li, X.-Q. LncRNA WTAPP1 Promotes Migration and Angiogenesis of Endothelial Progenitor Cells via MMP1 Through MicroRNA 3120 and Akt/PI3K/Autophagy Pathways. Stem Cells 2018, 36, 1863–1874. [Google Scholar] [CrossRef]
- Sun, L.-L.; Lei, F.-R.; Jiang, X.-D.; Du, X.-L.; Xiao, L.; Li, W.-D.; Li, X.-Q. LncRNA GUSBP5-AS promotes EPC migration and angiogenesis and deep vein thrombosis resolution by regulating FGF2 and MMP2/9 through the miR-223-3p/FOXO1/Akt pathway. Aging 2020, 12, 4506–4526. [Google Scholar] [CrossRef]
- Yang, L.-Q.; Yu, S.-P.; Yang, Y.-T.; Zhao, Y.-S.; Wang, F.-Y.; Chen, Y.; Li, Q.-H.; Tian, P.; Zhu, Y.-Y.; Zhang, J.-G.; et al. Muscone derivative ZM-32 inhibits breast tumor angiogenesis by suppressing HuR-mediated VEGF and MMP9 expression. Biomed. Pharmacother. 2021, 136, 111265. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Lin, Y.; Liao, B.; Zhou, L.; Ai, J.; Jin, X.; Li, H.; Wang, K. The role of interleukin-6/interleukin-6 receptor signaling in the mechanical stress-induced extracellular matrix remodeling of bladder smooth muscle. Arch. Biochem. Biophys. 2021, 702, 108674. [Google Scholar] [CrossRef] [PubMed]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [PubMed]
- Behm, C.; Nemec, M.; Weissinger, F.; Rausch, M.A.; Andrukhov, O.; Jonke, E. MMPs and TIMPs Expression Levels in the Periodontal Ligament during Orthodontic Tooth Movement: A Systematic Review of In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2021, 22, 6967. [Google Scholar] [CrossRef]
- Hohmann, T.; Dehghani, F. The Cytoskeleton—A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef]
- Molnar, K.; Labouesse, M. The plastic cell: Mechanical deformation of cells and tissues. Open Biol. 2021, 11, 210006. [Google Scholar] [CrossRef]
- Liu, W.; Xiong, S.; Du, J.; Song, Y.; Wang, T.; Zhang, Y.; Dong, C.; Huang, Z.; He, Q.; Yu, Z.; et al. Deciphering Key Foreign Body Reaction-Related Transcription Factors and Genes Through Transcriptome Analysis. Front. Mol. Biosci. 2022, 9, 843391. [Google Scholar] [CrossRef]
- Eger, A.; Stockinger, A.; Schaffhauser, B.; Beug, H.; Foisner, R. Epithelial mesenchymal transition by c-Fos estrogen receptor activation involves nuclear translocation of β-catenin and upregulation of β-catenin/lymphoid enhancer binding factor-1 transcriptional activity. J. Cell Biol. 2000, 148, 173–187. [Google Scholar] [CrossRef]
- Kim, S.; Song, G.; Lee, T.; Kim, M.; Kim, J.; Kwon, H.; Kim, J.; Jeong, W.; Lee, U.; Na, C.; et al. PARsylated transcription factor EB (TFEB) regulates the expression of a subset of Wnt target genes by forming a complex with β-catenin-TCF/LEF1. Cell Death Differ. 2021, 28, 2555–2570. [Google Scholar] [CrossRef]
- Phan, Q.M.; Fine, G.M.; Salz, L.; Herrera, G.G.; Wildman, B.; Driskell, I.M.; Driskell, R.R. Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds. eLife 2020, 9, e60066. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, Y.; Sun, C.; Wang, T.; Shen, Y.; Cai, W.; Sun, J.; Chi, L.; Wang, H.; Song, N.; et al. Feedback Activation of Basic Fibroblast Growth Factor Signaling via the Wnt/β-Catenin Pathway in Skin Fibroblasts. Front. Pharmacol. 2017, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Lu, C.; Wang, W.; Gao, W.; Yu, C. circBANP promotes colorectal cancer growth and metastasis via sponging let-7d-5p to modulate HMGA1/Wnt/β-catenin signaling. Mol. Ther. Oncolytics 2021, 21, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, W.; Fan, J.; Liu, L.; Tian, J.; Gan, C.; Jiao, H.; Yang, Z. Effect of Mechanical Tension on the circRNA Expression Profile of Human Skin Tissue. J. Craniofac. Surg. 2019, 30, e474–e477. [Google Scholar] [CrossRef]
- Qiao, P.; Guo, W.; Ke, Y.; Fang, H.; Zhuang, Y.; Jiang, M.; Zhang, J.; Shen, S.; Qiao, H.; Dang, E.; et al. Mechanical Stretch Exacerbates Psoriasis by Stimulating Keratinocyte Proliferation and Cytokine Production. J. Investig. Dermatol. 2019, 139, 1470–1479. [Google Scholar] [CrossRef] [PubMed]
- Takei, T.; Rivas-Gotz, C.; Delling, C.A.; Koo, J.T.; Mills, I.; McCarthy, T.L.; Centrella, M.; Sumpio, B.E. Effect of strain on human keratinocytes in vitro. J. Cell. Physiol. 1997, 173, 64–72. [Google Scholar] [CrossRef]
- Kippenberger, S.; Loitsch, S.; Guschel, M.; Müller, J.; Knies, Y.; Kaufmann, R.; Bernd, A. Mechanical stretch stimulates protein kinase B/Akt phosphorylation in epidermal cells via angiotensin II type 1 receptor and epidermal growth factor receptor. J. Biol. Chem. 2005, 280, 3060–3067. [Google Scholar] [CrossRef]
- Pincelli, C.; Marconi, A. Keratinocyte stem cells: Friends and foes. J. Cell. Physiol. 2010, 225, 310–315. [Google Scholar] [CrossRef]
- Houben, E.; De Paepe, K.; Rogiers, V. A keratinocyte’s course of life. Skin. Pharmacol. Physiol. 2007, 20, 141–147. [Google Scholar] [CrossRef]
- Kretzschmar, K.; Boonekamp, K.E.; Bleijs, M.; Asra, P.; Koomen, M.; de Sousa Lopes, S.M.C.; Giovannone, B.; Clevers, H. Troy/Tnfrsf19 marks epidermal cells that govern interfollicular epidermal renewal and cornification. Stem Cell Rep. 2021, 16, 2379–2394. [Google Scholar] [CrossRef]
- Rippa, A.L.; Kalabusheva, E.P.; Vorotelyak, E.A. Regeneration of Dermis: Scarring and Cells Involved. Cells 2019, 8, 607. [Google Scholar] [CrossRef] [Green Version]
- Yannas, I.V.; Tzeranis, D.S. Mammals fail to regenerate organs when wound contraction drives scar formation. NPJ Regen. Med. 2021, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Aarabi, S.; Bhatt, K.A.; Shi, Y.; Paterno, J.; Chang, E.I.; Loh, S.A.; Holmes, J.W.; Longaker, M.T.; Yee, H.; Gurtner, G.C. Mechanical load initiates hypertrophic scar formation through decreased cellular apoptosis. FASEB J. 2007, 21, 3250–3261. [Google Scholar] [CrossRef] [PubMed]
- Webb, K.; Hitchcock, R.W.; Smeal, R.M.; Li, W.; Gray, S.D.; Tresco, P.A. Cyclic strain increases fibroblast proliferation, matrix accumulation, and elastic modulus of fibroblast-seeded polyurethane constructs. J. Biomech. 2006, 39, 1136–1144. [Google Scholar] [CrossRef]
- Rolin, G.L.; Binda, D.; Tissot, M.; Viennet, C.; Saas, P.; Muret, P.; Humbert, P. In vitro study of the impact of mechanical tension on the dermal fibroblast phenotype in the context of skin wound healing. J. Biomech. 2014, 47, 3555–3561. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Yang, W.; Shangbin, L.; Tingting, L.; Yang, Z.; Feng, G.; Qingping, Y.; Wenhao, Z. A microscopic and biomechanical study of skin and soft tissue after repeated expansion. Dermatol. Surg. 2009, 35, 72–79. [Google Scholar] [CrossRef]
- Park, S.; Jung, S.-C. New Sources, Differentiation, and Therapeutic Uses of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 5288. [Google Scholar] [CrossRef]
- Mazini, L.; Rochette, L.; Admou, B.; Amal, S.; Malka, G. Hopes and Limits of Adipose-Derived Stem Cells (ADSCs) and Mesenchymal Stem Cells (MSCs) in Wound Healing. Int. J. Mol. Sci. 2020, 21, 1306. [Google Scholar] [CrossRef]
- Li, C.; Zheng, Y.; Wang, X.; Xia, W.; Gao, H.; Li, D.; Ma, X. Bone marrow-derived stem cells contribute skin regeneration in skin and soft tissue expansion. J. Cell. Physiol. 2011, 226, 2834–2840. [Google Scholar] [CrossRef]
- Yang, M.; Li, Q.; Sheng, L.; Li, H.; Weng, R.; Zan, T. Bone marrow–derived mesenchymal stem cells transplantation accelerates tissue expansion by promoting skin regeneration during expansion. Ann. Surg. 2011, 253, 202–209. [Google Scholar] [CrossRef]
- Liang, X.; Huang, X.; Zhou, Y.; Jin, R.; Li, Q. Mechanical Stretching Promotes Skin Tissue Regeneration via Enhancing Mesenchymal Stem Cell Homing and Transdifferentiation. Stem Cells Transl. Med. 2016, 5, 960–969. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, C.; Zheng, Y.; Xia, W.; Yu, Y.; Ma, X. Bone marrow mesenchymal stem cells increase skin regeneration efficiency in skin and soft tissue expansion. Expert Opin. Biol. Ther. 2012, 12, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.B.; Wang, J.; Chiang, C.A.; Sheng, L.L.; Li, Q.F. Mechanical stretch upregulates SDF-1α in skin tissue and induces migration of circulating bone marrow-derived stem cells into the expanded skin. Stem Cells 2013, 31, 2703–2713. [Google Scholar] [CrossRef] [PubMed]
- Bora, P.; Majumdar, A.S. Adipose tissue-derived stromal vascular fraction in regenerative medicine: A brief review on biology and translation. Stem Cell Res. Ther. 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Yang, M.; Du, Z.; Yang, Y.; Li, Q. Transplantation of stromal vascular fraction as an alternative for accelerating tissue expansion. J. Plast. Reconstr. Aesthet. Surg. 2013, 66, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.-C.; Chao, P.-C.; Cheng, C.; Chen, C.-H.; Huang, R.-L.; Zhou, S.-B.; Xie, Y.; Li, Q.-F. A randomized, controlled clinical trial of autologous stromal vascular fraction cells transplantation to promote mechanical stretch-induced skin regeneration. Stem Cell Res. Ther. 2021, 12, 243. [Google Scholar] [CrossRef]
- An, R.; Li, S.; Sheng, L.; Cao, W. Emulsified Fat Grafting Accelerates Tissue Expansion: An Experimental Study in a Rat Model. Ann. Plast Surg 2020, 85, 61–67. [Google Scholar] [CrossRef]
- Riekstina, U.; Muceniece, R.; Cakstina, I.; Muiznieks, I.; Ancans, J. Characterization of human skin-derived mesenchymal stem cell proliferation rate in different growth conditions. Cytotechnology 2008, 58, 153–162. [Google Scholar] [CrossRef]
- Amoh, Y.; Hoffman, R.M. Hair follicle-associated-pluripotent (HAP) stem cells. Cell Cycle 2017, 16, 2169–2175. [Google Scholar] [CrossRef]
- 1Ito, M.; Liu, Y.; Yang, Z.; Nguyen, J.; Liang, F.; Morris, R.J.; Cotsarelis, G. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat. Med. 2005, 11, 1351–1354. [Google Scholar] [CrossRef]
- Li, B.; Hu, W.; Ma, K.; Zhang, C.; Fu, X. Are hair follicle stem cells promising candidates for wound healing? Expert Opin. Biol. Ther. 2019, 19, 119–128. [Google Scholar] [CrossRef]
- Qiu, W.; Lei, M.; Li, J.; Wang, N.; Lian, X. Activated hair follicle stem cells and Wnt/β-catenin signaling involve in pathnogenesis of sebaceous neoplasms. Int. J. Med. Sci. 2014, 11, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Bastakoty, D.; Young, P.P. Wnt/β-catenin pathway in tissue injury: Roles in pathology and therapeutic opportunities for regeneration. FASEB J. 2016, 30, 3271–3284. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Feng, G.; Hu, X.; Yang, L.; Yang, X.; Jin, Q. SDF Factor-1α Promotes the Migration, Proliferation, and Osteogenic Differentiation of Mouse Bone Marrow Mesenchymal Stem Cells Through the Wnt/β-Catenin Pathway. Stem Cells Dev. 2021, 30, 106–117. [Google Scholar] [CrossRef]
- Pruijt, J.F.M.; Fibbe, W.E.; Laterveer, L.; Pieters, R.A.; Lindley, I.J.D.; Paemen, L.; Masure, S.; Willemze, R.; Opdenakker, G. Prevention of interleukin-8-induced mobilization of hematopoietic progenitor cells in rhesus monkeys by inhibitory antibodies against the metalloproteinase gelatinase B (MMP-9). Proc. Natl. Acad. Sci. USA 1999, 96, 10863–10868. [Google Scholar] [CrossRef]
- Schmidt, A.; Ladage, D.; Schinköthe, T.; Klausmann, U.; Ulrichs, C.; Klinz, F.-J.; Brixius, K.; Arnhold, S.; Desai, B.; Mehlhorn, U.; et al. Basic fibroblast growth factor controls migration in human mesenchymal stem cells. Stem Cells 2006, 24, 1750–1758. [Google Scholar] [CrossRef]
- Ball, S.G.; Shuttleworth, C.A.; Kielty, C.M. Vascular endothelial growth factor can signal through platelet-derived growth factor receptors. J. Cell Biol. 2007, 177, 489–500. [Google Scholar] [CrossRef]
- Langer, H.F.; Stellos, K.; Steingen, C.; Froihofer, A.; Schönberger, T.; Krämer, B.; Bigalke, B.; May, A.E.; Seizer, P.; Müller, I.; et al. Platelet derived bFGF mediates vascular integrative mechanisms of mesenchymal stem cells in vitro. J. Mol. Cell. Cardiol. 2009, 47, 315–325. [Google Scholar] [CrossRef]
- Li, Q.; Xia, S.; Fang, H.; Pan, J.; Jia, Y.; Deng, G. VEGF treatment promotes bone marrow-derived CXCR4+ mesenchymal stromal stem cell differentiation into vessel endothelial cells. Exp. Ther. Med. 2017, 13, 449–454. [Google Scholar] [CrossRef]
- Zhu, H.; Lan, L.; Zhang, Y.; Chen, Q.; Zeng, Y.; Luo, X.; Ren, J.; Chen, S.; Xiao, M.; Lin, K.; et al. Epidermal growth factor stimulates exosomal microRNA-21 derived from mesenchymal stem cells to ameliorate aGVHD by modulating regulatory T cells. FASEB J. 2020, 34, 7372–7386. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.; Liu, F.; Zou, F.; Zhao, G.; Jiang, Y.; Liu, L.; Shi, J.; Hao, D.; Zhang, Q.; Zheng, T.; et al. Epidermal Growth Factor Induces Proliferation of Hair Follicle-Derived Mesenchymal Stem Cells Through Epidermal Growth Factor Receptor-Mediated Activation of ERK and AKT Signaling Pathways Associated with Upregulation of Cyclin D1 and Downregulation of p16. Stem Cells Dev. 2017, 26, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Liu, Y.; Zheng, D.; Shan, S.; Wang, C.; Gao, Y.; Wang, J.; Xie, Y.; Zhang, Y.; Li, Q. The effects of mechanical stretch on the biological characteristics of human adipose-derived stem cells. J. Cell. Mol. Med. 2019, 23, 4244–4255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Luo, Q.; Chen, Z.; Sun, J.; Xu, B.; Ju, Y.; Song, G. Cyclic mechanical stretching promotes migration but inhibits invasion of rat bone marrow stromal cells. Stem Cell Res. 2015, 14, 155–164. [Google Scholar] [CrossRef]
- Fu, X.; Halim, A.; Tian, B.; Luo, Q.; Song, G. MT1-MMP downregulation via the PI3K/Akt signaling pathway is required for the mechanical stretching-inhibited invasion of bone-marrow-derived mesenchymal stem cells. J. Cell. Physiol. 2019, 234, 14133–14144. [Google Scholar] [CrossRef] [PubMed]
- Naji, A.; Eitoku, M.; Favier, B.; Deschaseaux, F.; Rouas-Freiss, N.; Suganuma, N. Biological functions of mesenchymal stem cells and clinical implications. Cell. Mol. Life Sci. 2019, 76, 3323–3348. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-B.; Zhang, G.-Y.; Xie, Y.; Zan, T.; Gan, Y.-K.; Yao, C.A.; Chiang, C.-A.; Wang, J.; Liu, K.; Li, H.; et al. Autologous Stem Cell Transplantation Promotes Mechanical Stretch Induced Skin Regeneration: A Randomized Phase I/II Clinical Trial. EBioMedicine 2016, 13, 356–364. [Google Scholar] [CrossRef]
- Maksimova, N.V.; Michenko, A.V.; Krasilnikova, O.A.; Klabukov, I.D.; Gadaev, I.Y.; Krasheninnikov, M.E.; Belkov, P.A.; Lyundup, A.V. Mesenchymal stromal cell therapy alone does not lead to complete restoration of skin parameters in diabetic foot patients within a 3-year follow-up period. BioImpacts. 2022, 12, 51–55. [Google Scholar] [CrossRef]
- Cai, Y.; Li, J.; Jia, C.; He, Y.; Deng, C. Therapeutic applications of adipose cell-free derivatives: A review. Stem Cell Res. Ther. 2020, 11, 312. [Google Scholar] [CrossRef]
- An, Y.; Lin, S.; Tan, X.; Zhu, S.; Nie, F.; Zhen, Y.; Gu, L.; Zhang, C.; Wang, B.; Wei, W.; et al. Exosomes from adipose-derived stem cells and application to skin wound healing. Cell Prolif. 2021, 54, e12993. [Google Scholar] [CrossRef]
- Deng, M.; Wang, X.; Yu, Z.; Cai, Y.; Liu, W.; Zhou, G.; Wang, X.; Yu, Z.; Li, W.; Zhang, W.J. Cell-free fat extract promotes tissue regeneration in a tissue expansion model. Stem Cell Res. Ther. 2020, 11, 50. [Google Scholar] [CrossRef]
- Kimbrel, E.A.; Lanza, R. Next-generation stem cells—ushering in a new era of cell-based therapies. Nat. Rev. Drug Discov. 2020, 19, 463–479. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, Z.; Wang, J.; Zhang, X.; Lei, L.; Zhang, Y.; Su, Y.; Ma, X. Effects of Botulinum Toxin A on the Blood Flow in Expanded Rat Skin. J. Investig. Surg. 2022, 35, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Jinming, W.; Caiyue, L.; Baojin, W.; Antang, L.; Yingfan, Z.; Hui, W.; Lie, Z.; Hua, J. Effects of Platelet-Rich Plasma on Tissue Expansion in Rabbits. Aesthetic Plast. Surg. 2017, 41, 454–460. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Song, Y.; Xiong, S.; Wang, T.; Liu, W.; Yu, Z.; Ma, X. Mechanical Stretch Induced Skin Regeneration: Molecular and Cellular Mechanism in Skin Soft Tissue Expansion. Int. J. Mol. Sci. 2022, 23, 9622. https://doi.org/10.3390/ijms23179622
Guo Y, Song Y, Xiong S, Wang T, Liu W, Yu Z, Ma X. Mechanical Stretch Induced Skin Regeneration: Molecular and Cellular Mechanism in Skin Soft Tissue Expansion. International Journal of Molecular Sciences. 2022; 23(17):9622. https://doi.org/10.3390/ijms23179622
Chicago/Turabian StyleGuo, Yaotao, Yajuan Song, Shaoheng Xiong, Tong Wang, Wei Liu, Zhou Yu, and Xianjie Ma. 2022. "Mechanical Stretch Induced Skin Regeneration: Molecular and Cellular Mechanism in Skin Soft Tissue Expansion" International Journal of Molecular Sciences 23, no. 17: 9622. https://doi.org/10.3390/ijms23179622
APA StyleGuo, Y., Song, Y., Xiong, S., Wang, T., Liu, W., Yu, Z., & Ma, X. (2022). Mechanical Stretch Induced Skin Regeneration: Molecular and Cellular Mechanism in Skin Soft Tissue Expansion. International Journal of Molecular Sciences, 23(17), 9622. https://doi.org/10.3390/ijms23179622